G蛋白偶联型受体包括多种神经递质、肽类激素和趋化因子的受体,在味觉、视觉和嗅觉中接受外源理化因素的受体亦属G蛋白偶联型受体。这类受体在结构上均为单体蛋白,氨基末端位于细胞外表面,羧基末端在胞膜内侧。完整的肽链要反复跨膜七次(图21-10),因此亦有人将此类受体称为七次跨膜受体。由于肽链反复跨膜,在膜外侧和膜内侧形成了几个环状结构,它们分别负责与配体(化学、物理信号)的结合和细胞内的信号传递。其胞浆部分可以与一种GTP结合蛋白(简称G蛋白)相互作用,这种G蛋白是该信号传递途径中的第一个信号传递分子,这也是这类受体被称为G蛋白偶联型受体的原因。
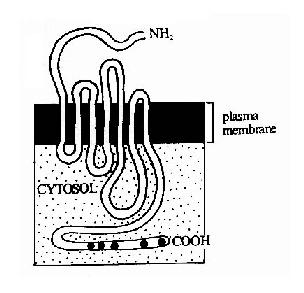
图21-10 七次跨膜受体-G蛋白偶联型受体跨膜结构示意图
G蛋白偶联受体的信号传递过程包括(1)配体与受体结合,(2)受体活化G蛋白;(3)G蛋白激活或抑制细胞中的效应分子;(4)效应分子改变细胞内信使的含量与分布,(5)细胞内信使作用于相应的靶分子,从而改变细胞的代谢过程及基因表达等功能。本节将逐一介绍这一过程的主要环节。
1.G蛋白的循环或活化(G Protein Cycle)
G蛋白偶联型受体的信号转导途径中的第一个信号传递分子是G蛋白,其活化过程称为G蛋白循环。
G蛋白以α、βγ亚基三聚体的形式存在于细胞质膜内侧。α亚基已发现有20余种,分子量为36~52kd。α亚基具有多个活化位点,其中包括可与受体结合并受其活化调节的部位、与βγ亚基相结合的部位、GDP或GTP结合部位以及与下游效应分子相互作用的部位等等。α亚基还具有GTP酶活性。α亚基结合GDP时是无活性状态,而与GTP结合时则为有活性状态,GTP的水解又使其返回无活性状态。
G蛋白中的β和γ亚基亦有数种,但不及α亚基种类多。在细胞内,β和γ亚基形成紧密结合的二聚体,只有在蛋白变性条件下方可解离,因此可以认为它们是功能上的单体。βγ亚基的主要作用是与α亚基形成复合体并定位于质膜内侧。近年来的研究表明,βγ亚基亦可作用于其下游效应分子。
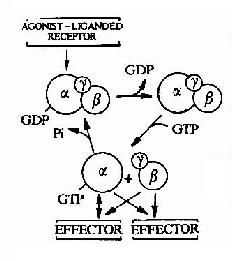
图21-11 G蛋白循环示意图
G蛋白循环的具体过程可见图21-11。当物理或化学信号刺激受体时,受体活化G蛋白使之发生构象改变。α亚基与GDP的亲和力下降,结合的GDP为GTP所取代。α亚基结合了GTP后即与βγ亚基发生解离,成为活化状态的α亚基。活化了的α亚基此时可以作用于下游的各种效应分子。这种活化状态将一直持续到GTP被α亚基自身具有的GTP酶水解为GDP。一旦发生GTP的水解,α亚基又再次与βγ亚基形成复合体,回到静止状态,重新接受新的化学信号。
由于G蛋白的种类不同,因此G蛋白可以作用于不同的效应分子,或对同一效应分子进行不同的调节。
2.效应分子及细胞内信使
