一、体细胞突变
肿瘤可以看作是在个体遗传素质的基础上,尤其是在个体对肿瘤的遗传易感性基础上,致癌因子引起细胞遗传物质结构或功能异常的结果。这种异常大多数不是由生殖细胞遗传得来,而是在体细胞中新发生的基因突变所致。发生突变的癌前细胞在一些促癌因素的作用下发展为肿瘤。因此,有人认为,多数为肿瘤可看成是一种体细胞遗传病。
在自然界,基因突变是经常发生的,突变如果发生在与细胞增殖有关的基因,就可能导致细胞摆脱正常的生长控制,表现出恶性细胞的表型性状。
许多致癌物都是致突变物。它们大多数能引起DNA的损伤。这些损伤可以修复,也可以导致细胞死亡。如果DNA的修复不正常,细胞虽可继续存活,但却成了潜在的癌细胞,例如在着色性干皮病时,细胞由于缺乏DNA修复酶,因而在DNA被紫外线损伤后不能正常切除修复,结果导致皮肤癌发生。
肿瘤既然是体细胞突变的结果,则它就起源于单个突变细胞(单克隆)。许多肿瘤细胞群都具有相同的染色体畸变和同工酶,就是肿瘤发生的单克隆学说的证据。
两次突变说 一些细胞的恶性转化需要两次或两次以上的突变。第一次突变可能发生在生殖细胞或由父母遗传得来,为合子前突变,也可能发生在体细胞;第二次突变则均发生在体细胞本身,这就是两次突变(击中)说。两次突变说对一些遗传性肿瘤,如视网膜母细胞瘤的发生作出了合理解释。遗传型的视网膜母细胞瘤发病很早,并多双侧性或多发性。这是因为患儿出生时全身所有细胞已有一次基因突变,只需要在出生后某个视网膜母细胞再发后一次突变(第二次突变)。就会转变成为肿瘤细胞,故较易表现为双侧性或多发性。非遗传型视网膜母细胞瘤的发生则需要同一个细胞在出生后积累两次突变,而且两次都发生在同座位,因而概率很小,因此发病较晚,不具有遗传性,并多为单侧性,但该座位如果已发生过一次突变,则较易发生第二次突变,这也是非遗传型肿瘤不是太少的原因。
一些学者认为,体细胞癌变并不一定有基因结构的改变,当基因以外的物质或因素如蛋白质、RNA、生物膜发生了改变,而这些改变也能使与生长、分休有关的基因异常关闭或启动,此时,细胞也能转化为癌细胞,这一观点称为基因外调节学说。近年来对遗传印记的研究表明,基因的修饰如甲基化也能影响肿瘤有关基因的表达,从而促进细胞的转化。
二、癌基因
与肿瘤发生相关的基因可分为两大类:一类称为癌基因(oncogenes),它们能促进细胞的生长和增殖;另一类称肿瘤抑制基因(tumor-suppresor gene)或抑癌基因,它们能调节细胞的生长和分化而抑制肿瘤的发生。这两类基因的作用正好相反。它们的异常,或增强细胞生长和增殖,或去除正常的生长抑制,结果都会导致肿瘤发生。
癌基因:在致瘤病毒、人体和动物肿瘤都发现了能导致细胞恶性转化的核酸片段,称之为癌基因。来自病毒的称为病毒癌基因(v-onc),来自细胞的称为细胞癌基因(c-onc)或原癌基因(proto-oncogene),它们具有转化的潜能,可被激活成为癌基因。后来发现从酵母菌到人类的正常细胞几乎都有与之类似的片段。这种进化上的高度保守性表明它们具有重要的生物学意义。
最早发现的病毒癌基因是鸡Rous肉瘤病毒的v-src基因,以后又发现了与之同源的、见于正常鸡细胞的细胞癌基因(c-src)。许多癌基因都以最先见于的病毒来命名,如见于猴肉瘤病毒(Simian sarcoma)的称为sis基因,见于鸟类髓细胞增多症病毒(Myelocytomatosis)的称为myc基因等。最早发现的人类癌基因是H-ras基因,它是通过用人膀胱癌细胞系提取的DNA片段转染小鼠细胞,引起后者恶性转化而证实。以后在其他许多肿瘤和白血病也证明了人癌基因的存在。
