第一章 绪论
第一节 生理学的研究对象和任务
一、生理学
生理学(physiology)是生物科学中的一个分支,它以生物机体的功能为研究对象。生物机体的功能就是整个生物及其各个部分所表现的各种生命现象或生理作用,例如呼吸、消化、循环、肌肉运动等等。生理学的任务就是要研究这些生理功能的发生机制、条件以及机体的内外环境中各种变化对这结功能的影响,从而掌握各种生理变化的规律。
二、生理学与医学的关系
生理学的发展与医学的密切联系。在医疗实践中和对人体的一般观察中积累了关于人体生理功能的许多知识,更通过对于人体和动物的实验分析研究,进一步深入探索这些生理功能的内在机制和相互关系,逐渐形成关于人和动物机体功能的系统性理论科学。医学中关于疾病问题的理论研究是以人体生理学的基本理论为基础的;同时,通过医学实践又可以检验生理学理论是否正确,并不断以新的内容和新的问题丰富生理学理论和推动生理学研究。因此,生理学是医学的一门基础理论科学。
三、生理学研究的不同水平
在研究生命现象的机制时,需要从各个不同水平提出问题进行研究。根据研究的层次不同,生理学研究可以分成三个水平。
一是关于生命现象的细胞和分子机制的研究。生理活动的物质基础是生物机体,构成机体的最基本结构和功能单位是各种细胞,每一器官的功能都与组成该器官的细胞的生理特性分不开,例如肌肉的功能与肌细胞的生理特性分不开,腺体的功能与腺细胞的生理特性分不开,等等。然而,细胞的生理特性又决定于构成细胞的各个物质的物理化学特性,尤其是生物大分子的物理化学特性。例如心脏之所以能搏动,是由于肌细胞中含有特殊的蛋白质,这些蛋白质分子具有一定的结合排列方式,在离子浓度的变化和酶的作用下排列方式发生变化,从而发生收缩或舒张的活动。因此,对心脏功能的功能的研究需要在肌细胞和生物大分子的水平上进行。这类研究的对象是细胞和它所含的物质分子,可称为细胞和分子水平的研究。这方面的知识称为普遍生理学或细胞生理学。
二是关于机体内各器官和系统的功能的研究。这方面的研究着重于阐明器官和系统对于机体有什么作用,它是怎样进行活动的,它的活动受到哪些因素的控制,等等。例如,关于心血管组成的血液循环系统的生理功能研究,需要阐明心脏各部分如何协同活动、心脏如何射血、血管如何调配血液供给、血管内血液流动的动力和阻力、心血管活动如何调节等规律。这类研究要对完整的心脏、血管和循环系统进行观察,是以器官和系统作为研究对象的,称为器官和系统水平的研究。这方面的知识称为器官和系统生理学。
三是关于机体内各器官、系统的相互联系和相互影响,以及机体与环境之间相互联系和相互影响的研究。由于人体生理学的研究对象是人的机体,整个人体的生理活动并不等于心、肺、肾等器官生理功能的简单总和,而是在各种生理功能之间体现着彼此相互联系、相互制约的完整而协调的过程。人的生理活动还具有个体的特点,并且随着个体生活条件的变异而不断变化发展着。机体内的这种联系制约、变化发展的规律也是需要加以研究的。例如,在完整人体内心脏搏动的频率和力量,会受体内外环境条件、人体的健康情况以及情绪等因素的影响。在这里,研究的对象是整个机体,可称为整体水平的研究。
生理功能虽然以细胞和分子特性为基础,并服从于物理化学的规律,但生理学毕竟不等同于物理学和化学,它们既有细胞和分子水平的研究和科学规律,还有器官、系统和整体水平的研究和科学规律。要全面地理解某一生理功能的机制,必须从细胞和分子、器官和系统、以及整体三个水平进行研究。
第二节 生理学的发展简史
人体生理的知识最初是随着生产和医疗实践而逐渐积累起来的。公元300-400年的《黄帝内经》一书是我国古代医疗实践经验的理论总结,书中阐述了经络、脏腑、七情六淫、营卫气血等生理学理论。在其他国家,早期对人体生理知识也有不少重要的贡献。例如,古罗马名医Galen曾从人体解剖的知识来推论生理功能,并曾进行初步的动物活体解剖,对医学的贡献很大。
生理学真正地成为一门实验性科学是从17世纪开始的。1628年英国医生Harvey证明了血液循环的途径,并指出心脏是循环系统的中心。他的结论是在几种动物身上应用活体解剖法通过多次实验而获得的。在17-18世纪,显微镜的发明和物理学、化学的迅速进步,都给生理学的发展准备了良好的条件。例如,应用显微镜发现了毛细血管,证实了Harvey对循环系统结构的推论;物质守恒与能量守恒及转化定律的提出,以及燃烧和呼吸原理的阐明,为机体新陈代谢的研究奠定了基础。到了19世纪,随着其他自然科学的迅速发展,生理学实验研究也大量开展,累积了大量各器官生理功能的知识。例如,关于感觉器官、神经系统、血液循环、肾的排泄功能、内环境稳定等的研究,均为生理功能提供了不少宝贵资料。我国现代生理学也有60余年的历史。1926年北京协和医学院生理系主任林可胜发起创建了中国生理学会,随即出版了《中国生理学杂志》。学会的成立和专业杂志的出版,对于生理学在我国的发展起了很好的推动作用。当时我国比较集中的研究工作是关于胃液分泌、物质代谢、神经肌肉和心血管运动的神经调节等问题,并在学术上作出了贡献,受到国际生理学界的重视。
近二三十年来,由于基础科学和新技术的迅速发展,以及相关学科间的交叉渗透,使生理学的研究有了很大的进展。细胞、分子水平的研究,已深入到细胞内部环境的稳态及其调节机制、细胞跨膜信息传递的机制、基因水平的功能调控机制等方面,使生命活动基本规律的研究取得了不少宝贵资料。在整体水平研究方面,由于学科的交叉渗透,不断产生了研究的新领域,如神经免疫内分泌学等;而且由于采用了许多先进技术,使整体生理学研究,尤其是各种特殊条件下(如劳动、运动、高空、高原、潜水等)的研究,取得了很大进展。生理学向微观的细胞分子水平深入发展,以及向宏观的整体水平加快扩展,是当今发展的总趋势。
第三节 生理功能的调节
人体和复杂多细胞动物的细胞直接生存于细胞外液中,而不与外环境发生接触。细胞新陈代谢所需的养料由细胞外液提供,细胞的代谢产物也排到细胞外液中,而后通过细胞外液再与外环境发生物质交换。由此,细胞外液被称为机体的内环境,以别于整个机体所生存的外环境。细胞的生存对内环境条件的要求很严格,内环境各项因素的相对稳定性乃是高等动物生命存在的必要条件。然而,内环境理化性质不是绝对静止的,而是各种物质在不断转换中达到相对平衡状态,即动态平衡状态。这种平衡状态称为稳态。由于细胞不断进行着新陈代谢,新陈代谢本身不断扰乱内环境的稳态,外环境的强烈变动也可影响内环境的稳态;为此,机体的血液循环、呼吸、消化、排泄等生理功能必须不断地进行着调节,以纠正内环境的过分变动。
一、神经调节
神经活动的基本过程是反射。反射的结构基础为反射弧,包括五个基本环节:感受器、传入神经、神经中枢、传出神经和效应器。感受器是接受刺激的器官,效应器是产生反应的器官;中枢在脑和脊髓中,传入和传出神经是将中枢与感受器和效应器联系起来的通路(参见第十章第二节)。例如当血液中氧分压下降时,颈动脉等化学感受器发生兴奋,通过传入神经将信息传至呼吸中枢导致中枢兴奋,再通过传出神经使呼吸肌运动加强,吸入更多的氧使血液中氧分压回升,维持内环境的稳态。反射调节是机体重要的调节机制,神经系统功能不健全时,调节将发生混乱。
巴甫洛夫(лавдов)将反射分成非条件反射与条件反射两类。非条件反射是先天遗传的,同类动物都具有的,是一种初级的神经活动。上述呼吸反射就是一种简单的非条件反射。条件反射是后天获得的,是个体在生活过程中按照它的生活条件而建立起来的,是一种高级的神经活动。例如,工人进入劳动环境中就会发生呼吸加强的条件反射,这时虽然劳动尚未开始,但呼吸系统已增强活动,为劳动准备提供足够的氧并排出二氧化碳。所以,条件反射是更具有适应性意义的调节。
二、体液调节
体液调节就是机体某些细胞产生某些特殊的化学物质,借助于血液循环的运输,到达全身各器官组织或某一器官组织,从而引起这器官组织的某些特殊的反应。许多内分泌细胞所分泌的各种激素,就是借体液循环的通路对机体的功能进行调节的。例如,胰岛B细胞分泌的胰岛素能调节组织、细胞的糖与脂肪的新陈代谢,有降低血糖的作用。内环境血糖浓度之所以能保持相对稳定,主要依靠这种体液调节。
有些内分泌细胞可以直接感受内环境中某种理化因素的变化,直接作出相应的反应。例如,当血钙离子浓度降低时,甲状旁腺细胞能直接感受这种变化,促使甲状旁腺激素分泌增加,转而导致骨中的钙释放入血,使血钙离子的浓度回升,保持了内环境的稳态。也有些内分泌腺本身直接或间接地受到神经系统的调节,在这种情况下,体液调节是神经调节的一个传出环节,是反射传出道路的延伸。这种情况可称为神经-体液调节。例如,肾上腺髓质接受感神经的支配,当交感神经系统兴奋时,肾上腺髓质分泌的肾上腺素和去甲肾上腺素增加,共同参与机体的调节。