G蛋白活化之后,可作用于腺苷酸环化酶和磷脂酶C等效应分子(Effector)上。有的α亚基(Gs)可以激活腺苷酸环化酶;有的α亚基(αi)可以抑制腺苷酸环化酶。腺苷酸环化酶催化ATP生成环状AMP(cAMP)的反应,因此细胞内的cAMP水平在配体与受体结合后,可受G蛋白α亚单位的作用而升高或降低,从而将细胞外信号转变为细胞内信号。这种细胞内信号可再作用于下游分子。这种细胞内信号的传递方式是G蛋白偶联型受体传递信号的主要方式,这些细胞内信号分子被称为细胞内信使。细胞内信使亦被称为第二信使。已知的细胞内信使包括cAMP、cGMP、甘油二酯(DAG)、IP3、和Ca2+等等(图21-12、13、14)。G蛋白的α亚基种类、其作用的效应分子及所调节的细胞内信使可参见表21-3。
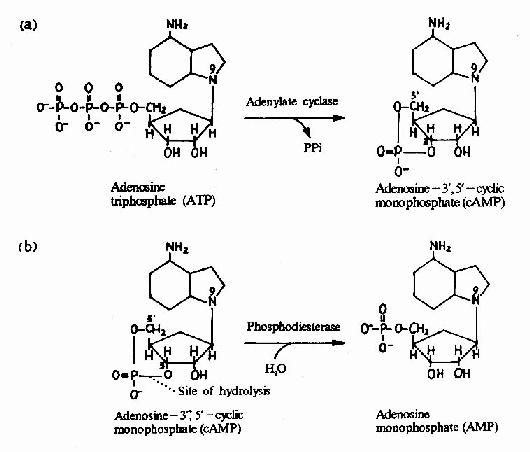
图21-12 cAMP的生成与水解
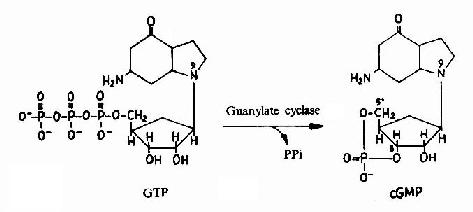
图21-13 cGMP的生成
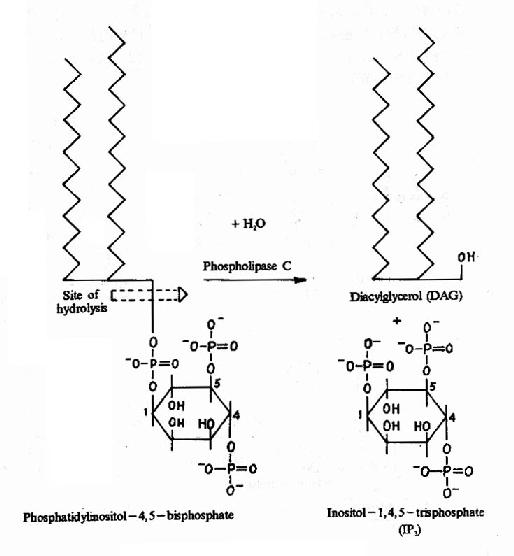
图21-14 DAG和IP3的生成
细胞内信使一般具有以下三个特点:(1)多为小分子,且不位于能量代谢途径的中心;(2)在细胞中的浓度或分布可以迅速地改变;(3)作为变构效应剂可作用于相应的靶分子,已知的靶分子主要为各种蛋白激酶。
表21-3 G蛋白的α亚基及其效应分子
| Ca种类 | 效应分子 | 细胞内信使 | 靶分子 |
| as | 腺苷酸环化酶活性急↑ | cAMP↑ | 蛋白激酶A活性↑ |
| ai | 腺苷酸环化酶活性↓ | cAMP↓ | 蛋白激酶A活性↓ |
| aq | 磷脂酶C活性↑ | Ca2+IP↑3DAG | 蛋白激酶C活化↑ |
| at | cGMP磷脂二酯酶↑活性 | cGMP↓ | Na+通道关闭 |
cAMP是第一个被发现的细胞内信使,催化它生成的腺苷酸环化酶为一重要的Gαi和Gαs的效应分子。cAMP是很多激素的细胞内信使。另一类重要的细胞内信使是在磷脂酰肌醇特异性磷脂酶C作用下,由PIP2(二磷酸磷脂酰肌醇)水解生成的三磷酸肌醇(IP3)和甘油二酯(DAG)。
需要指出的是,除G蛋白偶联型受体在其信号转导过程中需细胞内信使作为信号的传递者外,细胞内还存在受其它的信号转导方式调控的细胞内信使。九十年代以来,越来越多的以小分子物质作为细胞内信使参与细胞功能调控的过程得以阐明。
G蛋白偶联型受体在G蛋白介导下的信号传递过程可见图21-15和图21-16。
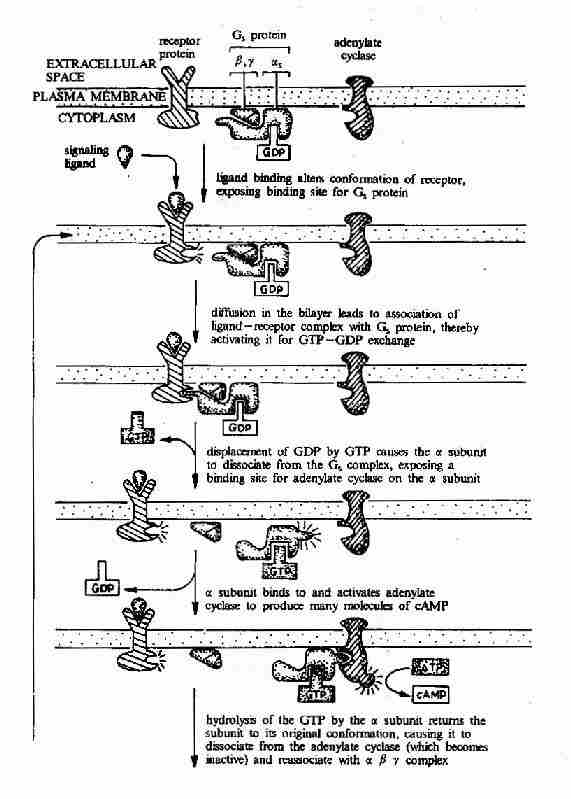
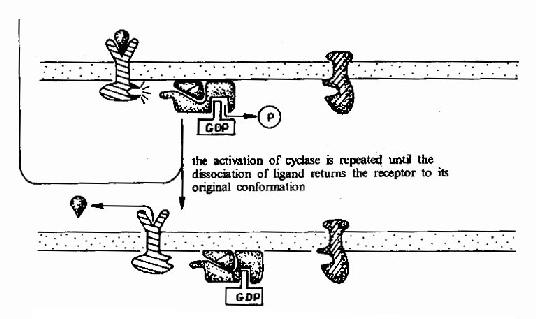
图21-15 G蛋白偶联型受体-G蛋白-腺苷酸环化酶信号转导途径示意图
