(1)维生素A 维生素A亦名视黄醇(Retinol,VA),在自然界中的多是全反式棕榈酸酯。他的衍生物具有特殊的生理功能,如视黄醛(Retinyl Aldehyde,VA醛)对暗适应有效;甘露糖视黄醇磷酸(MannosylRetiny Phosphate,MRP)也具有某些生理功能。VA衍生物的分子式列于图5-1,活性列于表5-2。
维生素A及其衍生物易氧化。氧化剂MnO2可使之成为醛,还原剂双可将维生素A酸还原到要醛或醇。对碱较稳定,但对酸不稳定,使其脱氢或将双键重新排列。在光或碘的作用下,全反式可变为较稳定的11顺式异构体。在强光下形成二聚体或多聚体。
(2)胡萝卜素 植物中胡萝卜素在体内分解为维生素A,并具有维生素A的生理作用。
表5-2 维生素A及基其衍生物的活性
| 化学变化 | 产物 | 生物活性 |
| 酯化 | 自然及人工产品 | 100 |
| 氧化 | 醛 | 100 |
| 顺式异构 | 顺式异构体 | 23~75 |
| 醚 | 苯及甲基醚 | 10~100 |
| 脱氢 | 维生素A2(3-脱氢视黄醇) | 30 |
| 失去氧 | Axerophene | 10 |
| 酮 | C21-酮 | 3 |
| 去甲基 | 新维生素A | 3 |
| 增加-CH2- | 高维生素A | 1.5 |
| 脱水 | 脱水维生素A | 0.4 |
| 氧化 | 环氧物 | |
| 氢化 | 二氢维生素A等 |
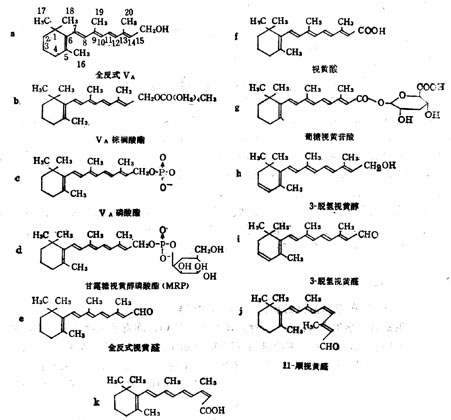
图5-1 维生素A衍生物的分子式
β-胡罗卜素含有两个β-紫罗兰酮环(β-Ionone ring)和四个异戊二烯侧链,加水断裂为两分子维生素A如图5-2。但实际上膳食中β-胡萝卜素只有1/6右变为维生素A。自然界中有环与维生素A相同,他们的维生素A活性低于β-胡萝卜素,有的没有与维生素A相似的环,没有生物活性(表5-3,5-4)。
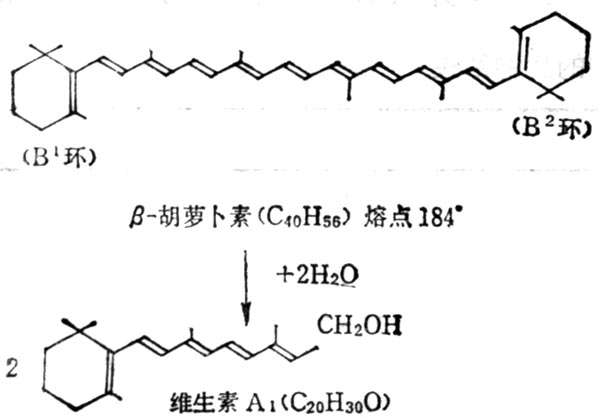
图5-2 β-胡萝止素水解为维生素A
5.2.2 维生素A及胡萝卜素的代谢
(1)胡萝卜素的吸收 胡萝卜素的微胶粒溶液在小肠内吸收。油溶液中者吸收最好,磷脂有助于形成微胶粒溶液而利于吸收。胆盐不但促进胡萝卜素运输至肠细胞,助其与细胞表面相结合,并促进胡萝卜素的分解。维生素E及其他抗氧化剂可保护侧链的共轭双键系统免于氧化。胡萝卜素进入到小肠细胞内,在胞浆内胡萝卜素双氧化酶(Carotene Dioxygenase)作用下,将1mol氧加入到中间位置的双键上,将其分解为视黄醛,但也可从一端将其分解生成具有与维生素A相同的环,侧链较长的醛即apo胡萝卜醛。所生成的醛双为脱氢酶作用还原为醇再酯化。小肠及肝都有胡萝卜素双氧化酶,但其活力以小肠中者较高,以器官计,肠为肝的2倍,以重量计为4~7倍。静脉注射β-胡萝卜素,在肝中也可转变为视黄醇,也以视醇酯形式储存,但能力有限。
胡萝卜素吸收是扩散性的,其吸收量与剂量大小有相反关系。类胡萝卜素都可吸收进入血浆内。血浆中的类胡萝卜素可分为β-胡萝卜素、叶黄素,番茄红素及其他类胡萝卜素,视摄入情况而定。可存在于肝、脂肪、肾、皮肤及血管粥样硬化的斑块中。
