一、C1抑制物
C1抑制物(C1INH)是血清中高度糖基化的一种蛋白质,含糖量高达35-49%。最初由Ranoff和lepow(1957)所发现,称其为C1酯酶抑制剂,与引同时Schultze等则将其称为α2神经氨酸糖蛋白。C1INH为一单链分子,由478个氨基酸残基组成,分子量为104kDa,由478个氨基酸残基组成,分子量为140kDa,链内有两对二硫键(图5-13)。C1INH调节的主要方式是,与活化的C1r或C1s结合形成稳定的复合物而导致C1丝氨酸蛋白酶失活。其作用机理是,C1INH通过提供一个酷似C1r或C1s的正常底物的序列为“锈铒”(“bait”),被c 1r或C1s裂解暴露出一个活性部位,然后再与C1r或C1s结合形成共价的酯键而发挥抑制作用。此外,C1INH还可防止在缺乏抗体时,C1以很低但仍有一定速率出现的自发激活。正常情况下,血液中的大多数C1可被7倍于其克分子浓度的C1INH所结合,以防止C1由于构象改变而引起的自发激活。但C1同抗原、抗体复合物的结合,可使C1从C1INH的抑制作用中而获释。除上述作用外,C1INH还可抑制凝血因子Ⅻa、Ⅺa、激肽释放酶及纤溶酶,因而其在凝血、激肽和纤溶系统中也有重要的调节作用。
经对C1INH cDNA序列的分析发现,C1INH与其它几种丝氨酸蛋白酶抑制物(serpin)超家族成员(α1抗胰蛋白酶、α1抗糜蛋白酶及抗凝血酶Ⅲ等)约有30%的氨基酸同源性,物别是在C端的120个氨基酸。C1INH的编码基因定位于第11号染色体的短臂11.2~长臂13亚区。C1INH先天性缺陷时,可导致遗传性血管神经性水肿(hereditary angioneurotic edema,HAE)。近年报道应用C1INH浓缩剂防治HAE效果良好,但尚未广泛用于临床。
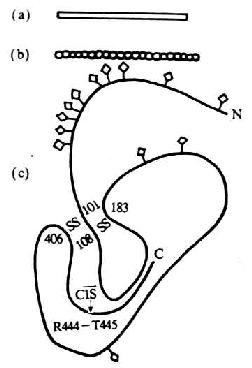
图5-13 C1 INH分子的结构模式图
注:(a)为电镜观察模式图
(b)为中子散射模式图
(c)为园二色谱分析的二级结构模式图
二、C4结合蛋白
C4结合蛋白(C4bp)是一种含量丰富的可溶性血清糖蛋白,分子量为550kDa,1977年由Ferreira等所报道。其分子结构模式现多以Dahlback等(1983)描述的“蜘蛛样”(spiderlike)结构来分析其结构及功能。C4bp由8个亚单位组成,电镜下观察形似蜘蛛,其中有7条分子量相同(均为70kDa)的长链(α链),由549个氨基酸残基组成,链间以二硫键相连结,并共同氨基酸残基组成,链间以二硫键相连结,并共同连结于一中心体。α链含有8个SCR,其N端是其头部,为与C4b相结合的部位,可能位于第332-395位氨基本酸残基。第8条链(β链)较短(45kDa)由235个氨基酸残基组成,含有4个SCR为与蛋白S(PS)相结合的部位(图5-14)。C4bp以两种方式抑制补体的活化。第一种方式是,通过其与C2竞争C4b,而从c 4b2a中取代C2a,并通过其与C4b的结合而阴止剩余的C2同C4b结合,由此抑制C3转化酶的形成。C4bp与C4b的结合能力与细胞表面C4b分子的数成正比,且较C2同C4b的结合能力高27倍。第二种方式是,C4bp作为I因子的一种辅助因子,促进I因子对C4b的裂解。有C4bp存在时,I因子可将C4b的a`链完全裂解;无C4pb时,I因子的裂解作用不完全。其与I因子结合的活性部位位于第177-322位氨基酸残基区域。PS与C4pb的第8条链(β链)结合,不影响C4bp同C4b的结合,但可延长C4bp的半衷期,从而强化C4bp的抑制作用。