除激素外,某些组织、细胞产生的一些化学物质,虽不能随血液到身体其他部位起调节作用,但可在局部组织液内扩散,改变邻近组织细胞的活动。这种调节可看作是局部性体液调节,或称为旁分泌(paracrine)调节。
神经调节的一般特点是比较迅速而精确,体液调节的一般特点是比较缓慢、持久而弥散,两者相互配合使生理功能调节更趋于完善。
三、自身调节
自身调节是指组织、细胞在不依赖于外来的或体液调节情况下,自身对刺激发生的适应性反应过程。例如,骨骼肌或心肌的初长(收缩前的长度)能对收缩力量起调节作用;当初长在一定限度内增大时,收缩力量会相应增加,而初长缩短时收缩力量就减小。一般来说,自身调节的幅度较小,也不十分灵敏,但对于生理功能的调节仍有一定意义。
有时候一个器官在不依赖于器官外来的神经或体液调节情况下,器官自身对刺激发生的适应性反应过程也属于自身调节。
第四节 生理功能的调节控制
本世纪40年代,通过运用数学和物理学的原理和方法,分析研究各种工程技术的控制和人体的各种功能调节,得出了一些有关调节和控制过程的共同规律,产生了一个新的学科,这就是控制论(cybernetics)。运用控制论原理分析人体的调节活动时,人体的各种功能调节可分为三类控制系统。
一、非自动控制系统
非自动控制系统是一个开环系统(open-loop system),其控制部分不受受控部分的影响,即受控部分不能反馈改变控制部分的活动。例如在应激反应中,当应激性刺激特别强大时,可能由于下丘脑神经元和垂体对血中糖皮质激素的敏感性减退,亦即糖皮质激素血中浓度升高时不能反馈抑制它们的活动,使应激性刺激能导致ACTH与糖皮质激素的持续分泌;这时,肾上腺皮质能不断地根据应激性刺激的强度作出相应的反应(参见第十一章)。在这种情况下,刺激决定着反应,而反应不能改变控制部分的活动。这种控制系统无自动控制的能力。非自动控制系统的活动在体内不多见。
二、反馈控制系统
反馈控制系统是一个闭环系统(open-loop system),其控制部分不断接受受控部分的影响,即受控部分不断有反馈信息返回输给控制部分,改变着它的活动。这种控制系统具有自动控制的能力。
图1-1是反馈控制系统的模式图。图中把该系统分成比较器、控制系统、受控系统三个环节;输出变量的部分信息经监测装置检测后转变为反馈信息,回输到比较器,由此构成闭合回路。在不同的反馈控制系统中,传递信息的方式是多种多样的,可以是电信号(神经冲动)、化学信号或机械信号,但最重要的是这些信号的数量和强度的变化中所包含的准确的和足够的信息。参考信息即输入信息(Si),它和反馈信息(Sf)比较后,即得出偏差信息(Se)。三者的关系为:Se=Si+Sf如果是负反馈(negative feedback),则Sf为负值;如果是正反馈(positive feedback),则Sf为正值。
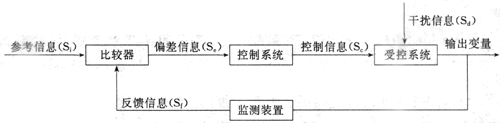
图1-1反馈控制系统模式图
在负反馈情况时,反馈控制系统平时处于稳定状态。如出现一个干扰信息(Sd)作用于受控系统,则输出变量发生改变,导致该反馈控制系统发生扰乱;这时反馈信息与参考信息发生偏差,偏差信息作用于控制系统使控制信息(Sc)发生改变,以对抗干扰信息的干扰作用,使输出变量尽可能恢复到扰乱前的水平。例如,人体的体温经常可稳定在37°C左右,就是负反馈调控作用的结果。现在认为下丘脑内有决定体温水平的调定点的神经元,这些神经元发出参考信息使体温调节中枢发出控制信息来调节产热和散热过程,保持体温维持在37°C左右。如果人体进行剧烈运动,产热突然增加(即发生干扰信息,使输出变量增加)体温随着升高,则下丘脑内的温度敏感(监测装置)就发生反馈信息与参考信息进行比较,由此产生偏差信息作用于体温调节中枢,从而改变控制信息来调整产热和散热过程,使升高的体温回降,恢复到场37°C左右。
在正反馈情况时,反馈控制系统则处于再生状态。正反馈控制系统一般不需要干扰信息就可进入再生状态,但有时也可因出现干扰信息而触发再生。例如,出现一个干扰信息作用于受控系统,则输出变量发生改变,这时反馈信息为正值,导致偏差信息增大;增大的偏差信息作用于控制系统使控制信息增强,导致输出变量的改变进一步加大;由于输出变量加大,又返回来加大反馈信息,如此反复使反馈控制系统活动不断再生。分娩过程是正反馈控制系统活动的实例。当临近分娩时,某些干扰信息可诱发子宫收缩,子宫收缩导致胎儿头部牵张子宫颈部;宫颈受到牵张可反射性导致催产素分泌增加,从而进一步加强宫缩,转而使宫颈进一步受到牵张;如此反复再生,直至胎儿娩出为止。
三、前馈控制系统
图1-2为一个前馈控制系统模式图。从图可以看出,输出变量不发出反馈信息,监测装置在检测到干扰信息后发出前馈(feed forward)信息,作用于控制系统,调整控制信息以对抗干扰信息对受控系统的作用,从而使输出变量保持稳定。因此,前馈控制系统所起的作用是预先监测干扰,防止干扰的扰乱;或是超前洞察动因,及时作出适应性反应。条件反射活动是一种前馈控制系统活动。例如,动物见到食物就引致唾液分泌,这种分泌比食物进入口中后引致唾液分泌来得快,而且富有预见性,更具有适应性意义。但前馈控制引致的反应,有可能失误;例如动物见到食物后并没有吃到食物,则唾液分泌就是一种失误。在进食过程中,导致迷走神经兴奋,促使胰岛B细胞分泌胰岛素来调节血糖水平,这样可及早准备以防止食物消化吸收后造成血糖水平出现过分波动,这也是前馈控制的例子。
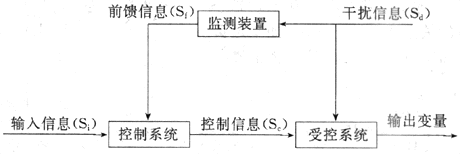
图1-2 前馈控制系统模式图
参考资料
徐丰彦,张镜如主编。人体生理学。北京:人民卫生出版社,1989
王志均,陈孟勤主编。中国生理学史。北京:北京医科大学、中国协和医科大学联合出版社,1993
黄秉宪,潘华。控制理论在生物医学中的应用。生理科学进展1979;10:54~62
Koushapour E.Renal Physiology,Principles and Functions,pp 525~552,WB SnundersCo,Philadelphia,1976
Houks JG.Homeostasis and ControlPrinciples,in Medical Physiology,VB Mountcastle ed,14th ed Vol 1,pp 246~269,CV Mosby Co,St Louis,1980
第二章 细胞的基本功能
细胞是人体和其他生物体的基本结构单位。体内所有的生理功能和生化反应,都是在细胞及其产物(如细胞间隙中的胶原蛋白和蛋白聚糖)的物质基础上进行的。一百多年前,光学显微镜的发明促成了细胞的发现。此后对细胞结构和功能的研究,经历了细胞水平、亚细胞水平和分子水平等具有时代特征的研究层次,从细胞这个小小的单位里揭示出众多生命现象的机制,积累了极其丰富的科学资料。可以认为,离开了对细胞及构成细胞的各种细胞器的分子组成和功能的认识,要阐明物种进化、生物遗传、个体的新陈代谢和各种生命活动以及生长、发育、衰老等生物学现象。要阐明整个人体和各系统、器官的功能活动的机制,将是不可能的。事实上,细胞生理学和分子生物学的实验技术和理论,已经迅速地向基础医学和临床医学各部门渗透。因此,学习生理学应由细胞生理开始。
细胞生理学的主要内容包括:细胞膜和组成其他细胞器的膜性结构的基本化学组成和分子结构;不同物质分子或离子的跨膜转运功能;作为细胞接受外界影响或细胞间相互影响基础的跨膜信号转换功能;以不同带电离子跨膜运动为基础的细胞生物电和有关现象;以及肌细胞如何在细胞膜电变化的触发下出现机械性收缩活动。
第一节 细胞膜的基本结构和物质转运功能
一切动物细胞都被一层薄膜所包被,称为细胞膜或质膜(plasmamembrane),它把细胞内容物细胞周围环境(主要是细胞外液)分隔开来,使细胞能相对地独立于环境而存在。很明显,细胞要维持正常的生命活动,不仅细胞的内容物不能流失,而且其化学组成必须保持相对稳定,这就需要在细胞和它所和的环境之间有起屏障作用的结构;但细胞在不断进行新陈代谢的过程中,又需要经常由外界得到氧气和营养物质。排出细胞的代谢产物,而这些物质的进入和排出,都必须经过细胞膜,这就涉及到物质的跨膜转运过程。因此,细胞膜必然是一个具有特殊结构和功能的半透性膜,它允许某些物质或离子有选择的通过,但又能严格地限制其他一些物质的进出,保持了细胞内物质成分的稳定。细胞内部也存在着类似细胞膜的膜性结构。组成各种细胞器如线粒体、内质网等的膜性部分,使它们与一般胞浆之间既存在某种屏障,也进行着某些物质转运。
