实验观察表明:当色氨酸达到一定浓度,但还没有高到能够活化R使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,而且产生的酶量与色氨酸浓度呈负相关。仔细研究发现这种调控现象与色氨酸操纵元特殊的结构有关。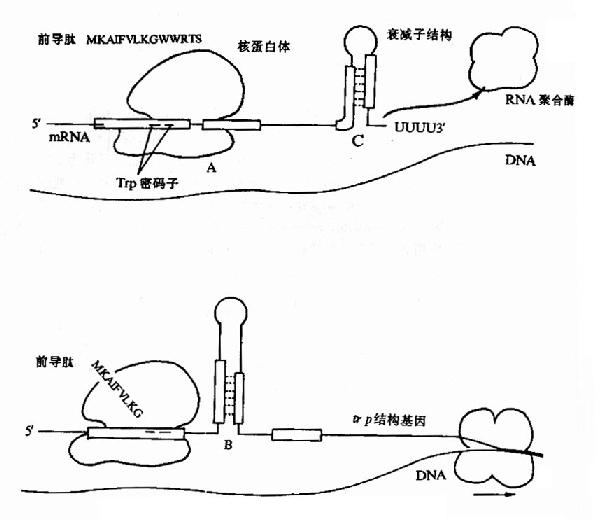
图19-11 色氨酸操纵元中的衰减子结构及其调控示意图
在色氨酸操纵元Ptrp-o与第一个结构基因trpE之间有162bp的一段先导序列(leadingsequence,L)实验证明当色氨酸达一定浓度时,RNA聚合酶的转录会终止在这里。这段序列中含有编码由14个氨基酸组成的短肽的开放读框,其序列中有2个色氨酸相连,在此开放读框前有核糖体识别结合位点(RBS)序列,提示这段短开放读框在转录后是能被翻译的。在先导序列的后半段含有3对反向重复序列(图19?1中A、B及C),在被转录生成mRNA时都能够形成发夹式结构,但由于B的序列分别与A和C重叠,所以如果B形成发夹结构,A和C都不能再形成发夹结构;相反,当A形成发夹结构时,B就不能形成发夹结构,却有利于C生成发夹结构。C后面紧跟一串A(转录成RNA就是一串U),C实际上是一个终止子,如果转录mRNA时它形成发夹结构,就能使RNA聚合酶停止转录而从mRNA上脱离下来。
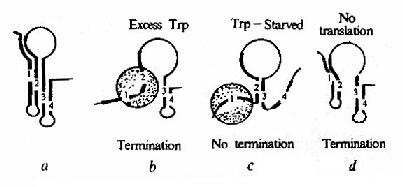
图19-12 三种不同情况下A、B、C形成发夹结构的状态
在色氨酸未达到能起阻遏作用的浓度时,从Ptrp起始转录,RNA聚合酶沿DNA转录合成mRNA,同时核糖体就结合到新生成的mRNA核糖体结合位点上开始翻译。当色氨酸浓度低时,生成的tRNAtrp色氨酸量就少,能扩散到核糖体mRNA形成的翻译复合体中供给合成短肽的几率低,使核糖体沿mRNA翻译移动的速度慢,赶不上RNA聚合酶沿DNA移动转录的速度,这时核糖体占据短开放读框的机会较多,使A不能生成发夹结构,于是B就形成发夹结构,阻止了C生成终止信号的结构,RNA聚合酶得以沿DNA前进,继续去转录其后trpE等基因,trp操纵元就处于开放状态。当色氨酸浓度增高时,tRNAtrp色氨酸浓度随之升高,核糖体沿mRNA翻译移动的速度加快,占据到B段的机会增加,B生成发夹结构的机会减少,C形成终止结构的机会增多,RNA聚合酶终止转录的的几率增加,于是转录减弱。如果当其他氨基酸短缺(注意:短开放读框编码的14肽中多数氨基酸能由环境充分供应的机会是不多的)或所有的氨基酸都不足时,核糖体翻译移动的速度就更慢,甚至不能占据A的序列,结果有利于A和C发夹结构的形成,于是RNA聚合酶停止转录,等于告诉细菌:“整个氨基酸都不足,即使合成色氨酸也不能合成蛋白质,不如不合成以节省能量”。
由此可见,先导序列起到随色氨酸浓度升高降低转录的作用,这段序列就称为衰减子attenuator)。在trp操纵元中,对结构基因的转录阻遏蛋白的负调控起到粗调的作用,而衰减子起到细调的作用。细菌其他氨基酸合成系统的许多操纵元(如组氨酸、苏氨酸、亮氨酸、异亮氨酸、苯丙氨酸等操纵元)中也有类似的衰减子存在。