表5-3胡萝卜素化合称的环结构
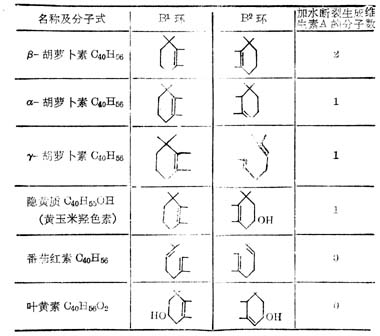
表5-4 类胡萝卜素化合物的生物活性(以β-胡萝卜生物活性为100%)
| 类胡萝卜素 | 活性 | 类胡萝卜素 | |
| β-胡萝卜素 | 100 | β-半胡萝卜素 | 有 |
| α-胡萝卜素 | 50~54 | β-apo-8'-胡萝卜醛 | 72 |
| γ-胡萝卜素 | 42~50 | β-apo-8'-胡萝卜酸 | 有 |
| β-新胡萝卜素 | 20~40 | 脱水叶黄素 | 21 |
| 3,4去氢-β-胡萝卜素 | 75 | 玉米黄素(隐黄质) | 57 |
| β-胡萝卜素 | 21 | 反-顺异构体比全反式的活性要小 | |
| β-胡萝卜素 | 50 |
(2)维生素A的吸收及储存维生素A为主动吸收,需要能量,速率比胡萝卜素要快7~30倍。食物中的维生素A为酯式,经肠中胰液或绒毛刷状缘中的视黄酯水解酶分解为游离式进入到小肠壁内,又为肠内细胞微粒体中的酯酶所酯化,合成维生素A棕榈酸酯,摄取维生素A3~5h后,吸收达到高峰。维生素A也需要胆盐。维生素E也可防止维生素A氧化破坏。维生素A与乳糜微粒相结合由淋巴系统输送到肝,酯式水解进入肝,然后又再酯化为棕榈酸酯。肝实质细胞负责摄取及新进入的维生素A代谢,一部分维生素A由实际细胞转入类脂储存细胞(Lipocyte),这种细胞中有许多类脂滴,如维生素A摄取量大,这些类脂滴大而多。85%的维生素A在类脂滴中,还有一些在高尔基体中。
维生素A的吸收与储存量均较大,大鼠每可吸收20,000IU。若维生素A摄入速度很快,VA酯与脂蛋白的结合已达饱合,多余者以酯式流入到血浆中。每日给大鼠50,000IU,4周后给以无维生素A膳食3周,血浆中维生素A酯较正常者多,而维生素A值正常。一方面由于组织的需要,维生素A从肝中释放经血流到靶组织,另一方面过剩的维生素A也从肝到血流中。
高蛋白膳食可以增加维生素A的利用,因而加速维生素A储存的空竭(Depletion)。蛋白质营养不良时,维生素A的吸收及胡萝卜素转变为维生素A的能力都受到影响,因而使维生素A的肝储存量降低。怀孕可使储存量增加,大鼠孕娠17~21天时,肝储存可增加2倍,为制造维生素A丰富的初乳及乳汁作准备。胎儿维生素A储存量甚少,不受母亲营养情况的影响。初生婴儿肝维生素A浓度仅为成人1/10。60岁以后储存量又低于成人。肝储存量可能与性别有关系,雌鼠比雄鼠储存较多。吃维生素A缺乏饲料,雄鼠肝储存空竭比雌鼠要快。男性的血清维生素A水平比女性稍高。夜盲的发生亦以男性较多。肾脏内也能储存维生素A,但其量仅为肝的1%。眼色素上皮组织内的维生素A是以酯式存在的,专为视网膜使用而储备,其空竭速率比肝中者要慢一些。
(3)维生素A的运输当靶组织需要维生素A时,维生素A从肝中释放出来,运输到靶组织。这个过程首先将肝内储存维生素A酯经酯酶水解为醇式,与视黄醇结合蛋白(Retionl BindingProetin,RBP)结合,再与前白蛋白(Prealbumin,PA)结合,形成维生素A-RBP-PA复合体后,才离开肝脏。经血流入靶组织。维生素A在一般情况下必须与蛋白质结合,使之具有水溶性,并较稳定。此外,还可减少维生素A对细胞的毒性,因细胞膜对复合体有识别能力,但对未结合的维生素A无识别力,以致对维生素A的摄取失去控制,过多维生素A进入细胞而产生毒性。