膜除了有物质转运功能外,还有跨膜信息传递和能量转换功能,这些功能的机制是由膜的分子组成和结构决定的。膜成分中的脂质分子层主要起了屏障作用,而膜中的特殊蛋白质则与物质、能量和信息的跨膜转运和转换有关。
一、膜的化学组成和分子结构
从低等生物草履虫以至高等哺乳动物的各种细胞,都具有类似的细胞膜结构。在电镜下可分为三层,即在膜的靠内外两侧各有一条厚约2.5nm的电子致密带,中间夹有一条厚2.5nm的透明带,总厚度约7.0~7.5nm左右这种结构不仅见于各种细胞的细胞膜,亦见于各种细胞器的膜性结构,如线粒体膜、内质网膜、溶酶体膜等,因而它被认为是一种细胞中普遍存在的基本结构形式。
各种膜性结构主要由脂质、蛋白质和糖类等物质组成;尽管不同来源的膜中各种物质的比例和组成有所不同,但一般是以蛋白质和脂质为主,糖类只占极少量。如以重量计算,膜中蛋白质约为脂质的1~4倍不等,但蛋白质的分子量比脂质大得多,故膜中脂质的分子数反较蛋白质分子数多得多,至少也超过蛋白质分子数100倍以上。
各种物质分子在膜中的排列形式和存在,是决定膜的基本生物学特性的关键因素。分子生物学的研究成果表明,各种物质特别是生物大分子在各种生物结构中的特殊有序排列,是各种生命现象得以实现的基础。尽管目前还没有一种能够直接观察膜的分子结构的较方便的技术和方法,但根据对生物膜以及一些人工模拟膜特性的分析研究,从30年代以来就提出了各种有关膜的分子结构的假说,其中得到较多实验事实支持而目前仍为大多数人所接受的则70年代初期(Singer和Nicholson,1972)提出的液态镶嵌模型(fluid mosaic model)。这一假想模型的基本内容是:膜的共同结构特点是以液态的脂质双分子层为基架,其中镶嵌着具有不同分子结构、因而也具有不同生理功能的蛋白质,后者主要以а-螺旋或球形蛋白质的形式存在(图2-1)
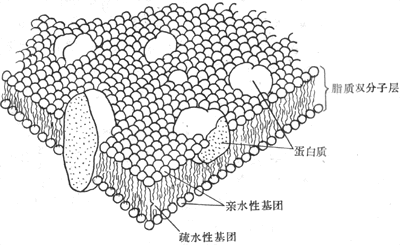
图2-1 膜的液态镶嵌式模型
膜外侧蛋白质和脂质分子上可能存在的糖链未画出
(一)脂质双分子层
膜的脂质中以磷脂类为主,约占脂质总量的70%以上;其次是胆固醇,一般低于30%;还有少量属鞘脂类的脂质。磷脂的基本结构是:一分子甘油的两个羟基同两分子脂酸相结合,另一个羟基则与一分子磷酸结合,后者再同一个碱基结合。根据这个碱基的不同,动物细胞膜中的磷脂主要有四种(图2-2):磷脂酰胆碱、磷脂酰乙醇胺、磷脂酰丝氨酸和磷脂酰肌醇。鞘脂类的基本结构和磷脂类似,但不含甘油。胆固醇结构很特殊,它含有一个甾体结构(环戊烷多氢菲)和一个8碳支链。
最初提示膜中脂质呈双分子层形式存在的,是对红细胞膜所作的化学测定和计算。Gortert和Grendel(1925)提取出红细胞膜中所含的脂质,并测定将这些脂质以单分子层在水溶液表面平铺时所占的面积,结果发现一个红细胞膜中脂质所占的面积,差不多是该细胞表面积的2倍。因此导致以下结论:脂质可能是以双分子层的形式包被在细胞表面的。以后提出的双分子层模型中,每个磷脂分子中由磷酸和碱基构成的基团,都朝向膜的外表面或内表面,而磷脂分子中两条较长的脂酸烃链则在膜的内部两两相对(图2-1)。脂质分子的这种定向而整齐的排列,是由脂质分子本身的理化特性和热力学定律所决定。所有的膜脂质都是一些双嗜性分子,磷脂的一端的磷酸和碱基是亲水性极性基团,另一端的长烃链则属疏水性非极性基团。当脂质分子位于水表面时,由于水分子是极性分子,脂质的亲水性基团将和表面水分子相吸引,疏水性基团则受到排斥,于是脂质会在水表面形成一层亲水性基团朝向水面而疏水性基团朝向空气的整齐排列的单分子层。从热力学业角度分析,这样组成的系统包含的自由能最低,因而最为稳定,可以自动形成和维持。根据同样的原理,如果让脂质分子在水溶液中受到激烈扰动时,脂质有可能形成含水的小囊,但这囊只能是由脂质双分子层形成,外层脂质的极性基团和囊外水分子相吸引,内层脂质的极性基团则和囊内水分子相吸引,而两层脂质的疏水性烃链将两两相对,排斥水分子在囊膜中的存在,其结构正和天然生物膜一致。这种人工形成的人工膜囊,称为脂质小体(liposome),似人造细胞空壳,有很大的理论研究和实用价值。由此可见,脂质分子在细胞膜中以双分子层的形式存在,是由脂质分子本身的理化特性所决定的。设想进化过程中最初有生物学功能的膜在原始的海洋中出现时(也可能包括新的膜性结构在细胞内部的水溶液中的生成),这些基本的理化原理也在起作用。
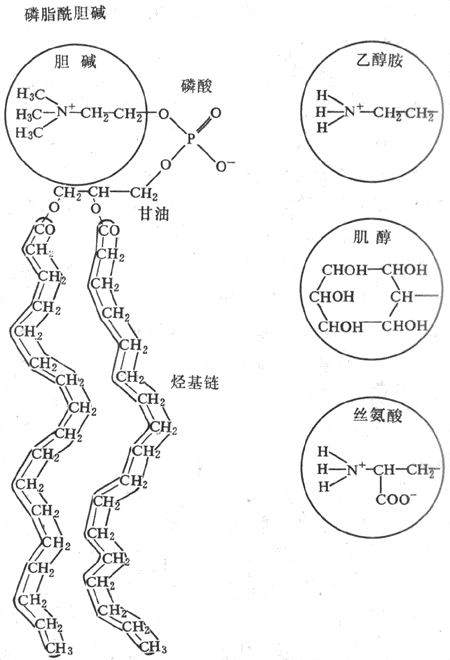
图2-2 磷脂的分子组成
脂质的熔点较低,这决定了膜中脂质分子在一般体温条件下是呈液态的,即膜具有某种程度的流动性。脂质双分子层在热力学上的稳定性和它的流动性,能够说明何以细胞可以承受相当大的张力和外形改变而不致破裂,而且即使膜结构有时发生一些较小的断裂,也可以自动融合而修复,仍保持连续的双分子层的形式。观察一下体内某些吞噬细胞通过毛细血管壁内皮细胞间隙时的变形运动和红细胞通过纤细的毛细血管管腔时被扭曲而不破裂的情况,当会对细胞膜的可变性和稳定性有深刻的印象。当然,膜的这些特性还同膜中蛋白质和膜内侧某些特殊结构(称为细胞架)的作用有关。应该指出的是,膜的流动性一般只允许脂质分子在同一分子层内作横向运动;由于分子的双嗜性,要脂质分子在同一分子层内作“掉头”运动;或由一侧脂质层移到另一侧脂质层,这意味着有极性的磷酸和碱基的一端要穿越膜内部的疏水性部分,这是不容易或要耗能的。
不同细胞或同一细胞而所在部位不同的膜结构中,脂质的成分和含量各有不同;双分子层的内外两层所含的脂质也不尽相同,例如,靠外侧的一层主要含磷脂酰胆碱和含胆碱的鞘脂,而靠胞浆侧的一层则有较多的磷脂酰乙醇胺和磷脂酰丝氨酸。胆固醇含量在两层脂质中无大差别;但它们含量的多少和膜的流动性大小有一定关系,一般是胆固醇含量愈多,流动性愈小。近年来发现,膜结构中含量相当少的磷脂酰肌醇,几乎全部分布在膜的靠胞浆侧;这种脂质与细胞接受外界影响,并把信息传递到细胞内的过程有关。
(二)细胞膜蛋白质
膜结构中含有蛋白质早已证实,但有兴趣的问题是膜中蛋白质究以何种形式存在。70年代以前,多数人主张蛋白质是平铺在脂质双分子层的内外两侧,后来证明,蛋白质分子是以а-螺旋或球形结构分散镶嵌在膜的脂质双分子层中。
膜蛋白质主要以两种形式同膜脂质相结合:有些蛋白质以其肽链中带电的氨基酸或基团,与两侧的脂质极性基团相互吸引,使蛋白质分子像是附着在膜的表面。这称为表面蛋白质;有些蛋白质分子的肽链则可以一次或反复多次贯穿整个脂质双分子层,两端露出在膜的两侧,这称为结合蛋白质。在用分子生物学技术确定了一个蛋白质分子或其中亚单位的一级结构、即肽链中不同氨基酸的排列顺序后,发现所有结合蛋白质的肽链中都有一个或数个主要由20-30个疏水性氨基酸组成的片段。这些氨基酸又由于所含基团之间的吸引而形成а-螺旋,即这段肽链沿一条轴线盘旋,形成每一圈约含3.6个氨基酸残基的螺旋,螺旋的长度大致相当于膜的厚度,因而推测这些疏水的а螺旋可能就是肽链贯穿膜的部分,它的疏水性正好同膜内疏水性烃基相吸引。这样,肽链中有几个疏水性а-螺旋,就可能几次贯穿膜结构;相邻的а-螺旋则以位于膜外侧和内侧的不同长度的直肽链连接(参看图2-7和8)。
膜结构中的蛋白质,具有不同的分子结构和功能。生物膜所具有的各种功能,在很大程度上决定于膜所含的蛋白质;细胞和周围环境之间的物质、能量和信息交换,大都与细胞膜上的蛋白质分子有关。
由于脂质分子层是液态的,镶嵌在脂质层中的蛋白质是可移动的,即蛋白质分子可以在膜脂分子间横向漂浮移位;不同细胞膜中的不同蛋白质分子的移动和所在位置,存在着精细的调控机制。例如,骨骼肌细胞膜中与神经肌肉间信息传递有关的通道蛋白质分子,通常都集中在肌细胞膜与神经未梢分布相对应的那些部分;而在肾小管和消化管上皮细胞,与管腔相对的膜和其余部分的膜中所含的蛋白质种类大不相同,说明各种功能蛋白质分子并不都能在所在的细胞膜中自由移动和随机分布,而实际存在着的有区域特性的分布,显然同蛋白质完成其特殊功能有关。膜内侧的细胞骨架可能对某种蛋白质分子局限在膜的某一特殊部分起着重要作用。