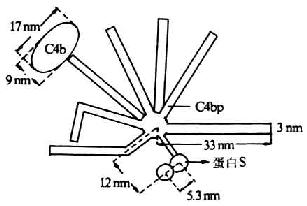
图15-14 C4bp的结构(模式图)
C4bp基因定位于人的第1号染色体长臂32区,同CR1、CR2、H因子、MCP及DAF等的基因相连锁,并均含有数目不同的SCR。
三、促衰变因子(CD55)
促衰变因子(decayaccelerating factor,DAF)是Nicholson-Weller等(1981)用正丁醇提取后,再以层析法从人和豚鼠红细胞基质中纯化的一种膜蛋白。因其具有促进C3转化酶衷变的活性故名。经在还原条件下做SDS-PAGE并以过碘酸-Schiff试剂染色表明,纯化的DAF为单链膜糖蛋白。人和豚鼠的DAF分了量不同分别为70kDa和60kDa。现已按白细胞分化抗原将其归为CD55。Davitz等(1986)通过用磷脂酰肌醇(PI)特异性的磷脂酶C(PI-PLC)处理人外周血细胞可释放DAF的事实探明,DAF是经糖磷脂酰肌醇(glycosylphodphatidylinositol,GPI)锚而固定于细胞膜中的。即糖蛋白的C末端共价结合于含PI的糖磷脂上,再经PI插入细胞膜脂质双层的外层小叶中。研究表明,膜DAF的迁移率接近于膜类脂的迁移率,比大多数膜蛋白迁移率高一个数量级。认为这有助于促进数目有限的膜DAF分子与细胞表面大量的C3b或C4b片段接触。另外DAF的糖磷脂酰肌醇结构还可能具有转导细胞信号的作用。
除Nicholson-Weller等证实的分子量为70kDa的膜DAF外,Kinoshita等(1987)用Western blotting在人红细胞表面还检出分子量为140kDa的一种膜DAF,称其为DAF-2。DAF-2在膜上的数目不足70kDa膜DAF的1/10,但也有促进C3b转化酶衰变的活性,也含有GPI锚结构。由于DAF-2的分子量较70kDa的膜DAF大一倍,提示其为膜DAF的二聚体,但用二巯基乙醇或以SDS使基变性,都不能将DAF-2裂解成两个成分,故DAF-2的精确结构仍有待进一步阐明。另外,近年应用两位点RIA测定法,在血浆、尿液、泪液、唾液、滑膜液和脑脊液,以及组织培养上清中均检出可溶性的DAF(sDAF),水平的40-400ng/ml范围。并发现尿液中的sDAF分子量略低于红细胞上膜DAF的分子量,疏水性也较膜DAF小,其抑制细胞表面C3转化酶内在装配的活性较膜DAF约低100倍,但仍具有促进已形成的C3转化酶衰变的作用,效能类似于C4bp。
膜DAF广泛分布于各种血细胞及其他各处的细胞上,包括红细胞、粒细胞、单核细胞、淋巴细胞(T、B)、血小板、骨髓单个核细胞、红细胞的始祖细胞,角膜、结合膜、消化道粘膜、外分泌腺、肾小管、膀胱、子宫粘膜、胞膜、心包及滑膜的上皮细胞,精子,以及培养的脐静脉内皮细胞上。但NK细胞上则缺如。阵发性血红蛋白尿(paroxysmal nocturnalhemohlobulinuria,PNH)病人的红细胞上也缺少DAF,并以缺乏程度将该病分为三个型。PNH病人的红细胞对补体介导的溶血作用高度敏感,就是由于红细胞上缺乏DAF及基它含GPI锚的分子而引起的。DAF带有Cromer抗原。极少数称之为Inaba或IFC缺乏的Cromer相关抗原的个体也缺乏DAF。
