TCR/CD3复合体中的两个多态型亚单位(TCRαβ或TCRγδ)主要功能是识别结合MHC分子的抗原,而胞浆区非常短;CD3分子的主要功能是参与TCR/CD3复合体的装配和稳定以及信号转导(表8-1)。CD3分子亚单位的胞浆内部分含有一个共同的序列,即D/EX2YX2L/IX8YX2L/I,其中含有两个YXXL/I结构。由于其序列同淋巴细胞抗原识别后淋巴细胞的活化及信号转导关系密切,因而把此序列称为抗原识别活化基序(antigen recognition acivation motifs,ARAMS)其中含有两个YXXL/I结构。由于其序列同淋巴细胞抗原识别后淋巴细胞的活化及信号转导关系密切,因而把此序列称为抗原识别活化基序(antigen recognition activation motifs,ARAMS)。其中CD3γ、ε、δ各含一个ARAM,η链含二个,ζ链含三个。此外,在B细胞受体(b cell receptor ,BCR)α链(Igα、CD79a)和β链(Igβ、CD79b)、FcγRⅢ以及Fcεr I的γ链、FcεR I β链等中也含有类似结构的ARAM。经突变及嵌合分子等研究证实,ARAM是TCR信号转导的结构基础,单独分离的ARAM能够转TCR介导的信号。TCR与抗原结合后可激活一些PTKs,包括TCR相连的p59fyn、ZAP-70(ζassociated protein-70)、CD4/CD8相连的p56lck以及其它src相关的PTKs(图8-2),随后引起多种底物的酪氨酸磷酸化。
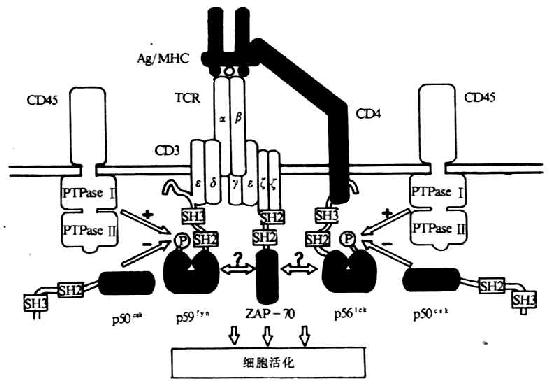
图8-2 TCR与抗原结合后导致一些激酶的活化
目前证实TCR激活的PTKs底物有原癌基因产物Vav、42kDa的微管相关蛋白激酶(Mi-crotubule-associated protein kinase,MAPK)、PLCγ1及CD3ζ链(图8-3)。这些磷酸化的蛋白在信号转导中具有重要作用。
TCR/CD3复合体根据结构特点可分为三组:(1)两个多态型亚单位即TCRβ或TCRγδ,属免疫球蛋白超家族成员,各含有一个V区和一个C1区;(2)CD3γ、δ和ε链,亦属免疫球蛋白超家族成员,各含有一个C2区;(3)ζ和η链,胞膜外区很短,不属于免疫球蛋白超家庭的成员(图8-4)。
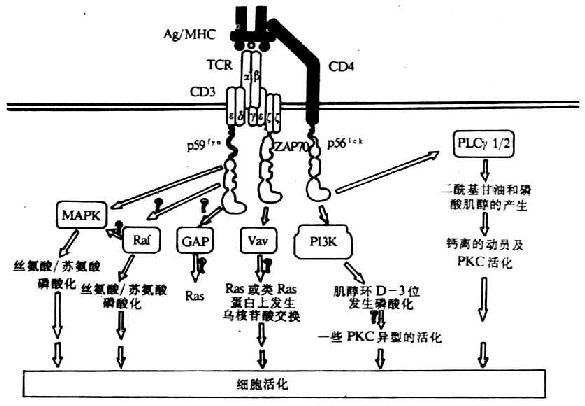
图8-3 TCR同抗原结合后所引起的PTKs底物磷酸化