Watson和Crick提出的DNA双螺旋结构属于B型双螺旋,它是以在生理盐溶液中抽出的DNA纤维在92%相对湿度下进行X-射线衍射图谱为依据进行推测的,这是DNA分子在水性环境和生理条件下最稳定的结构。然而以后的研究表明DNA的结构是动态的。在以钾或绝作反离子,相对湿度为75%时,DNA分子的X-射线衍射图给出的是A构象,A-DNA每螺旋含11个碱基对,而且变成A-DNA后,大沟变窄、变深,小沟变宽、变浅。由于大沟、小沟是DNA行使功能时蛋白质的识别位点,所以由B-DNA变为A-DNA后,蛋白质对DNA分子的识别也发生了相应变化。
一般说来,A-T丰富的DNA片段常呈B-DNA。采用乙醇沉淀法纯化DNA时,整个过程中,大部分DNA由B-DNA经过C-DNA,最终变构为A-DNA。若DNA双链中一条链被相应的RNA链所替换,会变构成A-DNA。当DNA处于转录状态时,DNA模板链与由它转录所得的RNA链间形成的双链就是A-DNA。由此可见A-DNA构象对基因表达有重要意义。此外,B-DNA双链都被RNA链所取代而得到由两条RNA链组成的双螺旋结构也是A-DNA。除A-DNA、B-DNA螺旋外,还存在B′-DNA、C-DNA、D-DNA等,其结构参数见表15-4。
表15-4 不同右手双螺旋DNA的结构参数
| 双螺旋 | 碱基倾 | 碱基夹 | 碱基间距 | 螺距 | 每轮碱 | 小沟宽/nm× | 大沟宽nm× |
| 角/(°) | 角(°) | /nm | /nm | 基数 | 小沟宽nm | 大沟宽nm | |
| B-DNA | 36.0 | 0.337 | 3.4 | 10 | 0.57×0.75 | 1.17×0.85 | |
| C-DNA | 6 | 38.0 | 0.331 | 3.1 | 9.3 | 0.48×0.79 | 1.05×0.75 |
| D-DNA | 45.0 | 0.303 | 0.13×0.67 | 0.89×0.58 | |||
| A-DAN | 20 | 32.7 | 0.256 | 2.8 | 11 | 1.10×0.28 | 0.27×1.35 |
总之,DNA的双螺旋结构永远处于动态平衡中,DNA分子构象的变化与糖基和碱基之间空间相对位置有关。
1979年,Wang和Rich等人在研究人工合成的CGCGCG单晶的X-射线衍射图谱时出人意料地发现这种六聚体的构象与上面讲到的完全不同。它是左手双螺旋,与右手螺旋的不同是螺距延长(4.5nm左右),直径变窄(1.8nm),每个螺旋含12个碱基对,分子长链中磷原子不是平滑延伸而是锯齿形排列,有如“之”字形一样,因此叫它Z构象(英文字Zigzag的第一个字母)。还有,这一构象中的重复单位是二核苷酸而不是单核苷酸;而且ZDNA只有一个螺旋沟,它相当于B构象中的小沟,它狭而深,大沟则不复存在(图15-7)。进一步的分析还证明,Z-DNA的形成是DNA单链上出现嘌呤与嘧啶交替排列所成的。比如CGCGCGCG或者CACACACA。
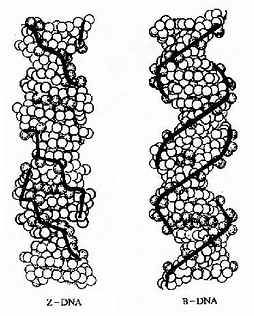
图15-7 Z-DNA和B-DNA
Z-DNA有什么生物学意义呢?应当指出Z-DNA的形成通常在热力学上是不利的。因为Z-DNA中带负电荷的磷酸根距离太近了,这会产生静电排斥。但是,DNA链的局部不稳定区的存在就成为潜在的解链位点。DNA解螺旋却是DNA复制和转录等过程中必要的环节,因此认为这一结构与基因调节有关。比如SV40增强子区中就有此结构,又如鼠类微小病毒DNS复制区起始点附近有GC交替排列序列。此外,DNA螺旋上沟的特征在其信息表达过程中起关键作用。调控蛋白都是通过其分子上特定的氨基酸侧链与DNA双螺旋沟中的碱基对一侧的氢原子供体或受体相互作用,形成氢键从而识别DNA上的遗传信息的。大沟所带的遗传信息比小沟多。沟的宽窄和深浅也直接影响到调控蛋白质对DNA信息的识别。ZDNA中大沟消失,小沟狭而深,使调控蛋白识别方式也发生变化。这些都暗示ZDNA的存在不仅仅是由于DNA中出现嘌呤一啶嘧交替排列之结果,也一定是在漫漫的进化长河中对DNA序列与结构不断调整与筛选的结果,有其内在而深刻的含意,只是人们还未充分认识而已。
DNA构象的可变性,或者说DNA二级结构的多态性的发现拓宽了人们的视野。原来,生物体中最为稳定的遗传物质也可以采用不同的姿态来实现其丰富多采的生物学功能。
多年来,DNA结构的研究手段主要是X射线衍射技术,其结果是通过间接观测多个DNA分子有关结构参数的平均值而获得的。同时,这项技术的样品分析条件使被测DNA分子与天然状态相差甚远。因此,在反映DNA结构真实性方面这种方法存在着缺陷。1989年,应用扫描隧道显微镜(scanning tummelingmicroscopy,STM)研究DNA结构克服了上述技术的缺陷。这种先进的显微技术,不仅可将被测物放大500万倍,且能直接观测接近天然条件下单个DNA分子的结构细节。STM技术的应用是DNA结构研究中的重要进展,可望在探索DNA结构的某些未知点上展示巨大潜力。