DAF生物学活性及生理功能已虱到充分证实。它可保护宿主细胞免遭补体介导的溶解破坏。其作用机理是,DAF不仅可阻止经典或替代途径C3和C5转化酶的装配,并且可通过诱导催化单位C2a或Bb的快速解离而使已形成的C4、C5转化酶失去稳定性,从而抑制补体攻击单位的活化(图5-15)。DAF的这种抑制作用仅限于直接结合在细胞上的C3、C5转化酶,也即DAF不抑制靶细胞上正常的补体激活剂如微生物和免疫复合物。但DAF不能作为I因子裂解C3b和C4b的辅因子而发挥作用。另外,DAF虽不能阻止C2和B因子(分别通过与C4b或C3b结合)与细胞的最初结合,但却可使C2a或Bb由它们结合的部位解离出来,以而阻止C3转化酶的装配。
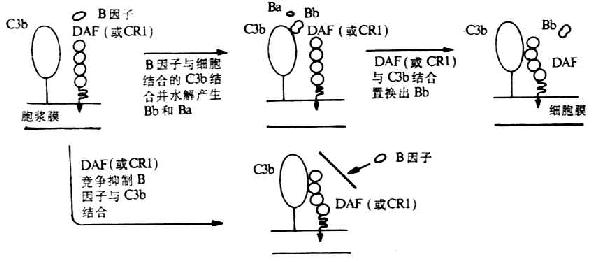
图5-15 DAF抑制替代途径中C3转化酶形成的的机理
编码人DAF的基因位于第1号染色体的长臂上32区一个800kb片段内,与其它几种补体激活调节剂(RCA)的基因紧密连锁,排列顺序依次为:MCP-CR1-CR2-DAF-C4bp。其中DAF基因的长度约为C4bp。其中DAF基因的长度约为35kb,以限制性酶谱分析表明为一单拷贝基因。在DAF基因的非编码区还有3个限制性酶切片段长度多态性(RFLP)结构,两个为HindⅢ酶切位点,1个为Bamh Ⅰ酶切位点。DAF的cDNA已克隆成功,并进行了核苷酸和氨基酸序列分析。关于DAf cDNA和膜糖蛋白的结构见图5-16。DAF的cDNA结构从5`端开始依次为:信号肽区、四个SCR(长度1143bp)、富含丝氨酸与苏氨酸(S/T)的区(约70个氨基酸)及疏水区,最后终止于3`端的poly(A)。由cDNA推导的氨基酸序列得出DAF蛋白由381个氨基酸所组成,包括34个氨基酸的信号肽,富含S/T的区为DAF中大多数延伸的O-连接的糖基化部位。SCR中有1上N-连接的的糖基化部位。C末端的疏水区在翻译后被糖磷脂所取代,为DAF分子与细胞膜相结合的部位。
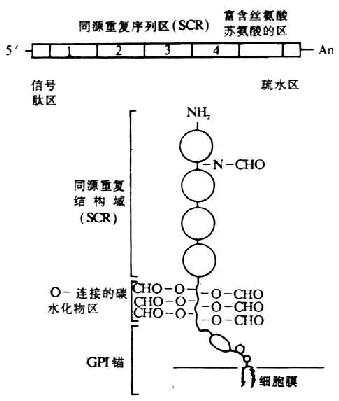
图5-16 DAF cDNA和其膜糖蛋白的结构
四、膜辅蛋白(CD46)
膜辅蛋白(membranecofactor protein,MCP)由Cole等(1985)应用C3b亲和层析在外周血淋巴细胞上发现的一种膜蛋白。由于其奇特的电泳特征,起初被命名为gp45-70,但进一步研究发现,它对I因子介导的对C3b和C4b的裂解有辅助活性,故更命为MCP。鉴于MCP的多克隆抗体与促衷变因子(DAF)、H因子或CR1均不起反应,从而认为它是补体系统的一种新的调节蛋白,在第四届国际白细胞分型讨论会上将其命名为CD46。
MCP为一单链穿膜糖蛋白,分子量45-70kDa,属于RCA基因簇的成员。也通过GPI锚固定于细胞上。MCP的细胞分布甚广,包括粒细胞、血小板、T细胞(Th、Ts、Tc)、B细胞、NK细胞、造血细胞系、成纤维细胞、表皮细胞、内皮细胞及星状胶质细胞等。但不同类型的细胞上表达的数量有所不同。外周血单个核细胞和粒细胞为1万个/细胞,造血细胞系为2-5万个/细胞,Hela和Hep-2细胞分别为10万个/细胞和25万个/细胞。这种表达数量的差异,可能反映了MCP在正常细胞和肿瘤细胞上不同的分化和活化状态。另外,由于不同细胞上表达的MCP不同,因此可调节两条激活途径的C3转化酶形成。这一点尤其重要,因为C3慢速运转(tickover)的机制,能够连续不断的产生C3b与C4b,有可能形成越来越多的C3转化酶。而由于大多数正常细胞上有高水平的MCP,因此可保护正常细胞免遭补体介导的损伤。相反,许多异物颗粒和致病微生物则缺乏MCP,这样沉积在它们表面的C3b便可得以保持其活化;且C3b与B因子结合的亲和力高于与H因子结合的亲和力,从而促进c 3bBb复合物的形成,导致在这些异物颗粒上补体有效地活化,最终将其破环清除。此外,细胞表面唾液酸的含量与其结合H因子和B因子的亲和力有关,唾液酸含量较高的细胞与H因子的结合亲和力超过与B因子的结合亲和力,唾液酸含量较低的细胞则与此相反。许多细菌表面涶液酸的含量低于哺乳类动物和人的细胞,因此侵入机体后易同B因子结合而导致补体替代途径的激活。