RBP为肝实质细胞合成的多肽,分子量21,000,电泳在α部分。肝每天能合成190mg,一分子RBP与一分子维生素A结合。维生素A-RBP-PA进入血流中,由于分子较大,不能从肾脏滤出。血流中RBP的水平受维生素A的影响,如果无维生素A可资运输时,RBP则积累于肝内。正常血浆RBP水平为40~50μg·m-1,维生素A缺乏的夜盲症状出现时,RBP则降为20μg·m-1。蛋白质缺乏,RBP合成减少,维生素A-RBP不能形成,维生素A结合的RBP-PA,其量为正常值的20%,但已变性,不能再与维生素A结合。PA的血浆浓度受维生素A营养状况的影响较少,其正常值为200~300μg·m-1,他不但与RBP结合,而且也与甲状腺素结合,两者互不影响。
维生素A-RBP-PA复合体随血流到肠粘膜、膀胱、角膜及上皮组织等靶细胞后,细胞膜上有RBP的特殊受全,可与RBP结合,并将维生素A释放出来,进入细胞内。RBP与维生素A分开后已变性,丧失与维生素A,PA或细胞膜上受体的结合能力。此种游离的RBP在肾小球中可滤过,而在肾小管重吸收,为肾皮层细胞所摄取,并在其溶酶体中分解为氨基酸。血浆中还有一种未与PA结合的维生素A-RBP,其量约为1μg·m-1。这部分可从肾小球滤过,在肾小管中重吸收,并浆维生素A释放出来。因此肾中有少量维生素A积累,然后酯化输送至肝。
维生素A进入到靶细胞后,立即与细胞视醇结合蛋白(Cellular retinol Binding Protein,CRBP)相结合。维生素A酸在运输过程中不需要与RBP相结合,但进入细胞内必须视黄酸结合蛋白相结合。
肝病患者的血浆蛋白合成受到影响,RBP及PA水平偏低而影响维生素A的血浆水平。肾病患者RBP分解代谢减弱,使RBP值增高。但囊纤维变性患者,肝内维生素A储存量适当,但血浆中RBP、PA及维生素A的水平均低。RBP也受内泌的调节,雄性动物血浆的RBP水平比雌性高。月经期或口服避孕药的妇女的RBP都有变化。新生儿的RBP只有成人的一半。这种水平一直维持至青春期,以后将增高。
(4)维生素A分解代谢及排出维生素A分解代谢途径可能如图5-3。视黄醇通过氧化转变为维生素A酸,其中一部分异构为β顺式。全反式或β顺式者均为维生素A的代谢产物,有维持上皮组织分化的活性,但体内不能储存,很快消失。注射维生素A酸4h后,肝中只剩下10%,24h后肝中已无。以后5,6位环氧化形成5,6环氧维生素A酸(5,6-epoxyretinoic acid)或环上4位羟基化,再进一步氧化形成4-氧-络维生素A酸(4-oxo-Retinoicaxcid)或形成葡萄视黄苷酸(retinoylglucronide)。大鼠注射标记维生素A酸后,在胆汁中60%为维生素A酸及基葡萄糖苷酸,少于40%的为13顺式葡糖视黄苷酸,还有少量5,6位从环氧维生素A酸及基葡萄糖视黄苷酸为0.2~0.4μg,这些维生素A代谢从胆汁流入小肠,形成肝肠循环。正常大鼠注射标记维生素A酸2h后,在小肠中的放射性约占剂量的7.2%,而胆道插管者仅为2.4%。在肾脏中也有维生素A代谢物。大鼠经胃给以标记的视黄醇或其酯后,在肾脏中除摄入物外,尚有全反式维生素A酸及5,6位环氧维生素A酸,但在血浆中并未发现后者,这种氧化作用可能仅在靶组织(如肾)中发生。他的活性仅为维生素A酸之1%,生物半寿期很短。
