以反式作用影响转录的因子可统称为转录因子(transcription factors, TF)。RNA聚合酶是一种反式作用于转录的蛋白因子。在真核细胞中RNA聚合酶通常不能单独发挥转录作用,而需要与其他转录因子共同协作。与RNA聚合酶Ⅰ、Ⅱ、Ⅲ相应的转录因子分别称为TFⅠ、TFⅡ、TFⅢ,对TFⅡ研究最多。表19-2列出真核基因转录需要基本的TFⅡ。
表19-2 RNA聚合酶Ⅱ的基本转录因子
| 转录因子 | 分子量(kD) | 功能 |
| TBP | 30 | 与TATA盒结合 |
| TFⅡ-B | 33 | 介导RNA聚合酶Ⅱ的结合 |
| TFⅡ-F | 30,74 | 解旋酶 |
| TFⅡ-E | 34,37 | ATP酶 |
| TFⅡ-H | 62,89 | 解旋酶 |
| TFⅡ-A | 12,19,35 | 稳定TFⅡ-D的结合 |
| TFⅡ-I | 120 | 促进TFⅡ-D的结合 |
以前认为与TATA盒结合的蛋白因子是TFⅡ-D,后来发现TFⅡ-D实际包括两类成分:与TATA盒结合的蛋白是TBP(TATAbox binding protein),是唯一能识别TATA盒并与其结合的转录因子,是三种RNA聚合酶转录时都需要的;其他称为TBP相关因子(TBPassociated factors TAF),至少包括8种能与TBP紧密结合的因子。转录前先是TFⅡ-D与TATA盒结合;继而TFⅡ-B以其C端与TBP-DNA复合体结合,其N端则能与RNA聚合酶Ⅱ亲和结合,接着由两个亚基组成的TFⅡ-F加入装配,TFⅡ-F能与RNA聚合酶形成复合体,还具有依赖于ATP供给能量的DNA解旋酶活性,能解开前方的DNA双螺旋,在转录链延伸中起作用。这样,启动子序列就与TFⅡ-D、B、F及RNA聚合酶Ⅱ结合形成一个“最低限度”能有转录功能基础的转录前起始复合物(preintitiationcomplex, PIC),能转录mRNA。TFⅡ-H是多亚基蛋白复合体,具有依赖于ATP供给能量的DNA解旋酶活性,在转录链延伸中发挥作用;TFⅡ-E是两个亚基组成的四聚体,不直接与DNA结合而可能是与TFⅡ-B联系,能提高ATP酶的活性;TFⅡ-E和TFⅡ-H的加入就形成完整的转录复合体(图19?5),能转录延伸生成长链RNA,TFⅡ-A能稳定TFⅡ-D与TATA盒的结合,提高转录效率,但不是转录复合体一定需要的。
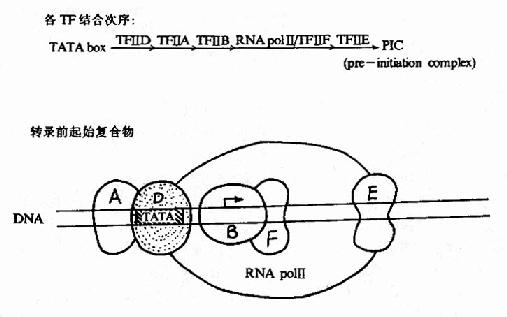
图19-15 RNA聚合酶Ⅱ转录复合体的形成示意图
以上所述是典型的启动子上转录复合体的形成,但有的真核启动子不含TATA盒或不通过TATA盒开始转录。例如有的无TATA盒的启动子是靠TFⅡ-I和TFⅡ-D共同组成稳定的转录起始复合体开始转录的。由此可以看到真核转录起始的复杂性。
不同基因由不同的上游启动子元件组成,能与不同的转录因子结合,这些转录因子通过与基础的转录复合体作用而影响转录的效率。现在已经发现有许多不同的转录因子,看到的现象是:同一DNA序列可被不同的蛋白因子所识别;能直接结合DNA序列的蛋白因子是少数,但不同的蛋白因子间可以相互作用,因而多数转录因子是通过蛋白质-蛋白质间作用与DNA序列联系并影响转录效率的(见图19-16)。转录因子之间或转录因子与DNA的结合都会引起构象的变化,从而影响转录的效率。