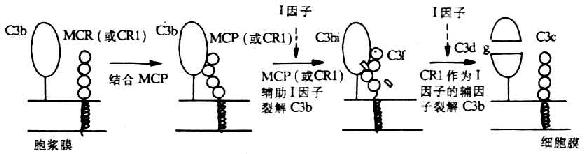
图5-17 MCP辅助I因子裂解C3b的机理
MCP作为一种补体调节蛋白具有重要的生物学功能。在低离子强度下,它可与C3b或C3bi结合,但较CR1的亲和力低。MCP的主要功能是其辅因子活性。即可与C3b或C4b结合而促进I因子对C3b和C4b的裂解灭活(图5-17),从而保护自身宿主细胞免遭补体介导的溶解破环。有人将MCP的这种作用称为内源性辅因子活性。Seya等还发现MCP具有增强C3转化酶的活性,尤其对替代途径的C3转化酶,但其生理意义尚不明了。CR1和H因子也是具有辅因子活性的补体调节蛋白,但H因子的这一活性仅为MCP的1/50。
人MCP的基因定位于第1号染色体长臂32区,基因长度至少为43kb,含有14个外显子和13个内含子。Seya等用从HSB-2T细胞纯化获得的MCP氨基末端设计了一个17mer的反义寡核苷酸探针,以此从U937的cDNA文库中筛选到一条长1.5kb的cDNA。经测序表明,该cDNA中含有一个43bp的5`端非编码区和一个编码384个氨基酸的开读框。其中前34个氨基酸为信号肽,后350个氨基酸为MCP的多肽链,分子量为39kDa。在多肽链的前250个氨基酸中,含有4个相邻的CSR。SCR后为一段富含丝氨酸、苏氨酸和脯氨酸残基的29个氨基酸片段(可能是高度O-连接的 糖基化位点),再后依次为13个氨基酸的功能不明区、24个氨基酸的跨膜区、105个氨基酸的胞浆锚和23个氨基酸构成的胞浆尾部。大多数MCCP的变异局限在富含丝氨酸/苏氨酸(S/T)区和胞浆尾部。在核苷酸序列上MCP与DAF具有同源性。
五、H因子
H因子由Nilson等(1965)发现,根据电泳位将其命名为β1H,而Whaley和Ruddy则将其命名为C3b灭活剂加速因子。现已确定其为由1213个氨基酸组成的单链糖蛋白,分子量155kDa,既有长杆状部分,也有球形区域。新近用透射电镜检查发现,H因子的影象为一长而柔顺的分子。伸长型分子长49.5nn,横截直径为3.4mm但大多数不呈线型而呈折叠状,因此分子的实际长度仅为伸展型的一半,但其构象则呈多样化。通过园二色谱分析表明,H因子既无α螺旋也无β折叠,借二硫键维持其功能活性的构象。H因子的功能包括以下几个方面:(1)为Ⅰ因子的辅助因子,可增加C4b对Ⅰ因子的敏感性。在无H因子时,Ⅰ因子与C3b的结合呈丝状;而在有H因子存在时,I因子与C3b的结合变为弯丝状,同时与C3b的结合亲和力增强(较无H因子时至少高15倍)。H因子强化I因子的机理,可能是H因子与C3b结合后,使C3b出现某些构象变化,增加了与I因子结合的亲和力。H因子与C3b结合的活性部位存在于其N端35kDa部分。(2)加速C3转化酶的衷变:H因子能将已同C3b结合的B因子或Bb从C3酶中逐出,而使之失去酶活性。(3)阻止替代途径中初始和放大C3转化酶的形成。已证实H因子和B因子在C3b上有同一结合部位,故H因子可同B因子或Bb竞争与C3b的结合。