转录是从DNA分子的特定部位开始的,这个部位也是RNA聚合酶全酶结合的部信这就是启动子。为什么RNA聚合酶能够仅在启动子处结合呢?显然启动子处的核苷酸序列具有特殊性,为了方便,人们将在DNA上开始转录的第一个碱基定为+1,沿转录方向顺流而下的核苷酸序列均用正值表示;逆流而上的核苷酸序列均用负值表示。
对原核行物的100多个启动子的序列进行了比较后发现;在RNA转录起始点上游大约-10bp和-35bp处有两个保守的序列,在-10bp附近,有一组5’-TATAATpu的序列,这是Pribnow首先发现的称为Pribnow框,RNA聚合酶就结合在互部位上。-35bp附近,有一组5’-TTGACG-的序列;已被证实与转录起始的辨认有关,是RNA聚合酶中的δ亚基识别并结合的位置。-35序列的重要性还在于在很大程度上决定了启动子的强度。
由于RNA聚合酶分子很大,大约能覆盖70bp的DNA序列,因此酶分子上的一个适合部位就能占据从-35到-10序列区域(图17-4)。
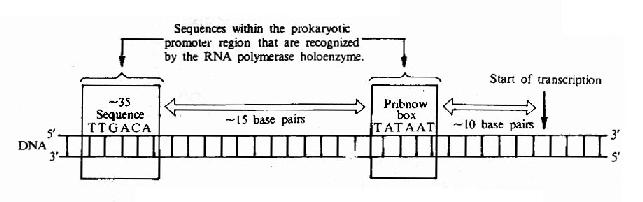
图17-4 Structure of the prokaryotic promoter region.
真核生物的启动子有其特殊性,真核生物有三种RNA聚合酶,每一咱都有自己的启动子类型。以RNA聚合酶Ⅱ的启动子结构为例,人们比较了上百个真核生物RNA聚合酶Ⅱ的启动子核苷酸序列伯发现;在-25区有TATA框,又称为Hogness框或Goldberg-Hogness框。其一致序列为T28A97A93A85A63T37A83A50T37,基本上都由A,T碱基所组成,离体转录实验表明,TATA框决定了转录起点的选择,天然缺少TATA框的基本可以从一个以上的位点开始转录。在-75区有CAAT框,其一致的序列为GGTCAATCT。有实验表明CAAT框与转录起始频率有关,例如缺失GG,兔子的β珠蛋白基因转录效率只有原来的12%(图17-5)。
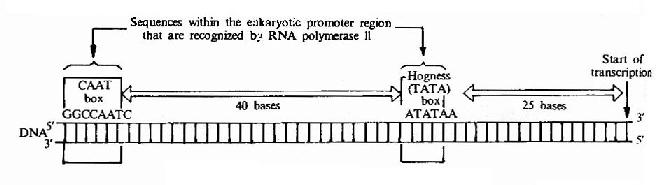
图17-5 Eukaryotic gene promoter sequences
除启动子外,真核生物转录起始点上游处还有一个称为增强子的序列,它能极大地增强启动子的活性,它的位置往往不固定,可存在于启动子上游或下游,对启动子来说它们正向排列和反向排列均有效,对异源的基因也起到增强作用,但许多实验证实它仍可能具有组织特异性,例如免疫球蛋白基因的增强子只有在B淋巴细胞内活性最高,胰岛素基因和胰凝乳蛋白酶基因的增哟子也都有很高的组织的特异性。