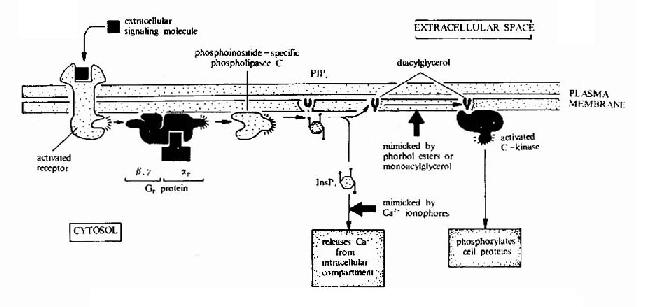
图21-16 G蛋白偶联型受体-G蛋白-磷脂酶C信号转导途径示意图
3.细胞内信使作用的主要靶分子
活化的Gα可作用于相应的效应分子,从而使相应的细胞内信使浓度发生迅速的改变。这些细胞内的信使可分别作用于相应的靶分子,从而使得细胞中的各种酶类和蛋白分子的活性发生改变。这些细胞内信使所作用的靶分子主要为各种蛋白激酶(表21-3)
(1)蛋白激酶A
cAMP作用于cAMP依赖性蛋白激酶(cAMP-dependent Protein Kinase,简称为cAPK),亦称为蛋白激酶A(Protein Kinase A, PKA),目前后一种命名较为公认。cAMP可以作为该激酶的变构激活剂,使无活性的蛋白激酶A转变为有活性的蛋白激酶A(详见第十一章,图11-4)。活化了的蛋白激酶A可作用于多种与糖脂代谢相关的酶类、一些离子通道和某些转导因子,使它们发生磷酸化并改变其活性状态。
(2)蛋白激酶G
细胞内的另一种环核苷酸信使为环鸟苷酸-cGMP。cGMP作用于cGMP依赖性蛋白激酶-cGMPdependent Protein Kinase, cGPK),亦称为蛋白激酶G(Protein Kinase G、PKG)。与PKA一样,PKG是目前较为公认的命名。蛋白激酶G以cGMP作为变构效应剂,在脑和平滑肌中含量较丰富。1997年,人们发现,PKG的基因突变与果蝇的觅食行为有关。我们可以推测,PKG很有可能在神经系统的信号传递过程中具有重要作用。
(3)蛋白激酶C
另外一些重要的细胞内信使还包括磷脂酰肌醇的衍生物如DAG、PIP3(三磷酸磷脂酰肌醇)、磷脂酰胆碱的衍生物、鞘磷脂的衍生物以及Ca2+等等。这些小分子信使的一个重要靶分子是蛋白激酶C(Protein KinaseC、PKC)。
PKC有多种同工酶形式,均以希腊字母排列,有PKCα、PKCβI、PKCβⅡ和PKCγ等等。不同的同功酶在结构和组织分布上各有不同,其对辅助因子(包括上述细胞内小分子信使)的需求亦有差别,并且对底物有选择性。
PKC在细胞的生长分化的调控中及其它多种细胞功能上具有关键性的调节作用,是一类非常重要的信号转导分子。细胞信号转导过程中的多条途径都可以导致PKC的活化。已经有很多实验研究证明,PKC的抑制剂可以使细胞失去对生长分化刺激信号的反应,表明这些功能都依赖于PKC的调控。例如,肥大细胞的脱颗粒反应(释放出大量组织胺等血管活性物质)是机体变态反应的重要形式之一。体外实验表明,如果用PKC抑制剂预先处理细胞,细胞就不会对刺激信号再发生脱颗粒反应。如果用改变细胞膜通透性的方式使胞内的PKC漏出,细胞也会失去发生脱颗粒反应的能力,此时若再加入PKC使之回到细胞中,则又可恢复细胞的脱颗粒反应。其它很多类似的实验亦表明,细胞的很多其它功能也受到PKC的调控。