在有H因子存在时,B因子不易与C3(H2C)及C3b结合,因此不易形成C3(H2C)Bb或C 3bBb。但H因子对固相上和液相中的C3b作用在差别。对液相中或结合于非激活剂固相上裂解。而对于固定到激活剂(如酵母多糖等)表面的C3b,H因子则对C3b的亲和力与B因子相当,二者竞争的结果,可形成部分C3转化酶,以保证替代途径的活化。有研究报道,在细胞膜上能增强C3b对H因子亲和力的化学成分是涎酸和肝素氨基多糖。由于大多数细菌表面缺乏涎酸,因而这些细菌侵入机体后,可活化替代途径,有助于在早期对感染的控制。(4)对已与P因子或肾炎因子(NeF)结合形成稳定的c 3bBbP或C 3bBbNeF,H因子对它们也有一定的作用,但其效力要比CR1差得多。H因子对C5转化酶(c 3bnBb或C 3bBb3b)的活性也有抑制作用,并能与C5竞争结合C3b使C5不能裂解。此外,近年发现H因子还可诱导单核细胞分泌IL-1参与免疫应答的调节。
编码H因子的基因定位于人的第1号染色体的长臂32区,具有多态性。已发现其有5个变异型,即FH1-5。H因子的核苷酸序列已进行了鉴定,并推导出其全部氨基酸的一级结构,含有20个SCR借此与C3b结合。H因子与MCP、CR1、CR2、DAF及C4bp具有同源性,共同属于补体激活调节剂(RCA)基因家族的成员。
六、I因子
I因子(旧称C3bINA)为异源二聚体血清蛋白,呈双球状结构,分子全长13nm。其中小球(L链)4.9nm,具有丝氨酸蛋白酶活性;大球(H链)5.4nm可与C3b结合。I因子的分子量为88kDa,重链50kDa,轻链38kDa,链间以二硫键相连接。I因子的生物学活性是,在C4bp、MCP、H因子和CR1等辅助因子的协同下,将C4b裂解为C4d和C4c;使C3b裂解出C3f形成C3bi,后者再进一步裂解为C3dg和C3c,由此而控制补体系统的活化。
编码入I因子的基因定位于第4号染色体上。经对I因子的cDNA序列分析发现,其轻链为丝氨蛋白酶的活性区,与糜蛋白酶、胰蛋白酶及弹性蛋白酶等有同源性。而其重链是具有富含半胱氨酸的功能区,与C8、C9和低密度脂蛋白受体等具有同源性。I因子结构基因的突变,可导致先天性I因子缺陷,此类患C3的过度消耗面引起反复感染和血管性水肿。
七、过敏毒素灭活剂
过敏毒素灭活剂(AI)又称血清羧肽酶N,分子量310kDa,由8条相同的多肽链组成,每条分子量各36kDa。AI具有羧基肽酶的活性,可去除C4a、C3a、和C5a C末端的精氨酸残基,使这些片段丧失其过敏毒素活性。
八、S蛋白
S蛋白(CP或S)为血清中一种α单链糖蛋白,分子量83kDa。SP的主要调节作用是可与C5b~7的亚稳态结合部位竞争靶细胞膜脂质,通过形成亲水性的SPC5b~7(简写为S5b~7)复合物,而使C5b~7失去膜结合活性。这样,便可保护补体活化部位邻近的细胞免遭偶然的攻击。这种亲水性的SC5b~7还可集资与1个分子的C8和3个分子的C9结合,分别形成SC5b~8和C5b~(9)3复合物,并C9聚合形成孔道,从而可保护补体活化部位邻近的细胞免于遭受补体的攻击而损伤。SP与C8和C9的结合部位为这两种分子中富含半胱氨酸的功能功能区。电镜下观察,SC5b~(9)3复合物呈一楔形结构,SP位于楔形的宽部可掩盖补体蛋白的疏水区,从而封闭MAC的膜结合部位。此外,2~3个分子的SP与C5b~7与C5b~8复合物的结合,还可使这些复合物易溶,出现亲水向疏水转换。SP也参与凝血过程,通过干扰抗凝血酶Ⅲ对凝血酶的来活而保护凝血酶。