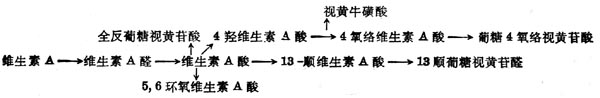
图5-3 维生素A的氧化代谢的可能途径
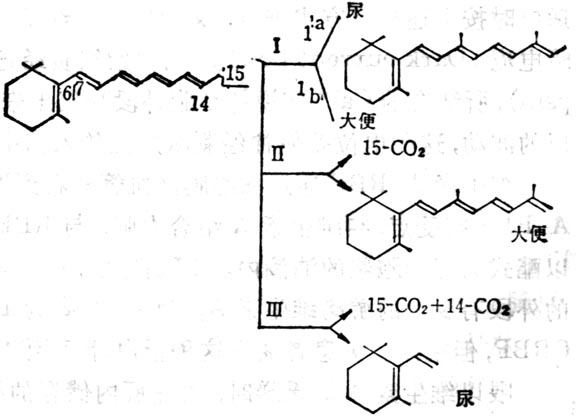
图5-4 维生素A的氧化代谢的可能途径
大鼠摄入维生素A后,大便及尿中都有其代谢物的排出。2/3排出物的异戊二烯侧链部分无改变。也有15-C氧化为CO2,有的侧链氧化分解成CO2及短链产品。大鼠给以20μg维生素A后,大便中排出以葡糖视苷酸为主,为主要排出途径,大便排出量变化较大,第2,3天达到高峰,约为剂量的9%,然后下降到较小数量。尿中排出24h内为剂量的8.7%,其中一半为水溶性者,一半为脂溶性者,尿排出量逐渐下降至第八天达到稳定,约为0.55%。尿中代谢产物β-紫罗兰酮环部分可氧化,甲基可脱去侧链中的双键可饱合,链也可缩枯木短。水溶性代谢物目前了解很少。
肝内储存的及摄入的维生素A都能被补充到需要维生素A的靶组织中去。因此,肝内维生素A的靶组织中。因此,肝内维生素A的储存量能影响维生素A的代谢率(即维生素A转变为尿及大便中维生素A代谢物之量)。SauberLich测定人的代谢率每天570~1250μg,这种差异由于肝储存量不同所致。注射标记维生素A,在试验中,发现维生素A有三种不同速率从尿中排出,亦即有三种不同代谢库:代谢库1代表新吸收的维生素A;代谢库2代表功能性和非功能性的维生素A;代谢库3代表功能性的代谢。在肝内储存空竭后的代谢率,才代表靶组织的功能性代谢率。新吸收的维生素A与体内储存者不同,二者在24h内不混合,血浆中新吸收者占90%,内源者仅占10%,二者逐渐混合,大鼠至第八天才完全混合。如果肝中维生素A储存量少,新吸收者在血浆中比例大,持续时间较长。如果肝储存量为0.6μg·m-1时,新吸收者由于其他维生素A已经空竭,需要维生素A补充,不能储存于肝内,而进入其他组织如肾等。若维生素A储存量较多时,肝储存量为18μg·m-1,60%存于肝内。从代谢库2来看,非功能性的代谢,即代表肝储存维生素A的分解。所以维生素A空竭的速率随维生素A储存量的指数而变化。大鼠的维生素A肝储存量可为全部生命需要量的数倍,但也可空竭,人体也是如此,储存量高代谢率亦高,摄入量高,代谢率也高。
5.2.3 生理功能
维生素A具有维持正常生长、生殖、视觉及抗感染的功能,但其作用机理迄今尚未完全清楚。
(1)维生素A与视觉的关系视网膜上有两种视细胞,即视杆细胞与视锥细胞,人类前者数量多,与暗视党有关;后者数量少,与明视觉及色觉有关,他们都有视色素,由视蛋白与生色团组成。各种动物的细胞的视蛋白不同,生色团由不同类型的维生素A醛组成。视杆细胞外段含有视色素是感光部分。视杆细胞膜连续不断地内陷,折起形成片层膜结成,每一层膜又由两层脂类分子膜由两层脂类分子膜构成称为双分子膜。视色素镶嵌在这种脂类双分子膜中。视色素视紫红质,由维生素A醛与视蛋白结合而成。维生素A醛由维生素A氧化而来,经异构酶作用使其变为11-顺式维生素A醛。