(三)细胞膜糖类
细胞膜所含糖类甚少,主要是一些寡糖和多糖链,它们都以共价键的形式和膜脂质或蛋白质结合,形成糖脂和糖蛋白;这些糖链绝大多数是裸露在膜的外面一侧的。这些糖链的意义之一在于以其单糖排列顺序上的特异性,可以作为它们所结合的蛋白质的特异性的“标志”。例如,有些糖链可以作为抗原决定簇,表示某种免疫信息;有些是作为膜受体的“可识别性”部分,能特异地与某种递质、激素或其他化学信号分子相结合。如人的红细胞ABO血型系统中,红细胞的不同抗原特性就是由结合在膜脂质的鞘氨醇分子上的寡糖链所决定的,A型抗原和B型抗原的差别仅在于此糖链中一个糖基的不同。由此可见,生物体内不仅是多聚糖核苷酸中的碱基排列和肽链中氨基酸的排列可以起“分子语言”的作用,而且有些糖类物质中所含糖基序列的不同也可起类似的作用。
二、细胞膜的跨膜物质转运功能
既然膜主要是由脂质双分子层构成的,那么理论上只有脂溶性的物质才有可能通过它。但事实上,一个进行着新陈代谢的细胞,不断有各种各样的物质(从离子和小分子物质到蛋白质等大分子,以及团块性固形物或液滴)进出细胞,包括各种供能物质、合成细胞新物质的原料、中间代谢产物和终产物、维生素、氧和二氧化碳,以及Na+、K+、Ca2+离子等。它们理化性质各异,且多数不溶于脂质或其水溶性大于其脂溶性。这些物质中除极少数能够直接通过脂质层进出细胞外,大多数物质分子或离子的跨膜转运,都与镶嵌在膜上的各种特殊的蛋白质分子有关;至于一些团块性固态或液态物质的进出细胞(如细胞对异物的吞噬或分泌物的排出),则与膜的更复杂的生物学过程有关。
现将几种常见的跨膜物质转运形式分述如下:
(一)单纯扩散
溶液中的一切分子都处于不断的热运动中。这种分子运动的平均动能,与溶液的绝对温度成正比。在温度恒定的情况下,分子因运动而离开某一小区的量,与此物质在该区域中的浓度(以mol/L计算)成正比。因此,如设想两种不同浓度的同种物质的溶液相邻地放在一起,则高浓度区域中的溶质分子将有向低浓度区域的净移动,这种现象称为扩散。物质分子移动量的大小,可用通量表示,它指某种物质在每秒内通过每平方厘米的假想平面的摩尔或毫尔数。在一般条件下,扩散通量与所观察平面两侧的浓度差成正比;如果所涉及的溶液是含有多种溶质的混合溶液,那么每一种物质的移动方向和通量,都只决定于各该物质的浓度差,而与别的物质的浓度或移动方向无关。但要注意的是,在电解质溶液的情况下,离子的移动不仅取决于该离子的浓度也取决于离子所受的电场力。
在生物体系中,细胞外液和细胞内液都是水溶液,溶于其中的各种溶质分子,只要是脂溶性的,就可能按扩散原理作跨膜运动或转运,称为单纯扩散。这是一种单纯的物理过程,区别于体内其他复杂的物质转运机制。但单纯扩散不同于上述物理系统的情况是:在细胞外液和细胞内液之间存在一个主要由脂质分子构成的屏障,因此某一物质跨膜通量的大小,除了取决于它们在膜两侧的浓度外,还要看这些物质脂溶性的大小以及其他因素造成的该物质通过膜的难易程度,这统称为膜对该物质的通透性。
人体体液中存在的脂溶性物质的数量并不很多,因而靠单纯扩散方式进出细胞膜的物质也不很多。比较肯定的是氧和二氧化碳等气体分子,它们能溶于水,也溶于脂质,因而可以靠各自的浓度差通过细胞膜甚或肺泡中的呼吸膜(参见第五章)。体内一些甾体(类固醇)类激素也是脂溶性的,理论上它们也能够靠单纯扩散由细胞外液进入胞浆,但由于分子量较大,近来认为也需要膜上某种特殊蛋白质的“协作”,才能使它们的转运过程加快。
(二)易化扩散
有很多物质虽然不溶于脂质,或溶解度甚上,但它们也能由膜的高浓度一侧向低浓度一侧较容易地移动。这种有悖于单纯扩散基本原则的物质转运,是在膜结构中一些特殊蛋白质分子的“协助”下完成的,因而被称为易化扩散(facilitateddiffusion)。例如,糖不溶于脂质,但细胞外液中的葡萄糖可以不断地进入一般细胞,适应代谢的需要;Na+、K+、Ca+等离子,虽然由于带有电荷而不能通过脂质双分子层的内部疏水区,但在某些情况下可以顺着它们各自的浓度差快速地进入或移出细胞。这些都是易化扩散的例子。易化扩散的特点是:物质分子或离子移动的动力仍同单纯扩散时一样,来自物质自身的热运动,所以易化扩散时物质的净移动只能是由它们的高浓度区移向低浓度区,但特点是它们不是通过膜的脂质分子间的间隙通过膜屏障,而是依靠膜上一些具有特殊结构的蛋白质分子的功能活动,完成它们的跨膜转运。由于蛋白质分子结构上的易变性(包括其构型和构象的改变)和随之出现的蛋白质功能的改变,因而使易化扩散得以进行,并使它处于细胞各种环境因素改变的调控之下。
由载体介导的易化扩散这种易化扩散的特点是膜结构中具有可称为载体(carrier)的蛋白质分子,它们有一个或数个能与某种被转物相结合的位点或结构域(指蛋白质肽链中的某一段功能性氨基酸残基序列),后者先同膜一侧的某种物质分子选择性地结合,并因此而引起载体蛋白质的变构作用,使被结合的底物移向膜的另一侧,如果该侧底物的浓度较低,底物就和载体分离,完成了转运,而载体也恢复了原有的构型,进行新一轮的转运,其终止点是最后使膜两侧底物浓度变得相等。上面提到的葡萄糖进入一般细胞,以及其他营养性物质如氨基酸和中间代谢产物的进出细胞,就属于这种类型的易化扩散。以葡萄糖为例,由于血糖和细胞外液中的糖浓度经常保持在相对恒定的水平,而细胞内部的代谢活动不断消耗葡萄糖而使其胞浆浓度低于细胞外液,于是依靠膜上葡萄糖载体蛋白的活动,使葡萄糖不断进入细胞,且其进入通量可同细胞消耗葡萄糖的速度相一致不同物质通过易化扩散进出细胞膜,都需要膜具有特殊的载体蛋白。
以载体为中介的易化扩散都具有如下的共同特性:(1)载体蛋白质有较高的结构特异性,以葡萄糖为例,在同样浓度差的情况下,右旋葡萄糖的跨膜通量大大超过左旋葡萄糖(人体内可利用的糖类都是右旋的);木糖则几乎不能被载运。(2)饱和现象,即这种易化扩散的扩散通量一般与膜两侧被转运物质的浓度差成正比,但这只是当膜两侧浓度差较小时是如此;如果膜一侧的浓度增加超过一定限度时,再增加底物浓度并不能使转运通量增加。饱和现象的合理解释是:膜结构中与该物质易化扩散有关的载体蛋白质分子的数目或每一载体分子上能与该物质结合的位点的数目是固定的,这就构成了对该物质的量并不能使载运量增加,于是出现了饱和。(3)竞争性抑制,即如果某一载体对结构类似的A、B两种物质都有转运能力,那么在环境中加入B物质将会减弱它对A物质的转运能力,这是因为有一定数量的载体或其结合位点竞争性地被B所占据的结果。目前已经有多种载体从不同动物的各类细胞膜提纯或克隆(clone)。与葡萄糖易化扩散有关的蛋白质的一级结构由一条含近500个氨基酸的肽链组成,而且此肽链有12个疏水性跨膜а-螺旋(二级结构),多次贯穿膜内外,并互相吸引靠拢,形成球形蛋白质分子(三级结构),但其转运葡萄糖时的具体变构过程尚不完全清楚。
2.由通道介导的易化扩散 它们常与一些带电的离子如Na+、K+Ca+、 CI+等由膜的高浓度一侧向膜的低浓度一侧的快速移动有关。对于不同的离子的转运,膜上都有结构特异的通道蛋白质参与,可分为别称为Na+通道、K+通道、Ca+通道等;甚至对于同一种离子,在不同细胞或同一细胞可存在结构和功能上不同的通道蛋白质,如体内至少已发现有三种以上的Ca+通道和7种以上的K+通道等,这种情况与细胞在功能活动和调控方面的复杂化和精密化相一致。通道蛋白质有别于载体的重要特点之一,是它们的结构和功能状态可以因细胞内外各种理化因素的影响而迅速改变:当它们处于开放状态时,有关的离子可以快速地由膜的高浓度一侧移向低浓度一侧;其离子移动的速度是如此之大,因而在关于通道蛋白的分子结构还知之甚少时,就推测是在这种蛋白质的内部出现了一条贯通膜内外的水相孔道使离子能够顺着浓度差(可能还存在着电场力的作用)通过这一孔道,因而其速度远非载体蛋白质的运作速度所能比拟。这是称为通道(channel)的原因。通道对离子的选择性,决定于通道开放时它的水相孔道的几何大小和孔道壁的带电情况,因而对离子的选择性没有载体蛋白那样严格。大多数通道的开放时间都十分短促,一般以数个或数十个ms计算,然后进入失活或关闭状态。于是又推测在通道蛋白质结构中可能存在着类似闸门(gate)一类的基团,由它决定通道的功能状态。许多的离子通道蛋白质已经用分子生物学的技术被克隆,对其结构的研究已证实了上述推测。
