氨基酸在进行合成多肽链之前,必须先经过活化,然后再与其特异的tRNA结合,带到mRNA相应的位置上,这个过程靠氨基酰tRNA合成酶催化,此酶催化特定的氨基酸与特异的tRNA相结合,生成各种氨基酰tRNA.每种氨基酸都靠其特有合成酶催化,使之和相对应的tRNA结合,在氨基酰tRNA合成酶催化下,利用ATP供能,在氨基酸羧基上进行活化,形成氨基酰-AMP,再与氨基酰tRNA合成酶结合形成三联复合物,此复合物再与特异的tRNA作用,将氨基酰转移到tRNA的氨基酸臂(即3'-末端CCA-OH)上(图18-5和图18-6)。
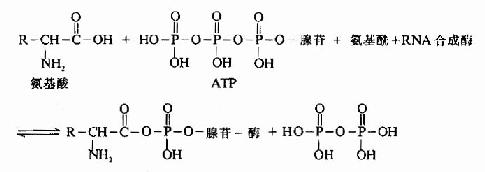
图18-6 氨基酰-tRNA的生成
原核细胞中起始氨基酸活化后,还要甲酰化,形成甲酰蛋氨酸tRNA,由N10甲酰四氢叶酸提供甲酰基。而真核细胞没有此过程。
前面讲过运载同一种氨基酸的一组不同tRNA称为同功tRNA。一组同功tRNA由同一种氨酰基tRNA合成酶催化。氨基酰tRNA合成酶对tRNA和氨基酸两者具有专一性,它对氨基酸的识别特异性很高,而对tRNA识别的特异性较低。
氨基酰tRNA合成酶是如何选择正确的氨基酸和tRNA呢?按照一般原理,酶和底物的正确结合是由二者相嵌的几何形状所决定的,只有适合的氨基酸和适合的tRNA进入合成酶的相应位点,才能合成正确的氨酰基tRNA。现在已经知道合成酶与L形tRNA的内侧面结合,结合点包括接近臂,DHU臂和反密码子臂(图18-7)。
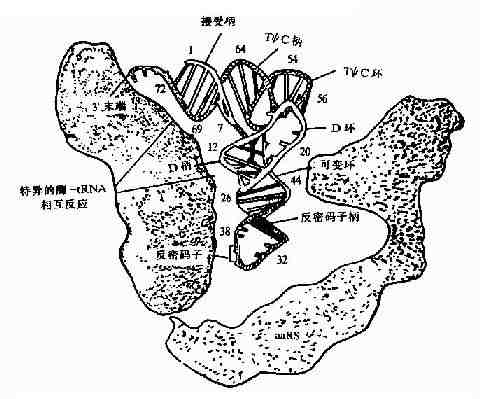
图18-7 氨基酰-tRNA合成酶与tRNA的相互作用,可见氨酸接受柄、
D柄、反密码子和可变环与酶反应
乍看起来,反密码子似乎应该与氨基酸的正确负载有关,对于某些tRNA也确实如此,然而对于大多数tRNA来说,情况并非如此,人们早就知道,当某些tRNA上的反密码子突变后,但它们所携带的氨工酸却没有改变。1988年,候稚明和Schimmel的实验证明丙氨酸tRNA酸分子的氨基酸臂上G3:U70这两个碱基发生突变时则影响到丙氨酰tRNA合成酶的正确识别,说明G3:U70是丙氨酸tRNA分子决定其本质的主要因素。tRNA分子上决定其携带氨基酸的区域叫做副密码子。一种氨基酰tRNA合成酶可以识别以一组同功tRNA,这说明它们具有共同特征。例如三种丙氨酸tRNA(tRNAAlm/CUA,tRNAAim/GGC,tRNAAin/UGC都具有G3:U70副密码子。)但没有充分的证据说明其它氨基酰tRNA合成酶也识别同功tRNA组中相同的副密码子。另外副密码子也没有固定的位置,也可能并不止一个碱基对。