维生素A醛由维生素A氧化而来,经异构酶作用使其变为11-顺式维生素A醛。维生素A醛由视蛋白之间的一级结构是11-顺维生素A醛的醛基与视蛋白中赖氨酸ξ氨基形成希夫碱(Schiff base)键,随之引起视蛋白高级结构改变,产生相互保护的二级结构,在黑暗中非常稳定。当光照时,一个视紫红质接受一个光子后,维生素A醛在11-C上扭成转成全反维生素A醛,视蛋白的立体构形也发生变化,维生素A醛又经视细胞外端的维生素A还原酶使成其变为维生素A,然后由色素上皮细胞微粒体中酯酶将其酯化而储存于色素上细胞内,需要时再异构为11-顺维生素A。暗适应按上述相反方式进行,又形成视紫红质。暗时Na+从视杆细胞内段移到外段,形成暗电流(Dark current flow)。当视紫红质经光异构后变为前光视紫红质(Prelumirhodopsin),所结合的Ca2+在视杆细胞外段释放出来,中止了Na+流动及暗电流,其结果产生电压的波动,这种电位差使神经刺激加强传入脑中发生光感,也可用视网膜电图测定。
维生素A-RBP通过脉络膜微血管分布到视网膜上皮组织,其细胞表面有维生素A-RBP接受点,与维生素A结合力强,与RBP结合弱,维生素A进入到色素上皮细胞内以酯式储存于胞浆的油滴内。暗适应的眼中,有一半的维生素A为11-顺式,在视杆细胞的外段有少量的游离维生素A,其中60%为11-顺式,生理上很重要,视网膜细胞中有CRBP,但视网膜炎患者缺少这种蛋白,因而维生素A不能进入到细胞内。
喂以维生素A缺乏膳食时,首先肝内储存的维生素A释放供给其他组织,肝的储存量下降,血浆维生素A正常。当肝储存量将近空竭时,血浆维生素A水平才下降,但由于视网膜的色素上皮组织有维生素A积累,视紫红质的维生素A醛下降较慢。色素上皮细胞内维生素A缺乏时,才有夜盲症出现。若视杆细胞内视紫红质减少,影响光的吸收,电流也减少。如视紫红质为正常值之74%,光阈上升14倍,若为正常值之16%,光阈上升680倍,维生素A非常缺乏时,光蛋白也减少,视细胞可以溶解。
(2)对糖蛋白质合成的影响 缺乏维生素A的动物的某组织,如小肠、角膜、气管上皮组织及血清等的特殊糖蛋白减少,给以维生素A或维生素A酸可以促进其合成。细胞膜表面的蛋白主要为糖蛋白,细胞膜的功能如接触抑制、分化及识别环境的能力,可能与糖蛋白有关。膜上有糖基视黄醇磷酸,所以,维生素A可能与膜的糖蛋白合成有关。免疫球蛋白也是糖蛋白,维生素A营养状况影响免疫功能,可能与此有关。
糖蛋白的合成步骤,首先为合成多肽链,然后纳入单糖,单糖与多肽链上的丝或苏氨酸的羟基或门冬酰胺相偶联形成糖蛋白。糖蛋白中的糖为甘露糖、岩藻糖、半乳糖、葡萄糖及乙酰葡萄胺。糖蛋白的合成需要脂类-糖作为中间体,其中脂类多为异戊二烯醇类的多萜醇或视黄醇。将单糖从GDP-单糖或UDP-章糖转移到维生素A或多萜醇上。如章糖为甘露糖结合到维生素A上,则生成甘露糖视黄醇磷酸酯(Mannosylretinylphosphate,MRP),若结合到多萜醇上生成甘露糖多萜醇磷酸酯(Dilichylphosphomannose,DMP),再将单糖转移至糖蛋白上。