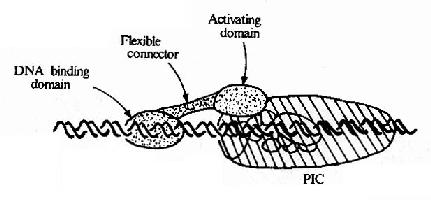
图19-16 转录因子与转录复合体相互作用模式图
图19-16所示,作为蛋白质的转录因子从功能上分析其结构可包含有不同区域,①DNA结合域(DNa binding domain),多由60-100个氨基酸残基组织的几个亚区组成;②转录激活域(activating domain),常由30-100氨基酸残基组成,这结构域有富含酸性氨基酸、富含谷氨酰胺、富含脯氨酸等不同种类,以酸性结构域最多见;③连接区,即连接上两个结构域的部分。不与DNA直接结合的转录因子没有DNA结合域,但能通过转录激活域直接或间接作用于转录复合体而影响转录效率。
与DNA结合的转录因子大多以二聚体形式起作用,与DNA结合的功能域常见有以下几种:
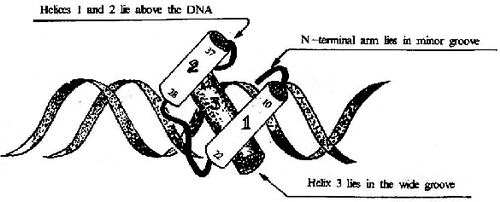
图19-17 HTH结构及其与DNA的结合
①螺旋转角螺旋(helixturnhelix, HTH)及螺旋-环-螺旋(helixloophelix,HLH) 这类结构至少有两个α螺旋,其间由短肽段形成的转角或环连接,两个这样的motif结构以二聚体形式相连,距离正好相当于DNA一个螺距(3.4nm),两个α螺旋刚好分别嵌入DNA的深沟(图19-17)。
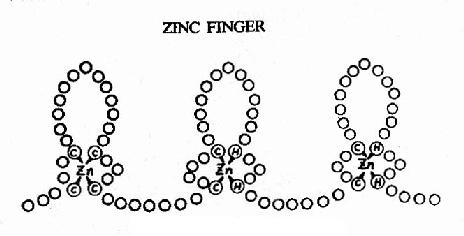
图19-18 蛋白质的锌指结构
②锌指(zinc finger) 其结构如图19-18所示,每个重复的“指”状结构约含23个氨基酸残基,锌以4个配价键与4个半胱氨酸、或2个半胱氨酸和2个组氨酸相结合。整个蛋白质分子可有2?个这样的锌指重复单位。每一个单位可以其指部伸入DNA双螺旋的深沟,接触5个核苷酸。例如与GC盒结合的转录因子SP1中就有连续的3个锌指重复结构。
③碱性-亮氨酸拉链(basic leucine zipper, bZIP),该结构的特点是蛋白质分子的肽链上每隔6个氨基酸就有一个亮氨酸残基,结果就导致这些亮氨酸残基都在α螺旋的同一个方向出现。两个相同结构的两排亮氨酸残基就能以疏水键结合成二聚体,该二聚体的另一端的肽段富含碱性氨基酸残基,借其正电荷与DNA双螺旋链上带负电荷的磷酸基团结合。若不形成二聚体则对DNA的亲和结合力明显降低。在肝脏、小肠上皮、脂肪细胞和某些脑细胞中有称为C/EBP家族的一大类蛋白质能够与CAAT盒和病毒增强子结合,其特征就是能形成bZIP二聚体结构。
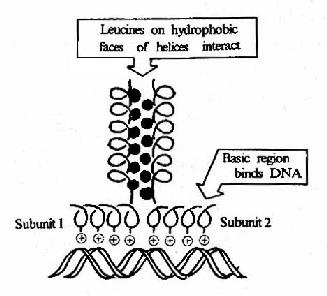
图19-19 碱性亮氨酸拉链结构及其与DNA的结合
从上述可见:转录调控的实质在于蛋白质与DNA、蛋白质与蛋白质之间的相互作用,构象的变化正是蛋白质和核酸“活”的表现。但对生物大分子间的辨认、相互作用、结构上的变化及其在生命活动中的意义,人们的认识和研究还只在起步阶段,其中许多内容甚至重要的规律我们可能至今还一无所知,有待于努力探索。