通道的开放造成了带电离子的跨膜移动,这固然是一种物质转运形式;但通道的开放是有条件的、短暂的,百离子本身并不像葡萄糖等是一些代谢物,从生理意义上看,载体和通道活动的功能不尽相同。当通道的开放引起带电离子跨膜移动时(如Na+、Ca2+进入膜内或K+移出膜外),移动本身形成跨膜电流(即离子电流);而移位的带电离子在不导电的脂质双分子层(具有电容器的性质)两侧的集聚,将会造成膜两侧电们即跨膜电位的改变,而跨膜电位的改变以及进入膜内的离子、特别是Ca2+,将会引起该通道所在细胞一系列的功能改变。由此可见,通道的开放并不是起转运代谢的作用,而离子的进出细胞,只是把引起通道开放的那些外来信号,转换成为通道所在细胞自身跨膜电位的变化或其他变化,因而是细胞环境因素影响细胞功能活动的一种方式。
(三)主动转运
主动转运指细胞通过本身的某种耗能过程,将某种物质的分子或离子由膜的低浓度一侧移向高浓度一侧的过程。按照热力学定律,溶液中的分子由低浓度区域向高浓度区域移动,就像举起重物或推物体沿斜坡上移,或使电荷逆电场方向移动一样,必须由外部供给能量。在膜的主动转运中,这能量只能由膜或膜所属的细胞来供给,这就是主动的含义。前述的单纯扩散和易化扩散都属于被动转运,其特点是在这样的物质转运过程中,物质分子只能作顺浓度差、即由膜的高浓度一侧向低浓度一侧的净移动,而它所通过的膜并未对该过程提供能量。被动转运时物质移动所需的能量来自高浓度所含的势能(图示2-3左),因而不需要另外供能(2-3右)。被动转运最终可能达到的平衡点是膜两侧该物质的浓度差为零的情况;如果被动转运的是某种离子,则离子移动除受浓度差的影响外,还受当时电场力的影响,亦即当最终的平衡点达到时,膜两侧的电-化学势*的差为应为零。主动转运与此不同,由于膜以某种方式提供了能量,物质分子或离子可以逆浓度或逆电-化学势差而移动。体内某种物质分子或离子由膜的低浓度一侧向高浓度一侧移动,结果是高浓度一侧浓度进一步升高,而另一侧该物质愈来愈少,甚至可以全部被转运到另一侧。如小肠上皮细胞吸收某些已消化的营养物;肾小管上皮细胞对小管液中某些“有用”物质进行重吸收,均属此现象。由于此过程在热力学上为耗能过程,不可能在无供能的情况下自动进行,因此如果在生物体内出现这种情况,说明有主动的跨膜转运在进行,必定伴随了能源物质(常常是ATP)的消耗。
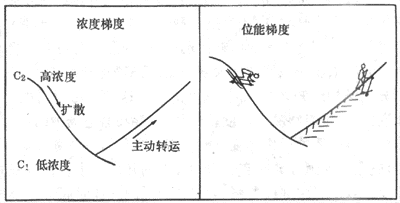
图2-3 物质的主动转运和被动转运原理示意图
物质分子可由高浓度处自动向低浓度处扩散,而分子
由低浓度处移向高浓度处则需另行供能,正如滑雪者可
由高坡自动下滑,而上坡却需要由人体费力一样。
被动转运和主动转运的根本区别即在于此
在细胞膜的主动转运中研究得最充分,而且对细胞的生存和活动可能是最重要的,是膜对于钠和钾离子的主动转运过程。所有活细胞的细胞内液和细胞外液中Na+和K+的浓度有很大的不同。以神经和肌细胞为例,正常时膜内K+浓度约为膜外的30倍,膜外的Na+浓度约为膜内的12倍;这种明显的离子浓度差的形成和维持,要依靠新陈代谢的进行,提示这是一种耗能的过程;例如,低温、缺氧或应用一些代谢抑制剂可引起细胞内外Na+、K+的浓度差减小,而在细胞恢复正常代谢活动后,巨大的浓度差又可恢复。由此认为各种细胞的细胞膜上普遍存在着一种钠-钾泵(sodium-potassium pump)的结构,简称钠泵,其作用是在消耗代谢能的情况下逆烊浓度差将细胞内的Na+移出膜外,同时把细胞外的K+移入膜内,因而保持了膜内高K+和膜外高Na+的不均衡离子分布。
钠泵是镶嵌在膜的脂质双分子层中的一种特殊蛋白质,它除了有对Na+、K+的转运功能外,还具有ATP酶的活性,可以分解ATP使之释放能量,并能利用此能量进行Na+和K+的主动转运;因此,钠泵就是Na+-K+依赖式ATP酶的蛋白质。钠泵蛋白质已用近代分子生物学方法克隆出来,它们是由α-和β-亚单位组成的二聚体蛋白质,肽链多次穿越脂质双分子层,是一种结合蛋白质。α-亚单位的分子量约为100kd,转运Na+、K+和促使ATP分解的功能主要由这一亚单位来完成;β-亚单位的分子量约为50kd,作用还不很清楚。钠泵蛋白质转运Na+、K+的具体机制尚不十分清楚,但它的启动和活动强度与膜内出现较多的Na+和膜外出现较多的K+有关。钠泵活动时,它泵出Na+和泵入K+这两个过程是同时进行或“耦联”在一起的;根据在体内或离体情况下的计算,在一般生理情况下,每分解一个ATP分子,可以使3个Na+移到膜外同时有2个K+移入膜内;但这种化学定比关系在不同情况下可以改变。
细胞膜上的钠泵活动的意义是:(1)由钠泵活动造成的细胞内高K+,是许多代谢反应进行的必需条件;(2)如果细胞允许大量细胞外Na+进入膜内,由于渗透压的关系,必然会导致过多水分了进入膜内,这将引起细胞的肿胀,进而破坏细胞的结构;(3)它能够建立起一种势能贮备。如所周知,能量只能转换而不能消灭,细胞由物质代谢所获得的能量,先以化学能的形式贮存在ATP的高能磷酸键之中;当钠泵蛋白质分解ATP时,此能量用于使离子作逆电-化学势跨膜移动,于是能量又发生转换,以膜两侧出现了具有高电-化学势的离子(分别为K+和Na+)而以势能的形式贮存起来;换句话说,泵出膜外的Na+由于其高浓度而有再进入膜内的趋势,膜内高浓度的K+、则有再有再移了膜的趋势,这就是一种势能贮备。由钠泵造成的离子势能贮备,可用于细胞的其他耗能过程。如下节将详细讨论的Na+、K+等离子在膜两侧的不均衡分布,是神经和肌肉等组织具有兴奋性的基础;由K+、Na+等离子在特定条件下通过各自的离子通道进行的顺电-化学势的被动转运,使这些细胞表现出各种形式的生物电现象。
继发性主动转运钠泵活动形成的势能贮备,还可用来完成一些其他物质的逆浓度差的跨膜转运,这主要见于前面提到的肠上皮和肾小管上皮细胞对葡萄糖、氨基酸等营养物质的较为安全吸收现象,这显然有主动转运过程的参与。但据观察,这种理论上要耗能的过程并不直接伴随ATP或其他供能物质的消耗。这些物质的跨膜转运经常要伴有Na+由上皮细胞的管腔侧同时进入细胞;后者是葡萄糖等进入细胞的必要条件,没有Na+由高浓度的膜外顺浓度差进入膜内,就不会出现葡萄糖等分子逆浓度差进入膜内。在完整的在体肾小管和肠粘膜上皮细胞,由于在细胞的基底-外侧膜(或基侧膜,即靠近毛细血管和相邻上皮细胞侧的膜)上有钠泵存在(图2-4),因而能造成细胞内Na+浓度经常低于小管液和肠腔液中Na+浓度的情况,于是Na+不断由小管液和肠腔液顺浓度差进入细胞,由此释放的势能则用于葡萄糖分子的逆浓度进入细胞。葡萄糖主动转运所需的能量不是直接来自ATP的分解,而是来自膜外Na+的高势能;但造成这种高势能的钠泵活动是需要分解ATP的,因而糖的主动转运所需的能量还是间接地来自ATP,为此把这种类型的转运称为继发性主动转运,或称为联合转运(cotransport)。每一种联合转运也都与膜中存在的特殊蛋白质有关,称为转运体(transporter);而且在不同的情况下,被转运的物质分子有的与Na+移动的方向相同,有时两者方向相反。甲状腺细胞特有的聚碘作用,也属于继发性主动转运。
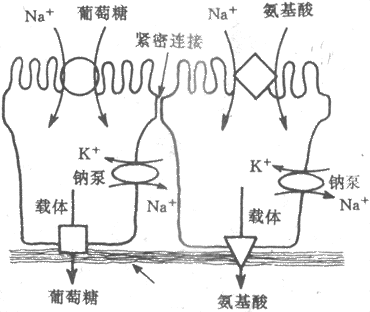
图2-4 葡萄糖和一些氨基酸的继发性主动转运模式图
上方弯曲的管腔侧膜上的圆和方块,分别表示同葡萄糖和某些氨
基酸的继发性转运有关的转运蛋白质
主动转运是人体最重要的物质转运形式,除上述的钠泵外,目前了解较多的还有钙泵(Ca2+-Mg2+依赖式ATP酶)、H+-K+泵(H+-K+依赖式ATP酶)等。这些泵蛋白在分子结构上和钠泵有很大类似,都以直接分解ATP为能量来源,将有关离子进行逆浓度的转运。钙泵主要分布在骨骼肌和心肌细胞内部的肌浆网上,激活时可将胞浆中的Ca+迅速集聚到肌浆网内部,使胞浆中Ca+浓度在短时期内下降达成100倍以上;这是诱发肌肉舒张的关键因素。H+-K+泵主要分布在胃粘膜壁细胞表面,与胃酸的分泌有关。
(四)出胞与入胞式物质转运
细胞对一些大分子物质或固态、液态的物质团块,可通过出胞和入胞进行转运。