维生素A在糖蛋白合成中变化分为三个步骤:①形成维生素A磷酸酯(Retinylphosphate,RP);②由RP合成MRP,这一步骤在微粒体内合成;③将单糖转移至糖蛋白上(图5-5)。
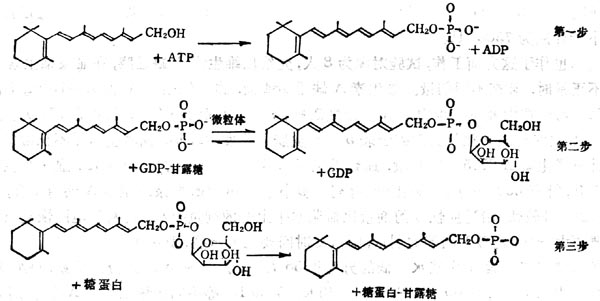
图5-5 VA在糖蛋白合成中的作用
MRP与DMP都是蛋白糖基化的中间体,由MRP合成的糖蛋白的分子量比从DMP合成要小一些。维生素A缺乏的仓鼠肝微粒体合成的MRP少而合成的DMP多。有些特殊蛋白,如大鼠血清糖蛋白(α1-巨球蛋白)的合成需要VA,他在肝内合成,含有2~7%甘露糖。维生素A严重缺乏时,降低至正常值的20%。维生素A的靶组织(肝、胆管、肠)的上皮组织中,都有MRP或半乳糖视黄醇磷酸酯。维生素A可使上皮细胞正常,呈柱状分泌粘液。维生素A缺乏时,上皮组织鳞状变形而角化。维生素A在糖蛋白合成中的作用与上皮组织的生理功能的关系,尚须进一步研究。
5.2.4 来源
动物的肝、肾、蛋及奶中的都有维生素A,尤以肝中最丰富,绿色蔬菜及红黄色蔬菜与水果中有胡萝卜素,但各种胡萝卜素的生物效用不一样,以β-胡萝卜素最高。供应量为1500~4000IU时,食物中1μgβ-胡萝卜素相当于0.167μg的维生素A,其他胡萝卜素仅为其一半。植物中有些黄红色素并无维生素A的效用(表5-4)。
胡萝卜素为脂溶性的,在动植物中与脂类相结合,在消化道中可以酶解,烹调中胡萝卜素比较稳定。我国的炒菜方法,胡萝卜素的保存率为76~94%,如烹调时间短,用水量少,温度低都可减少损失。植物老化过程中,胡萝卜素转变为环氧化合物,为氧化破坏的第一步。加工烹调及储存过程也容易异构,减少生物活性,食物加工中使用亚硫酸,产生自由基,对胡萝卜素也有破坏作用。食品制作方法也影响胡萝卜素的利用。胡萝卜片的胡萝卜素76%由大便排出,在匀浆中者只有44%排出。
5.2.5 需要量的研究
VA过去以国际单位IU表示,现在以视黄醇当量RE表示,
1视黄醇当量=1μg维生素A
二次大战时,英国医学研究会给16人以维生素A缺乏膳食作为试验组,为期61/2~25个月。7人为对照组(其中2人每日补充25IU维生素A,5人每日补充5000IU胡萝卜素)。当被试者有缺乏症状后,补充不同剂量的维生素A或胡萝卜素以测定需要量,在此研究中观察指标较多,但仅血浆维生素A及暗适应较为灵敏。结果最低需要量为1300IU维生素A,1500IU胡萝卜素(从大便中排出者不计算在内),正常需要量为2500IU维生素A,胡萝卜素为3000IU,但各种食物的胡萝卜素有效剂量不一样,所以正常需要量以食物中胡萝卜素计应为7500IU。
美军也作了这方面工作,试验对象为8人,先给以维生素A缺乏膳,在血浆维生素A及视力不正常时,补充不同剂量。维生素A缺乏184天,血浆维生素A从57~78μg%降至20~32μg%。视力及血浆维生素A显出缺乏症的最短时间为361天,最长为771天。血浆维生素A浓度与缺乏症的关系列于表5-5。皮肤角化发生较早,视网膜电图不正常发现较晚。补充维生素A37.5~150,血浆维生素A上升不明显,补充300μg,血浆维生素A略有上升,补充600μg者,明显上升,补充胡萝卜素1200μg,血浆维生素A略有上升,补充2400μg上升较快,暗适应恢复的剂量比浆水平上升的剂量要小,视网膜电图恢复正常的剂量要更大一些,若摄入同等的剂量。则恢复时间要延长一些(表5-6)。