1.癌基因的功能和分类已知的原癌基因已近100种,其中许多已定位不同的染色体区带。这些基因与细胞的生长、增殖等基本功能有关。它们或编码生长因子、生长因子受体和蛋白激酶而在生长信号的传递和细胞分裂中发挥作用;或者编码DNA结合蛋白而参与基因的表达或复制的调控(表9-2)。因此,可以按原癌基因的产物将其分为若干类型。如以src为代表的酪氨酸激酶类,以ras 为代表的G蛋白类,以myc为代表的核蛋白类,以sis为代表的生长因子类,以reb为代表的生长因子受体类等。原癌基因在个体发育或细胞分裂的一定阶段十分重要,但在成体或平时却不表达或表达受到严格的控制,当其发生突变或被异常激活时,产生的癌蛋白在性质或数量上异于正常,就可能导致细胞发生恶性转化。
表9-2 一些重要的癌基因
| 癌基因 | 来源 | 染色体定位 | 生化性质 |
| 信号传导蛋白 | |||
| abl | Abelson鼠白血病病毒 | 9q34 | 蛋白激酶 |
| src | 鸡Rous肉瘤病毒 | 20q12-13 | 蛋白激酶 |
| trk | 人大肠癌 | 1q32 | 蛋白激酶 |
| H-ras | Harvery大鼠肉瘤病毒 | 11p15.5 | GTPase |
| N-ras | 各种人类肿瘤(如神经母细胞瘤) | 1p13 | GTPase |
| 分泌生长因子 | |||
| sis | 猴肉瘤病毒 | 22q12.3-13.1 | 血小板生长因子(PLGF)β链 |
| 生长因子细胞 | |||
| 表面受体 | |||
| erb-A | 17q21-22 | 固醇受体 | |
| 核蛋白结合蛋白 | |||
| myc | 鸡肉瘤 | 8q24 | 结合DNA |
| N-myc | 人神经母细胞瘤 | 2p24 | 未知 |
2.癌基因的激活癌基因可以通过多种方式被激活而过度表达。
(1)突变激活:体细胞内的原癌基因可以因点突变而成为癌基因,产生异常的基因产物;也可由于点突变使基因摆脱正常的调控而过度表达。因此,突变激活又称为激活的质变模式(qualitative model)。例如在膀胱癌细胞株由于癌基因ras的12位密码子GGC变为GTC,使甘氨酸变为缬氨酸,结果导致细胞具有转化细胞的特征。现今已在膀胱癌、结肠癌等许多肿瘤发现了ras基因,后者编码一种膜蛋白,称为p21蛋白。
(2)易位激活:易位导致癌基因的重排或融合,产生异常的蛋白而使细胞转化。例如慢性粒细胞白血病9;22易位,形成了一种结构与功能异常的融合基因bcr-abl。它编码的蛋白能促成细胞的恶性转化。
原癌基因通过易位插到强力的启动子附近也可导致激活。Burkitt淋巴瘤的t(8;14)即c-myc癌基因由8号染色体易位到14号染色体的免疫球蛋白重链基因附近,易位使c-myc置于免疫球蛋白H链基因的启动子控制下。免疫球蛋白基因是一个活跃的基因,为了抵抗进入体内的各种抗原,它不断编码各种抗体蛋白,其启动子特别活跃,因而易位的c-myc基因转录活性明显增高。增多的myc蛋白质使一些控制生长的基因活化,最终导致细胞恶变。
(3)癌基因扩增:原癌基因还可因某种原因自身扩增而过度表达。在肿瘤细胞尤其是胚胎神经组织肿瘤细胞中有时见到的双微体(double minutes)和染色体上的均染区 (homogenouslystaining region)就是原癌基因DNA片段扩增的表现。例如,在肿瘤细胞中c-myc癌基因可扩增数百到数千倍。
三、肿瘤抑制基因
肿瘤抑制基因又称为抑癌基因或抗癌基因(anti-oncogenes)。它们的功能是抑制细胞的生长和促进细胞的分化。当两个等位都因突变或缺失而丧失功能,即处于纯合失活状态时,细胞就会因正常抑制的解除而恶性转化。视网膜母细胞瘤的Rb基因就是一个典型的例子。遗传型视网膜母细胞瘤患者出生时Rb基因的一个等位基因由于生殖细胞突变而丧失功能,出生后如视网膜母细胞中另一个等位基因发生了体细胞突变,这个细胞就会转化为肿瘤细胞。现今在不少的肿瘤均已证实了相应的抑制基因的存在、缺失及其在肿瘤发病中的作用(表9-3)。这些肿瘤在临床上都呈染色体显性遗传方式,但从基因水平上看,它们都是隐性的,因两个等位基因均有突变才能使细胞恶性转化。