出胞主要见于细胞的分泌活动,如内分泌腺把激素分泌到细胞外液中,外分泌腺把酶株颗粒和粘液等分泌到腺管的管腔中,以及神经细胞的轴突末梢把神经递质分泌到突触间隙中。根据在多种细胞进行观察,细胞的各种蛋白性分泌物先是在粗面内质网生物合成;在它们由内质网到高尔基复合体的输送过程中,逐渐被一层膜性结构所包被,形成分泌囊泡;后者再逐渐移向特定部位的质膜内侧,准备分泌或暂时贮存。有些细胞的分泌过程是持续进行的,有些则有明显的间断性。分泌过程或一般的出胞作用的最后阶段是:囊泡逐渐向质膜内侧移动,最后囊泡膜和质膜在某点接触和相互融合,并在融合处出现裂口,将囊泡一次性的排空,而囊泡的膜也就变成了细胞膜的组成部分(图2-5)。这个过程主要是由膜外的特殊化学信号或膜两侧电位改变,引起了局部膜中的Ca2+通道的开放,由内流的Ca2+(内流的Ca2+也有的进而引发细胞内Ca2+贮存库释放Ca2+)触发囊泡的移动、融合和排放。最近在肥大细胞的研究表明,囊泡与质膜的融合,可能与预先“装配”在两侧膜上的类似形成细胞间通道的那种蛋白质分子有关(见下节),当两者“对接”时,囊泡内容与细胞外液相沟通;以后由于组成通道的蛋白质各亚单位分散开来,造成原孔洞的扩大,完成囊泡内容的快速排出,囊泡膜也伸展开来,成为细胞膜的一部分。
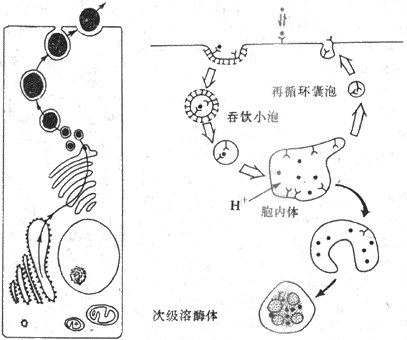
图2-5 分泌物的出胞过程 图 2-6 受体介导式入胞过程示意图
分泌囊泡逐渐向细胞膜内侧面靠近,两者的膜
相互融合,融合处膜膜断裂,分泌物排出,而后囊泡膜成为细胞膜的组成部分
入胞和出胞相反,指细胞外某些物质团块(如侵入体内的细菌、病毒、异物或血浆中脂蛋白颗粒、大分子营养物质等)进入细胞的过程。入胞进行时,首先是细胞环境中的某些物质与细胞膜接触,引起该处的质膜发生内陷,以至包被吞食物,再出现膜结构的断离,最后是异物连同包被它的那一部分膜整个地进入细胞浆中。
一种通过被转运物质与膜表面的特殊受体蛋白质相互作用而引起的入胞现象,称为受体介导式入胞。通过这种方式进入细胞的物质已不下50余种,包括以胆固醇为主要成分的血浆低密度脂蛋白颗粒、结合了铁离子的运铁蛋白、结合了维生素B12的运输蛋白、多种生长调节因子和胰岛素等一部分多肽类激素、抗体和某些细菌毒素,以及一些病毒(流感和小儿麻痹病毒)等(图示2-6)。首先是细胞环境中的某物质为细胞膜上的相应受体所“辨认”,发生特异性结合;结合后形成的复合物通过它们在膜结构中的横向移动,逐渐向膜表面一些称为衣被凹陷(coated pit)的特殊部位集中。衣被陷处的膜与一般膜结构无明显差异,只是向细胞内部呈轻度下凹,而且在膜的胞浆侧有一层高电子密度的覆盖物,后者经分析是由多种蛋白质组成的有序结构;当受体复合物的聚集使衣被凹陷成为直径约0.3μm的斑片时(可以在约1分钟的时间内完成),该处出现膜向胞浆侧的进一步凹入,最后与细胞膜断离,在胞浆内形成一个分离的吞食泡,这称为内移(internalization);原来附在衣被凹陷内侧的蛋白性结构,现在正好位于吞食泡膜的外侧,仍面向胞浆;但在吞食泡形成后不久,这种蛋白结构就消失,可能是溶解在胞浆中,大概还可以再用于在细胞膜上形成新的衣被凹陷。这类蛋白质的功能,据认为是为吞食泡的形成提供所需的能量。失去了这种特殊的附膜蛋白结构的吞食泡,进而再与胞浆中称为胞内体(endosome)的球状或管状膜性结构相融合,此胞内体的特点是内部具有较低的PH值环境,有助于受体同与它结合的物质分离;以后的过程是这些物质(如进入细胞的低密度脂蛋白颗粒和铁离子等)再被转运到能利用它们的细胞器,而保留在胞内体膜上的受体,则与一部分膜结构形成较小的循环小泡,移回到细胞膜并与之融合,再成为细胞的组成部分,使受体和膜结构可以重复使用(图2-6)。据测算,在人工培养液中的吞噬细胞1小时内通过形成吞食泡而进入胞浆的细胞膜面积,大约相当于原细胞膜总面积的50%-200%,而实际细胞膜的总面积并未明显改变,可见通过上述以胞内体为转站的膜的再循环,不仅维持了细胞膜的总面积的相对恒定,而且使相应的受体可以反复使用。
第二节 细胞的跨膜信号传递功能
不论是单细胞生物或组成多细胞有机体的每一个细胞,在它们的生命过程中,都会不断受到来自外部环境的各种理化因素的影响。在多细胞动物,由于绝大多数细胞是生活在直接浸浴它们的细胞外液、即内环境之中,因此出现在内环境中的各种化学分子,是它们最常能感受到的外来刺激:这不仅是指存在于细胞外液中的激素或其他体液性调节因子;而且就是在神经调节过程中,当神经信息由一个神经元向其他神经元传递或由神经元传给它的效应器细胞时,在绝大多数情况下,也都要通过一种或多种神经递质和调质为中介,通过这些化学分子在距离极小的突触间隙液中的扩散,才能作用到下一级神经元或效应器细胞。尽管激素和递质(或调质)等分子作为化学信号在细胞外液中播散的距离和范围有所不同,但对接受它们影响的靶细胞并不存在本质的差别。
细胞外液中的各种化学分子,并不需要自身进入它们的靶细胞后才能起作用(一些脂溶性的小分子类固醇激素和甲状腺激素例外,详见第十一章)它们大多数是选择性地同靶细胞膜上具有特异的受体性结构相结合,再通过跨膜信号传递(transembrane signaling)或跨膜信号转换(transmembranesognal transduction)过程,最后才间接地引起靶细胞膜的电变化或其他细胞内功能的改变。
机体和细胞也可能受到化学信号以外的其他性质的刺激,如机械的、电的和一定波长电磁波等来自外界环境的刺激的影响;但在动物进化的过程中,这些刺激信号大都由一些在结构和功能上高度分化了的特殊的感受器细胞来感受,引起相应的感受器细胞出现某种电反应。仔细分析各种感受器细胞接受它们所能感受的某种特异刺激信号的过程时发现(如耳蜗毛细胞接受声波振动和视网膜光感受细胞接受光刺激等),它们也涉及到外来刺激信号的跨膜传递,即刺激信号也要先作用于膜结构中的感受性结构,才能引起感受器细胞的电变化和随后的传入神经冲动。
不论是化学信号中的激素分子和递质(包括数十种可能起调质作用神经肽类物质)分子,以及非化学性的外界刺激信号,当它们作用于相应的靶细胞时,都是通过为数不多、作用形式也较为类似的途径来完成跨膜信号传递的;这些过程所涉及的膜蛋白质也为数不多,在生物合成上由几类特定基因家族所编码;正因为如此,由每个特定基因家族所表达生成的蛋白质分子,在肽链的氨基酸排列顺序上有较大的相同性(或同源性,homogeneity),功能上也较为类似。因此,关于跨膜信号传递的研究,早已超出了递质或激素作用机制的范畴,成为细胞生理学中一个有普遍意义的新篇章。试想,人体细胞都具有相同的遗传基因,因而一个感光细胞或一个普通体细胞,通过细胞膜上类似的蛋白质,以类似的方式接受它们所受到的外来刺激,可引起细胞本身功能的改变;而且各种不同的细胞通过少数几类膜蛋白质和几种作用方式,就能接受多种多样可能遇到的外界刺激信号的影响,显然符合“生物经济”的原则。
一、由具有特异感受结构的通道蛋白质完成的跨膜信号传递
(一)化学门控通道
对这种跨膜信号的传递方式的研究,最早是从对运动神经纤维末梢释放的乙酰胆碱(Ach)如何引起它所支配的骨骼肌细胞兴奋的研究开始的。早已知道,当神经冲动到达神经末梢处时,先是由末梢释放一定数量的Ach分子,后者再同肌细胞膜上称为终板(指有细胞膜上同神经末相对的那部分膜,其中所含膜蛋白与一般肌细胞膜不同)处的“受体”相结合,引起终板膜产生电变化,最后引起整个肌细胞的兴奋和收缩。由于神经-肌接头处的“受体”也可同烟碱相结合,因而过去在药理学分类中称它为N-型Ach受体。80年代后期,我国学者李镇源发现α-银环蛇毒同N-型受体有极高的特异性结合能力又有人发现一些电鱼的电器官中有密集的这种受体蛋白质分子存在;再依靠70年代以来蛋白质化学和分子生物学技术的迅速发展,目前不仅已将这种蛋白质分子提纯,而且基本上搞清了它的分子结构和它们在膜中的存在形式。原来它是由4种不同的亚单位组成的5聚体蛋白质(图2-7),总分子量约为290kd;每种亚单位都由一种mRNA编码,所生成的亚单位在膜结构中通过氢键等非共价键式的相互吸引,形成一个结构为α2βγδ的梅花状通道样结构(图2-7,A和B),而其中的两个α-亚单位正是同两分子ACH相结合的部位,这种结合可引起通道结构的开放,其几何大小足以使终板膜外高浓度的Na+内流,同时也能使膜内高浓度的K+外流结果是使原来存在两侧的静息电位近于消失,亦即使该处膜内外电位差接近于0值,这就是终板电位,于是完成了Ach这种化学信号的跨膜传递,因为肌细胞后来出现的兴奋和收缩都是以终板电位为起因的。
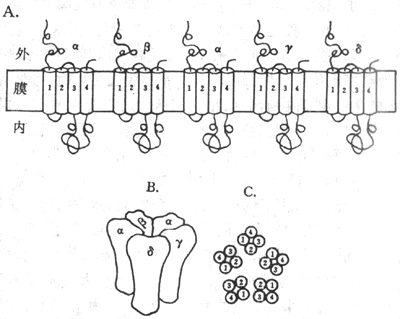
图2-7 N-型Ach门控通道的分子结构示意图