我国曾将维生素A血浆水平低的男青年56人(血浆30~65IU),分成7组,每组8人,分别给以400,2000,3100,3500,3800,4500,5500IU等不同剂量维生素A。试验期为11周,观察血浆维生素A、暗适应及生理盲点的变化,每日摄入2000~3000IU者,血浆维生素A水平略有上升,但未达到100~120IU%,3500~5000IU者,在9周时使血浆维生素A达到100IU%,11周时维持在107~130IU%,这次试验未观察到暗适应的变化,但在试验初期,几乎所有试验对象都有生理盲点扩大的现象,摄取量为400IU者,生理盲点继续扩大,一次1,000,000IU大剂量,24h内恢复正常。摄取量2000~3000IU两组,至11周时恢复正常,摄取量3500IU以上的各组,至5~9周时大都恢复正常。因此,维生素A供应量为3500IU时,不仅能使血浆维生素A达到正常,而且生理盲点也正常。上述三次人体试验的地区、人种、工作不同,但其维生素A正常需要量均为1200~1300μg。
表5-5 美军维生素A缺乏症状与相应的血浆浓度
| 维生素A血浆浓度μg% | |||
| 被试者 | 暗适应不正常 | 视网膜电图不正常 | 皮肤角化 |
| 1 | 22~24 | 9~11 | 29~35 |
| 2 | 25~30 | 未发现 | 32~37 |
| 3 | 20~25 | 未发现 | 29~38 |
| 4 | 4~5 | 7~10 | 20~25 |
| 5 | 4~8 | 4~8 | 5~10 |
| 65 | 30~35 | 未发现 | 25~30 |
| 7 | 2~5 | 5~9 | 13~ 20 |
| 8 | 3 | 5~10 | 20 ~ 24 |
表5-6 美军维生素A需要量
| 维生素A需要量 | ||
| 维生素A(μg/日) | β-胡萝卜素(μg/日) | |
| 暗适应正常 | 150~300 | 300~600 |
| 视网膜电图正常 | 300~600 | 600~1200 |
| 皮肤正常 | 600 | 1200 |
| 血浆维生素A水平大于20μg% | 600 | 1200 |
| 大于30μg% | 1200 | 2400 |
①血浆30μg%以上肝储存量为中等水平。维生素A代谢率为570~1250μg。
5.2.6 临床应用
维生素A除用以防治缺乏病外,尚用以治疗皮肤角化,并对某些癌有预防作用。
(1)维生素A与皮肤病维生素A可抑制皮肤角化,维生素A酸也有这种功能,但它不储存于肝内,运输不需要RBP,可进入组织中,迅速代谢,很快从体内消失,毒性较少。所以用维生素A酸及其衍生物治疗毛囊角化、痤疮等。β-顺式维生素A及芳香族维生素A酸衍生物(如Etretinate)疗效分别为全反式维生素A酸的2.5及10倍。β-顺式维生素A酸用以抑制皮脂形成治疗痤疮有效。用Etretinate治疗牛皮癣及其他衍生物可迅速达到上皮真皮。
(2)维生素A癌维生素A可维持正常上皮组织的分化,维生素A缺乏的某些组织的形态变化与某些上皮组织癌症的早期病变(癌前期)相近,都是上皮组织鳞状变形。因此,维生素A可能在这一阶段中,对癌细胞起回转修复作用。维生素A缺乏,可能增加某些靶组织对致癌物质的敏感性。维生素A只在癌前期起作用,到第三期癌出现后已无效。大鼠的试验证实上述说法。