视网膜母细胞瘤(Rb)基因定位于13号染色体1区4带,全长约200kb,有27个外显子,编码924个氨基酸的核磷蛋白(Rb蛋白),分子量为110kb,具有抑制肿瘤细胞增殖生长的作用。Rb基因的缺失或功能丧失不仅见于视网膜母细胞瘤,而且还见于骨肉瘤、小细胞肺癌、乳腺癌等肿瘤中。
表9-3 肿瘤抑制基因及其产物
| 肿瘤抑制基因 | 染色体定位 | 基因产物及其功能 | 基因异常引起的肿瘤 | |
| 遗传型 | 散发型 | |||
| Rb1 | 13q14 | p110细胞周期调节 | 视网膜母细胞瘤 | 小细胞肺癌等 |
| WT1 | 11p13 | 锌脂蛋白,结合DNA | Willm瘤 | 肺癌等 |
| p53 | 17p13.1 | p53,细胞周期调节 | Li-Frameni综合征 | 肺癌,乳腺癌等 |
| NF1 | 17p11.2 | GTP酶活化蛋白 | 神经纤维病,Ⅰ型 | - |
| DCC | 18q21 | 参与细胞表面作用 | 未知 | 结肠直癌 |
| APC | 5q21 | 家族性腺瘤息肉 | ||
| MCC | 5q21-22 | 家族性腺瘤息肉 | ||
如果抑癌基因的纯合型丢失是基因缺失的结果,可以通过比较正常组织与癌组织有关DNA片段的杂合性丢失(loss of heterozygosity,LOH)来证实。例如,正常组织的DNA等位片段与特定内切酶和特定的探针杂交后证明是杂合的,而癌组织的等位片段经证明是纯合的,表明抑癌基因发生了杂合丢失。
然而并非所有的抑癌基因都必需有两个等位基因的丧失才导致肿瘤发生。现知APC基因在这一过程中作用。同图可见,在肿瘤发生发展的过程中既有肿瘤抑制基因的丢失,也有癌基因的活化,而且不止一种。如结肠癌发生的过程中,就有多个基因的异常,其中包括ras癌基因和p53、APC和MCC等几个抑癌基因。这些改变中任何一种基因异常如果是遗传的,这就构成了肿瘤的家族性易感性的基础。
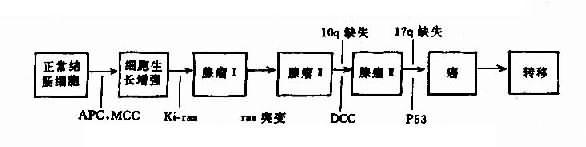
图9-8 结肠癌发生的各个阶段和肿瘤相关基因的异常
恶性肿瘤的转移是一个复杂的过程,包括瘤细胞由原发肿瘤脱落,进入细胞外基质和血管或淋巴管,并在远处适宜的组织中生长。近年来的研究发现,存在着促进转移的转移基因(metastatic genes)和抑制转移的基因(metastasissuppressor genes)。
1.转移基因一些编码细胞表面受体的基因可能和瘤细胞的转移有关。例如整合素(integrins)是一类细胞表面粘合受体,能识别细胞基质中的粘附蛋白,起着固定细胞抑制其迁移的作用。可以设想,这些受体基因的突变和失去功能将有利于瘤细胞的转移。
瘤细胞的浸润能力与其分泌的能降解基质的蛋白有关,已知内糖苷酶和Ⅳ型胶原酶能降解基底膜中的相应成分,增加瘤细胞侵袭基底膜的能力。
最后,用许多癌基因ras、fos、mos、src,或突变了的肿瘤抑制基因如p53基因的片段转染培养中的细胞,都可提高这些细胞的浸润和转移能力。
2.转移抑制基因一些基因编码的蛋白酶能够直接或间接地抑制具有促进转移的蛋白。例如金属蛋白组织抑制因子基因(TIMP)编码一种糖蛋白,能与转移密切有关的胶原酶结合,降低瘤细胞的侵袭和转移能力。
在人和小鼠中已发现nm23基因的表达与乳腺癌等肿瘤的转移密切相关。nm23基因编码一个17kd的蛋白质,基因在转移的乳腺癌中表达高,而在无转移的肿瘤中表达低,表明nm23是一个肿瘤转移抑制基因。至于它如何抑制肿瘤转移还有待阐明。
除了肿瘤细胞外,受体细胞的一些基因在转移灶生成中也具有一定意义。由此可见转移基因、转移抑制基因与宿主有关基因在表达最终决定于肿瘤细胞的转移。
综上所述,肿瘤的发生发展是一个复杂的生物学过程,它是细胞遗传物质异常的结果,同时也涉及机体的内环境中的各种因素,包括机体的免疫能力、各种生长因子和生物活性物质,而它们都是基因表达的结果,其中癌基因与抑癌基因的异常起着关键的作用。