A:N-型Ach门控通的5个亚单位和它们所含α-螺旋在膜中存在形式的平面示意图
B:5个亚单位相互吸引,包绕成一个通道样结构 C:在跨膜通道结构中,各
个亚单位亿含α-螺旋在通道结构中的位置
用分子生物学实验技术证明,同其他膜结合蛋白质类似,在上述4种不同的亚单位肽链中,都存在有4种主要由20-25个疏水性氨基酸形成的α-螺旋,因而推测每个亚单位的肽链都要反复贯穿膜4次(图2-7,A),而5个亚单位又各以其第2个疏水性跨膜α-螺旋构成了水相孔道的“内壁”(图2-7C)。
由上述分子水平的研究成果可以知道,原初将终板膜上完成Ach跨膜信号传递的蛋白质称作“受体”是不符合实际情况的;它们是一种通道样结构,只是在组成通道的蛋白质亚单位中有两个亚单位具有同Ach分子特异地相结合的能力,并能因此引起通道蛋白质的变构作用而使通道开放,然后靠相应离子的易化扩散而完成跨膜信号传递。因此,这种蛋白质应称为N-型(或烟碱型)Ach门控通道,属于化学门控通道或化学依从性通道中的一种。
Ach在神经-肌接头处的跨膜信号传递机制的阐明,曾一度错误地推测,其他一些神经递质也都是以类似的方式作用于下一级神经元或相应的效应器细胞的;但后来的研究表明并非如此。目前只证明了一些氨基酸递质,包括谷氨酸、门冬氨酸、γ-氨基丁酸和甘氨酸等,主要是通过同N-型Ach门控通道结构类似的化学门控通道影响其靶细胞。
(二)电压门控通道
应用类似的技术,在80年代还陆续克隆出几种重要离子(如Na+、K+和Ca2+等离子)的电压门控通道,它们具有同化学门控能道类似的分子结构,但控制这类通道开放与否的因素,是这些通道所在膜两侧的跨膜电位的改变;也就是说,在这种通道的分子结构中,存在一些对跨膜电位的改变敏感的基团或亚单位,由后者诱发整个信道分子功能状态的改变。
在动物界,除了一些特殊的鱼类,一般没有专门感受外界电刺激或电场改变的器官或感受细胞,但在体内有很多细胞,如神经细胞和各种肌细胞,在它们的细胞膜中却具有多种电压门控通道蛋白质,它们可由于同一细胞相邻的膜两侧出现的电位改变而再现通道的开放,并由于随之出现的跨膜离子流而出现这些通道所在膜的特有的跨膜电位改变。例如,前述的终板膜由Ach门控通道开放而出现终板电位时,这个电位改变可使相邻的肌细胞膜中存在的电压门控式Na+通道和K+通道相继激活(即通道开放),出现肌细胞的所谓动作电位;当动作电位在神经纤维膜和肌细胞膜上传导时,也是由于一些电压门控通道被邻近已兴奋的膜的电变化所激活,结果使这些通道所在的膜也相继出现特有的电变化。由此可见,电压门控通道所起的功能,也是一种跨膜信号转换,只不过它们接受的外来刺激信号是电位变化,经过电压门控通道的开闭,再引起细胞膜出现新的电变化或其他细胞内功能变化,后者在Ca2+通道打开引起膜外Ca2+内流时甚为多见。
根据对Na+、K+、Ca2+三种离子的电压门控通道蛋白质进行的分子结构分析,发现它们一级结构中的氨基酸排列有相当大的同源性,说明它们属于同一蛋白质家族,与之有关的mRNA在进化上由同一个远祖基因演化而来。图2-8是与体内动作电位(见后)产生至关重要的Na+通道在膜内结构的模式图,它主要由一个较大的α-亚单位组成,分子量约260kd;有时还另有一个或两个小分子量的亚单位,分别称为β1和β2。但Na+通道的主要功能看来只靠α-亚单位即可完成。这个较长的α-单位肽链中包含了4个结构类似的结构域(domain,每个结构域大致相当于上述Ach门控通道中的一个亚单位,但结构域之间由肽链相连,是一个完整的肽链,应由一个mRNA编码和合成),而每个结构域中又各有6个由疏水性氨基酸组成的跨膜α-螺旋段(图示2-8,A);这4 个结构域及其所包含的疏水α-螺旋,在膜中包绕成一个通道样结构(图2-8,B)。现已证明,每个结构域中的第4个跨膜α-螺旋在氨基酸序列上有特点,即每隔两个疏水性氨基酸,就再现一个带正电荷的精氨酸或赖氨酸;这些α-螺旋由于自身的带电性质,在它们所在膜的跨膜电位有改变时会产生位移,因而被认为是该通道结构中感受外来信号的特异结构,由此再诱发通道“闸门”的开放;还有实验提示,每个结构域中的第2、第3个α-螺旋构成了该通道水相孔道的“内壁”;据测算,水相孔道内径最窄处横断面积约为0.3×0.5nm差不多刚能通过一个水化的Na+(图2-8,B)。
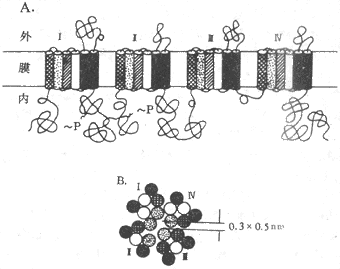
图2-8 电压门控Na+通道的分子结构示意图
A:构成电压门控Na+通道的α-亚单中的4个结构以及每个结构域中6个
α-螺旋在膜中存在形式平面 ~P表示磷酸化位点
B:4个结构域及其α-螺旋形成通道时的相对位置
(三)机械门控通道
体内存在不少能感受机械性刺激并引致细胞功能改变的细胞。如内耳毛细胞顶部的听毛在受到切和力的作用产生弯曲时,毛细胞会出现暂短的感受器电位,这也是一种跨膜信号转换,即外来机械性信号通过某种结构内的过程,引起细胞的跨膜电位变化。据精细观察,从听毛受力而致听毛根部所在膜的变形,到该处膜出现跨膜离子移动之间,只有极短的潜伏期,因而推测可能是膜的局部变形或牵引,直接激活了附近膜中的机械门控通道。
细胞间信道 还有一种通道,不是沟通胞浆和细胞外液的跨膜通道,而是允许相邻细胞之间直接进行胞浆内物质交换的通道,故称为细胞间通道。这种通道研究,是从缝隙连接超微结构观察开始的。在缝隙连接处相邻两细胞的膜仅隔开2.0nm左右,而且像是有某种物质结构把两者连接起来;将两侧细胞膜分离进行超微结构观察和分子生物学分析,发现每一侧的膜上都整齐地地排列着许多蛋白质颗粒,每个颗粒实际是由6个蛋白质亚单位(分子量各为25kd)构成的6聚体蛋白质,中间包绕一个水相孔道;构成颗粒的蛋白质和中心孔道贯穿所在膜的脂质双分子层;在两侧细胞膜靠紧形成细胞间的缝隙连接时,两侧膜上的各颗粒即通道样结构都两两对接起来,于是形成了一条条沟通两细胞胞浆的通路,而与细胞间液不相沟通。这种细胞间通道的孔洞大小,一般可允许分子量小于1.0~1.5kd或分子直径小于1.0nm的物质分子通过,这包括了电解质离子、氨基酸、葡萄糖和核苷酸等。这种缝隙连接或细胞间通道多见于肝细胞、心肌细胞、肠平滑肌细胞、晶状体细胞和一些神经细胞之间。缝隙连接不一定是细胞间的一种永久性结构;至少在体外培养的细胞之间的缝隙连接或其中包含颗粒的多少,可因不同环境因素而变化;似乎是细胞膜中经常有单方面装配好的通道颗粒存在,在两侧膜靠近并有其他调控因素存在时,就有可能实现对接,而在另一些因素存在时,两方面还可再分离。已对接的通道是否处于“开放”状态,也要受到多种因素的调控,例如当细胞内Ca2+、H+浓度增加时,可促使细胞间通道关闭。细胞间通道的存在,有利于功能相同而又密接的一组细胞之间进行离子、营养物质,甚至一些信息物质的沟通,造成它们进行同步性活动的可能性。
二、由膜的特异受体蛋白质、G-蛋白和膜的效应器酶组成的跨膜信号传递系统
这是另一类型的跨膜信号传递。最初是从对激素作用机制的研究开始的。60年代在研究肾上腺素引起肝细胞中糖原分解为葡萄糖的作用机制时,发现如果使肾上腺素单独和分离出的细胞膜碎片相互作用,可以生成一种分子量小、能耐热的物质,当把这种物质同肝细胞的胞浆单独作用时,也能引起其中糖原的分解,同肾上腺素作用于完整的肝细胞时有类似的效应。实验提示,在肾上腺素正常起作用时,它只是作用于肝细胞的膜表面。通过某种发生在膜结构中的过程,先在胞浆中生成一种小分子物质,后者再实现肾上腺素分解糖原的作用。这种小分子物质不久被证明是环-磷酸腺苷(即cAMP,环磷腺苷)。以后又陆续发现,很多其他激素类物质作用于相应的靶细胞时,都是先同膜表面的特异受体相结合,再引起膜内侧胞浆中cAMP含量的增加(有时是它的减少),实现激素对细胞内功能的影响。这样就把cAMP称作第二信使,这是相对于把激素分子这类外来化学信号看作第一信使而言的。
导致cAMP产生的膜结构内部的过程颇为复杂:它至少与膜中三类特殊的蛋白质有关。第一类是能与到达膜表面的外来化学信号作特异性结合的受体蛋白质,这是一些真正可以称作受体的物质。目前已用分子生物学的方法证明,它们是一些独立的蛋白质分子;已经确定的近100种这类受体,都具有类似的分子结构,也属于同一蛋白质家族:即它们都由约300~400个氨基酸残基组成,有一个较长的细胞外N-末端,接着在肽链中出现7个由22~28个主要为疏水性氨基酸组成的α-螺旋,说明这肽链至少要反复贯穿膜7次,形成一个球形蛋白质分子,还有一段位于膜内侧的肽链C-末端。目前认为,受体分子中第7个跨膜螺旋是能够识别、即能结合某种特定外来化学信号的部位;在受体因结合了特异化学信号而激活时,将进而作用于膜中另一类蛋白质,即G-蛋白质。