编码人SP基因定位于第17号染色体的长臂上,其cDNA已克隆成功。经过序列分析表明,其与具有细胞粘附作用的玻璃粘连蛋白(vitronectin)的序列完全相同,已证明二者属同一蛋白。
九、CD59
CD59是近年(1989)才发现的一种分子量只有18~20kDa的膜性调节蛋白。由Sugita等(1988年)首先描述,曾有不同的名称,如同种限制因子20(homologous restriction factor-20),保护素(protectin)及膜反应性溶破抑制物(membraneinhibitor of reactive lysis,MIRL)等。CD59由103个氨基酸残基组成,含有单一的N端糖基化位点,其C端借GPI锚固定于细胞表面。CD59分布甚广泛,已证明皮肤、肝、肾、胰、肺、唾腺、神经系统、胎盘以及各种血细胞(红细胞、淋巴细胞、中性粒细胞及血小板)和精子上均有表达。C59的主要生理功能是,防止MAC对同种或自身细胞的溶解破坏,即同种限制作用(homologous species restriction,HSR)。作用机理为:通过其与C7、C8或C9的结合而阻止MAC组装。当其与C5b~7复合物结合后,可阻止其再与C8结合;当基与C8结合后,则可阻碍结合至MAC上的第1个C9分子的铺展,从而是阻止后继C9的结合;而当其与MAC中的C9结合后,又可阻止C9进一步分子的聚合。这样,在有CD59存在的情况下,在同种或自身细胞的表面便不能顺利地完成MAC的全部组装,或不能组装成C5b~(9)n复合物,从而限制了对同种或自身细胞的溶解作用。Rooney等(1991)的报道表明,CD59的这种同种限制作用的机理对保护子宫内的胎儿不被MAC的攻击也有作用。此外,CD59还促进T细胞粘附与激活,维持精子在女性生殖道内的稳定性与活性,以及参与PNH发病机理的作用。
CD59的cDNA已经克隆,并确定CD59的基因定位于人的第11号染色体短臂,其核苷酸序列与氨基酸序列相对应,并与小鼠的CD59有同源性。
十、SP40/40
自1982年起,生物科学技术文献中先后提出一批术语。后经鉴定它们都是蛋白质,且结构类同、功能相近,与补体的末端成分相关。这些成分有不同的名称:如Clusterin、SGP-2、SP40/40、TRRM-2、T64、GP111、SP80、CLI、apo-J和NA1/NA2等。Fritz等建议用Clusterin(暂译为群集素)来概括。
群集素的调节作用是在SP40/40这一同源体中发现的。SP40/40是于70年代末首先在人精浆中发现的,继后在肾小球沉积的IC、血液和MAC中也检出。并由于基常伴随末端补体复合物而存在,故认为其是末端补体复合物的成员之一。SP40/40为血浆中的α糖蛋白,分子量80kDa,由分子量均为40kDa的α和β亚单位所组成,所以称其为SP40/40。现知SP40/40由427个氨基酸残基所组成,α亚单位为222个氨基酸残基,β亚单位为205个氨基酸残基。α和β亚单位借3个二硫键相连接而构成一异二聚体。两个亚单位的氨基酸组成除甘氨酸外,其余均很相似,但氨基酸的序列则相差明显。人血浆中SP40/40的含量为35~105μg/ml(平均62μg/ml),但在人精浆中却可高达15mg/ml,认为这与保护精子的活性有关。
SP40/40是MAC组装的调节蛋白。它可与C5b~7、C5b~8或C5b~9结合,对MAC的组装起抑制作用,从而可防止MAC的溶细胞作用。此外SP40/40与S蛋白还具有协同作用,可使MAC变为可溶性的而失去溶细胞作用。现知C8与SP40/40的结合部位是C8β、C9与SP40/40的结合部位是C9b。
