转录是在DNA模板某一位置上停止的,人们比较了若干原核生物RNA转录终止位点附近的DNA序列,发现DNA模板上的转录终止信号有两种情况,一类是不依赖于蛋白质因子而实现的终止作用,另一类是依赖蛋白质辅因子才能实现终止作用,这种蛋白质辅因子称为释放因子,通常又称ρ因子。两类终止信号有共同的序列特征,在转录终止之前有一段回文结构,回文序列是一段方向相反,碱基互补的序列,在这段互补序列之间由几个碱基隔开,不依赖ρ因子的终止序列中富含G·C碱基对,其下游6-8个A;而依赖ρ因子的终止序列中G·C碱基对含量较少,其少游也没有因固定的特征,其转录生成的RNA可形成二级结构即柄一噜噗结构,又称发夹结构,这样的二级结构可能与RNA聚合酶某种特定的空间结构相嵌合,阻碍了RNA聚合酶进一步发挥作用(图17-8)。除DNA模板本身的终止信号外,在入噬菌体中,发现一些蛋白质有协助RNA聚合酶跨越终止部位的作用,叫做抗转录终止蛋白,例如入噬菌体的N基因产物。
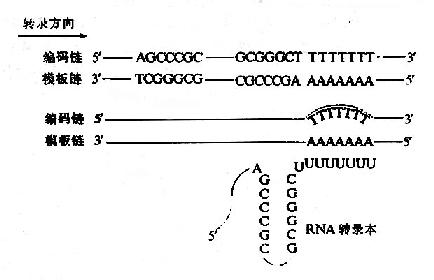
图17-8 原核生物转录作用的终止信号
真核生物由于RNA转录后很快就进行了加工,因此很难确定原初转录物的3’末端。病毒SV40的终止位点经过研究发现,很像大肠杆菌的不依赖ρ因子的终止子,转录后的RNA可形成一个发夹结构,3’末端带有一连串的U。爪蟾5sRNA的3’末端有4个U,它们前后的序列为富含G·C的序列,这是所有真核生物RNA聚合酶Ⅲ转录的终止信号。这种序列特征高度保守,从酵母到人都很相似,任何改变这种序列特征的突变都将导致转录终止位置的改变。