摄取大量的维生素A,储存于肝,血液中含量受到RBP的 限制,到靶组织之量有限。预防癌变也多用维生素A酸。现在正研制无毒或毒性较小而又能作用于靶组织的维生素A酸衍生物:①β-紫罗兰酮环的改变,β变为α,或变为二甲基乙酰戊烯环。如3甲基甲氧基环已烯环酸即Etretinate,可以有效地减低吸烟者的气管组织变形,膀胱癌复发病也减低;②侧链上的改变,9,11,13共轭双键为维生素A活性必要部分,必须保留。顺式异构体有活性,β-顺式维生素A酸最先临床应用于防治膀胱癌呼吸道癌,但他的毒性比维生素A酸酯或维生素A乙基酰胺要大;③末端极性基团的改变,维生素A甲酯或丁酯毒性低,活性高,肝中积累少,而在乳腺中较多,可用以预防乳腺癌的诱导,但尚未用于人体。
维生素A对免疫系统也有影响,他不但对疾病抵抗力有关。而且对一病毒引起的癌也有预防作用。小鼠黑瘤癌细胞移植给小鼠,未补充维生素A者,75%在3~5周内发展成瘤,而补充维生素A(每日5000IU,共补充5日者仅5%有瘤。关于这方面工作也在进行。
维生素A对癌预防作用可能由于它与核作用使基因改变。维生素A靶细胞中有维生素A或维生素A酸结合蛋白(CRBP,CRABP),维生素A或维生素A酸与之结合后,可能被携带运输至细胞核从而改变基因,影响细胞分化。CRBP在肾、肝、小肠、肺、脾、眼及睾丸中较多,而血清、脑、肌肉、脂肪、心脏中较中少。CRABP在胚胎及未分化的组织中多,有些癌组织中,CRABP的量比正常组织中者要多。组织中CRBP及CRABP之量决定维生素A或维生素A酸及其衍生物的生物作用。不同癌组织对维生素A及其衍生敏感性可能取决于细胞中CRABP之不同。
维生素A及胡萝卜素也试用于辅助治疗。癌在切除、放疗及化疗后,可使癌细胞水平降至低水平。维生素A或胡萝卜素对残余的癌细胞的分裂起到抑制作用,从而减少复发率或延长潜伏期。
(3)胡萝卜素与癌 流行病学调查中发现肺癌的发病率与胡萝卜素摄取量有相反关系,对于胃癌及前列腺癌也有相似的结果。美国麻省调查66岁以上的老人在5年内的死亡率,发现胡萝卜素摄取量多者,癌的危险性少些。
口腔粘膜细胞在烟、酒、X照射及咀嚼槟榔等刺激下,易有微核细胞(micronuclei cell),这种细胞是DNA损伤染色体破碎的标记,可能发展为癌前期的病变,如果给以维生素A及胡萝卜与槟榔咀嚼者,可以减低这种细胞的百分率,补充的时间愈长,减少的愈多。
在小鼠试验中,大剂量β-胡萝卜素可抑制2甲基苯蒽诱导的皮肤致癌作用,有些类胡萝卜素无此作用。
一般认为,胡萝卜素在小肠及肝中转变为维生素A,肾中仅有少量的可转变。但最近小鼠的试验给以β-胡萝卜素后,肺及皮肤中可积累维生素A,所以有可能β-胡萝卜至少转变为维生素A后才起作用。有些人认为胡萝卜素能熄灭单线态氧(Singlet O2)或捕捉自由基,这种单线态氧的反应性强,对机体有副作用,动植物代谢中产生,植物中类胡萝卜素有熄灭光合作用下产生的这类有害物质。胡萝卜素的抗癌作用可能由于这些作用。
胡萝卜素大量摄取后,除在皮肤脂肪积累使基呈黄色外,尚未发现有其他毒性。