㈠肾上腺皮质激素及类固醇激素的生物合成
肾上腺皮质可分泌多种激素,按生理生化功能及分泌组织,可分做三类:①球状带分泌的盐皮质激素(mineralocorticoide),主要是醛固酮(aldosterone)和脱氧皮质酮(deoxycorticosterone);②束状带分泌的糖皮质激素(glucocorticoide),主要有皮质醇 (cortisol)及少量的皮质酮(corticosterone);③网状带分泌的性激素,如脱氢异雄酮(dehydroepiandrosterone)、雄烯二酮(androstenodione)及少量雌激素。从化学结构上看,这三类激素及性腺合成的其他性激素,均是胆固醇的衍生物,故统称类固醇激素(steroid hormones),而上述三类肾上腺皮质激素又合称皮质类固醇(corticosteroids)。
类固醇激素在人体内均是以胆固醇为原料,经过一系列酶促反应而合成的,只是由于某些酶活性在某些内分泌腺或同一腺体不同的组织中特别高,从而生成不同的激素。类固醇激素的主要合成途径及产物见图12-3。图中标示出了酶活性存在腺体差异性的反应步骤,以及临床常见的先天性酶缺陷所影响的步骤,有助于有关酶缺陷的临床生化诊断参考。
有关盐皮质激素的临床生化在第五章已作介绍,性激素的临床生化将在本章第五节讨论,故本节将只介绍有关糖皮质激素的临床生化。
㈡糖皮质激素的运输及代谢
释放入血液中的糖皮质激素主要为皮质醇及10%左右的皮质酮。二者均约75%左右与肝脏合成的一种α1-球蛋白,即皮质素转运蛋白(transcortin),亦称皮质类固醇结合球蛋白(corticosteroid-bindingglobulin,CBG)可逆结合,15%与白蛋白可逆结合,仅10%左右以游离形式存在。CBG对糖皮质激素的亲和力高,但每分子CBG仅有一个结合部位,且血浆浓度低,故其结合容量有限。白蛋白虽然与糖皮质激素亲和力低,但可有多个结合位点,血浆浓度又高,因此结合容量大。当血中皮质激素浓度明显升高时,与CBG结合易达饱和,将出现与白蛋白结合部分及游离部分比率不成比例的升高。只有游离糖皮质激素才能进入靶细胞发挥生理生化作用及反馈调节自身分泌。
糖皮质激素的代谢主要在肝细胞中进行。主要反应方式为C-3酮基及环节中双键被加氢还原,生成多种加氢代谢物,以四氢皮质醇最多,此外尚有少量二氢、六氢代谢物。90%氢化皮质醇等代谢物及少量原型糖皮质激素,与葡糖醛酸或硫酸结合成相应的酯化物,从尿中排出,亦有少量可随胆汁排入肠道随着粪便排泄。以游离原型从尿中排出的皮质醇仅为血液总量的1%左右。
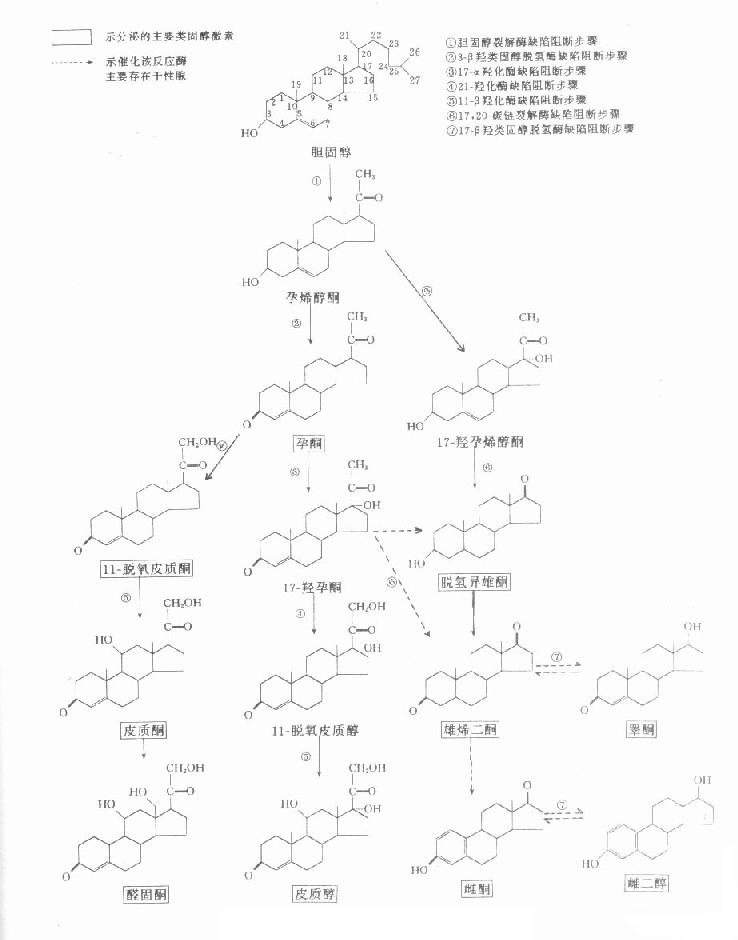
图12-3 类固醇激素生物合成的途径
㈢糖皮质激素的生理生化功能
游离皮质醇等糖皮质激素可经靶细胞扩散入胞质内,与其受体结合。糖皮质激素-受体复合物转运入细胞核内,可启动某些DNA片段转录,生成的mRNA进入胞质,指导合成特异的酶和脂皮素(lipocortin)等蛋白质或肽类介质,产生广泛的生理生化作用。生理性浓度下,糖皮质激素的主要功能为:
⒈调节糖、脂肪、蛋白质三大营养物质的代谢 对糖代谢,糖皮质激素可促进糖原异生,增加肝糖原和肌糖原含量,另一方面又抑制除脑和心脏外其他组织对糖的利用,使血糖升高。对蛋白质代谢,可促进除肝脏外多种器官、组织的蛋白质分解,抑制蛋白质的合成,升高血中氨基酸,出现尿酸、尿素氮排泄增多等负氮平衡表现。糖皮质激素能激活四肢皮下的脂酶,促进这些部位的脂肪分解,血脂肪酸升高,并使脂肪呈向心性重新分布。
⒉影响水电解质代谢 糖皮质激素有弱的盐皮质激素样潴钠排钾作用,亦有弱的促尿排钙排泄及抗利尿激素作用。
⒊允许作用(permissibleaction) 机体内其他一些激素、神经递质等生物活性物质的作用,需有适当浓度的糖皮质激素存在,才能正常表达,此即糖皮质激素的“允许作用”。主要为对肾上腺素及胰高血糖素的作用。
但高浓度的糖皮质激素如药用或肾上腺皮质功能亢进等,则除上述作用增强外,还可表现出抑制炎症、免疫反应,影响血细胞等作用,将在肾上腺皮质功能亢进症中介绍。
㈣糖皮质激素分泌的调节
和甲状腺激素分泌调节相似,肾上腺糖皮质激素的合成和分泌亦主要受图12-1所示的下丘脑-垂体-内分泌腺调节轴的控制。血液中游离糖皮质激素水平的变化,负反馈地引起下丘脑及垂体分别释放促肾上腺皮质激素释放激素(corticotropin releasing hormone,CRH)和促肾上腺皮质激素(corticotropin 或adrenocorticotropic hormone,ACTH)的增多或减少。CRH为下丘脑产生的一种含41个氨基酸残基的多肽,可选择性地促进腺垂体释放ACTH。ACTH是腺垂体促肾上腺皮质素细胞释放的39肽激素,可通过作用于肾上腺皮质束状带或网状带细胞膜上的ACTH受体,激活腺苷酸-cAMP-蛋白激酶系统,促进细胞增殖,合成和分泌糖皮质激素、性激素增多。持续的高ACTH状态仅早期一过性地引起盐皮质激素分泌增加,无持久影响。ACTH和CRH亦可负反馈地调节下丘脑CRH的释放。和甲状腺激素分泌调节不同,在肾上腺皮质激素的分泌调节中,最主要的是血液中游离糖皮质激素对下丘脑CRH释放的负反馈调节。ACTH和糖皮质激素的分泌存在明显的昼夜节律,分泌高峰见于晨6-8小时,低谷在午夜22-24时。此外,糖皮质激素是机体应激反应时释放的主要激素,因此,各种伤害性刺激均可通过高级神经中枢-下丘脑-垂体-肾上腺皮质轴,促进糖皮质激素的分泌。
除垂体外,一些垂体外的肿瘤主要是肺燕麦细胞癌,其次为胸腺癌、胰岛细胞癌、类癌、甲状腺髓样癌、嗜铬细胞瘤等,亦可分泌异源性ACTH。但这些异位肿瘤ACTH的分泌既不受血液糖皮质激素水平的负反馈调控,也不受CRH促进。此外,近年还发现有少数肿瘤可不受糖皮质激素反馈调节地释放异源性CRH。
现已明确,无论是腺垂体还是异位肿瘤分泌的ACTH,均和γ-黑色细胞刺激素(γ-melanocyte stimulating hormone,γ-MSH)、β-促脂解素(β-lipotropichormone,β-LPH)、β-内啡肽(β-endophin)等,来自同一由265个氨基酸残基组成的大分子前体蛋白,称阿片皮质素原(proopiomelanocortin,POMC)。ACTH等有关多肽激素与POMC的关系见图12-4。
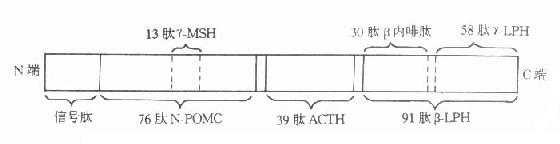
图12-4 阿片皮质素原及其多肽激素裂解产物的关系