十一、同种限制因子
同种限制因子(homologousrestriction factor,HRF),又称C8结合蛋白(C8bp)。为Zalman等(1986)用C9-Sepharose亲和层析从人红细胞膜上分离的一个能与C8、C9结合并参与C9阶段同种限制性的膜蛋白,通过GPI锚固定于细胞膜表面。新鲜红细胞膜上的HRF分子量为65kDa,而在冷冻的陈旧性红细胞膜上则降解为38kDa。HRF存在于正常人的红细胞、中性粒细胞、单核细胞、淋巴细胞及血小板上。HRF对反应性溶血作用的严格的种属限制性。如经抗HPF抗体处理的人红细胞对人C8、C9的反应性溶血作用敏感性增高,而对8种动物来源的C8、C9的反应性溶血作用则不敏感。体外试验表明,HRF能抑制C9与C8的结合及C9聚合,从而阻止MAC插入自身细胞的膜脂质双层及细胞溶解。HRF可能还与淋巴细胞杀(C9RP)介导的人大颗粒淋巴对羊红细胞和PNH患者红细胞的ADCC作用。
关于上述11种补体调节分子的特性及生物学活性见表5-2
表5-2 补体调节分子的特性及生物学活性
| 调节分子 | 分子量(kDa)(分子特征) | 血清浓度(μg/ml)或细胞分布 | 能特异性相互作用的分子 | 生物学活性 |
| 血清蛋白: | ||||
| C1INH | 104 | 200 | C1r,C1s | 与C1r和C1s共价结合使C1失活; |
| (serpin) | 与C1结合阻止其自发激活:抑制激肽释放酶、纤溶酶及凝血因子Ⅻa、Ⅺa | |||
| C4bp | 550(12个SCR) | 250 | C4b | 抑制CP中的C3转化酶形成;加速CP中的C3、C5转化酶衰变;作为I因子裂解C4b的辅助因子 |
| H因子 | 155(20个SCR) | 480 | C3b | 作为I因子裂解C3b的辅助因子;加速AP中的C3转化酶衰变 |
| I因子 | 88 | 35 | C4b,C3b | 在辅助因子协同下,裂解C4b和C3b |
| AI | 31(羧肽酶N) | 35 | C3a,C4a,C5a | 水解C末端精氨酸残基,灭活过敏毒素 |
| S蛋白 | 83(Vitronectin) | 505 | C 5b~7 | 与C 5b~7结合形成SC5b~7,使之失去膜结合活性 |
| SP40/40 | 80(异二聚体) | 50 | C5b~9 | 调控MAC的组装;保护精子活性 |
| 膜蛋白分子: | ||||
| MCP(CD46) | 40~70(4个SCR,GPI锚) | 除红细胞外的血细胞、上皮细胞、内皮细胞 | C3b,C3b | 作为I因子裂解C3b和C4b的辅助因子 |
| DAF(CD55) | 70(4个SCR,GPI锚) | 绝大多数血细胞、内皮细胞、粘膜上皮细胞 | C 4b2a,C 3bBb | 抑制C3转酶形成 促进CP和AP中的C3转化酶衰变 |
| HRF(C8bp) | 65(GPI锚) | 红细胞、PMN、单核细胞、淋巴细胞、血小板 | C8、C9 | 抑制C9与C8结合及C9聚合;阻止MAC插入自身细胞膜 |
| CD59(MIRL) | 18~20(GPI锚) | 红细胞、PMN、淋巴细胞、血小板 | C7,C8,C9 | 通过与C7、C8或C9结合而阻止MAC的组装,防止MAC溶解同种或自身细胞 |