G-蛋白是鸟苷酸结合蛋白(guaninenucleotide-binding protein)的简称,也是存在于膜结构中的一类蛋白质家族,根据它们分子结构中少数氨基酸残基序列上的不同,已被区分出有数十种,但结构和功能极为相似。G-蛋白通常由α-、β-、和γ-3个亚单位组成;α-亚单位通常起催化亚单位的作用,当G-蛋白未被激活时,它结合了一分子的GDP(二磷酸鸟苷);当G-蛋白与激活了的受体蛋白在膜中相遇时,α-亚单位与GDP分离而又与一分子的GTP(三磷酸鸟苷)结合,这时α-亚单位同其他两个亚单位分离,并对膜结构中(位置靠近膜的内侧面)的第三类称为膜的效应器酶的蛋白质起作用,后者的激活(或被抑制)可以引致胞浆中第二信使物质的生成增加(或减少)。上述肾上腺素的作用,就是先由激素激活膜上相应的受体后,通过一种称为Gs(兴奋性G-蛋白)的G-蛋白的中介,激活了作为效应器酶的腺苷酸环化酶(图2-9箭头1),使胞浆中的ATP生成了起第二信使作用的cAMP(图2-9中箭头2)。由于第二信使物质的生成经过多级催化作用,少数几个膜外化学信号分子同受体的结合,就可能在胞浆中生成数目众多的第二信使分子,这是这种类型的跨膜信号传递的重要特点之一。
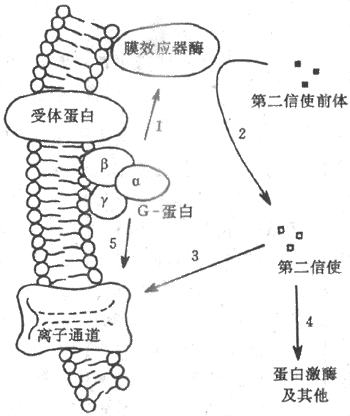
图2-9 由膜受体-G-蛋白-膜效应器酶组成的跨膜信号
传递系统和第二信使类物质的生成
目前发现膜的效应器酶并不只腺苷酸环化酶一种,因而第二信使物质也不只cAMP一种,如近年来还发现,有相当数量的外界刺激信号作用于受体后,可以通过一种称为Go的G-蛋白,再激活一种称为磷脂酶C的膜效应器酶,以膜结构中称为磷脂酰肌醇的磷脂分子为间接底物,生成两种分别称为三磷酸酰肌醇(IP3)和二酰甘油(DG)的第二信使,影响细胞内过程,完成跨膜信号传递。虽然如此,对应于细胞所能接受的多种刺激和与它们相对应的受体数目而言,膜内G-蛋白、效应器酶和最后生成的第二信使类物质的种类,还是相对地少得多。这说明,上述由膜中蛋白质酶促反应生成第二信使的途径,具有相当程度的“通用”性质。
由于上述这种跨膜信号传递的形式是在研究激素的作用机制时发现的,而且后来发现绝大多数肽类激素都是通过这一形式起作用的,因此曾一度错误地认为,这只是激素性化学信号跨膜信号传递方式。但近年的资料说明,事实并非如此:在神经递质类物质中,除了上述氨基酸类递质外,其余不论是小分子的经典递质还是后来发现的数量众多的神经肽类物质(目前已近50种),都主要是以在突触后细胞中产生第二信使类物质来完成跨膜信号传递的,这些第二信使物质通过在胞浆中的扩散,在膜的内侧面作用于某些特殊的离子通道(图2-9中箭头3),引起突触后膜较广泛而缓慢的电变化。最近证明,在视网膜信号转换过程中,光量子被作为受体的视色素如视紫红质(也具有7个跨膜α-螺旋的结构特点)吸收后,也是先激活称为Gt(转换蛋白)的G-蛋白,再激活作为效应器的磷酸二酯酶,使视杆细胞外段中cGMP的分解加强,最后使光刺激转变为外段膜的电变化(见第九章)。
上述两种主要的跨膜信号传递方式的作用过程,有以下几点值得注意。第一,这两种作用形式并不是绝对分离的,两者之间可以互相影响或在作用上有交叉。一些第二信使类物质可以调节某些电压门控通道和化学门控通道蛋白质的功能状态;而且被某种受体激活了的G-蛋白,有的不通过第二信使就能直接作用于膜结构中的通道结构(图2-9中的箭头5),如上述Gs激活时可以直接打开Ca2+通道。第二,对于许多外来化学信号分子,并不是一种化学信号只能作用于两种跨膜信号传递系统中的一种;以ACh为例,当它们作用于神经-肌接头处时,终板膜上有同它们作特异结合的化学门控通道;但当ACh作用于心肌或内脏平滑肌时,遇到的却是受体-G-蛋白-第二信使系统(受体称为M-型毒蕈硷型受体)。由此可见,同一种刺激信号通过何种跨膜信号传递系统起作用,关键因素在于靶细胞膜上具有何种感受结构;近年还发现,即便是M-型ACh受体,也可再区分出许多种亚型,有的亚型以cAMP为第二信使,有的以IP3和DG为第二信使。不同细胞甚或同一细胞的膜上具有对应于同一化学信号的不同受体型或其亚型,在跨膜信号传递中并不少见。近年来发现基本嗅觉刺激(大约是7种)全都是通过嗅上皮中不同的膜受体-第二信使系统起作用的,但在4种基本味觉刺激中,只有咸和酸刺激是通过细胞上相应的化学门控上通道起作用的,甜味物质是通过受体-第二信使系统起作用的,而苦味物质则因物质分子不同而分别通过通道和受体两种途径起作用。第三,跨膜信号传递的方式虽然相对地较少,但也不一定只限于上述两种。近年来有一些特殊的化学信号影响其靶细胞的方式受到广泛的重视,很可能成为跨膜信号传递的一种新类型;这就是发现胰岛素等一些肽类激素和其他与机体发育、生长、修复、增生、甚至细胞癌变有关的因子,如神经生长因子、表皮生长因子、血小板源生长因子、纤维母细胞生长因子、以及与细胞生成有关的集落刺激因子等,都是通过靶细胞表面一类称为酪氨酸激酶受体(tyrosine kinase riceptor)的蛋白质起作用的,这类受体结构简单,只有一个跨膜α-螺旋,当位于膜外侧的较长的肽链部分同特定的化学信号结合后,可以直接引起受体肽链的膜内段激活,使之具有磷酸激酶活性,通过使自身肽链和膜内蛋白质底物中的酪氨酸残基发生磷酸化,因而产生细胞内效应。这方面的新资料正在积累之中。
癌基因和跨膜信号传递近年发现与上述跨膜信号传递有关的一些蛋白质,如受体、G-蛋白、各种生长刺激因子和营养因子、以及各种蛋白激酶等,它们在细胞内的生物合成,是由人正常染色体中被称为细胞原癌基因(cellular proto-oncogene,进行表达时称细胞癌基因)的一类基因所编码和表达生成的。这些基因所以被称为原癌基因,是因为它们的硷基排列顺序同一些(近100种)能在动物引起肿瘤的病毒DNA(或称病毒癌基因,viral oncogene)的硷基排列顺序相一致。关于细胞癌基因与人类肿瘤发生的关系目前尚不清楚,但它们的正常表达产物,却是人体无时无刻不在进行着的各种跨膜信号传递过程所必需的。试设想,如果由于遗传和变异等原因使细胞不能合成结构和功能正常的G-蛋白,对人体将会有何等广泛而重要的影响!另外,在细胞原癌基因中,有一类可被胞浆中产生的第二信使等物质所激活,生成某种蛋白质;但它们在胞浆中生成后,一般又进入核内,进而诱导另一些基因进行表达。这类癌基因从激活到蛋白质生成,比一般基因表达为快,称为快速基因(或即早基因),而它们生成的蛋白质的作用则是激活另一些(可能在细胞功能活动中更重要)基因的表达,故快速基因的表达产物可称为转录调节因子或第三信使。所以称为第三信使是因为它们由第二信使类物质的作用而生成,而它们自身的作用又引起新的基因表达,生成一些可能对细胞结构和功能有较长远影响的蛋白质。这样外来信号(第一信使)不仅通过第二信使的合成在胞浆中引起一些即时反应,还可能通过第三信使引起细胞功能和结构长时间的适应性改变。
第三节 细胞的兴奋性和生物电现象
恩格斯在100多年前总结自然科学成就时指出:“地球几乎没有一种变化发生而不同时显示出电的现象”;生物体当然也不例外。事实上,在埃及残存史前古文字中,已有电鱼击人的记载;但对于生物电现象的研究,只能是在人类对于电现象一般规律和本质有所认识以后,并随着电测量仪器的精密化而日趋深入。目前,对健康人和患者进行心电图、脑电图、肌电图,甚至视网膜电图、胃肠电图的检查,已经成为发现、诊断和估量疾病进程的重要手段;但人体和各器官的电现象的产生,是以细胞水平的生物电现象为基础的,并且在生理学的发展历史上,生物电现象的研究是同生物组织或细胞的另一重要特性--兴奋性--的研究相伴随进行。
一、兴奋性和刺激引起兴奋的条件
(一)兴奋性和兴奋含义及其变迁
上世纪中后期的生理学家用两栖类动物做实验时,发现青蛙或蟾蜍的某些组织在离体的情况下,也能在一定的时间内维持和表现出某些生命现象。这些生命现象的表现之一是:当这些组织受到一些外加的刺激因素(如机械的、化学的、温热的或适当的电刺激)作用时,可以应答性出现一些特定的反应或暂时性的功能改变。这些活组织或细胞对外界刺激发生反应的能力,就是生理学最早对于兴奋性(excitability)的定义。例如,把蟾蜍的腓肠肌和支配它的神经由体内剥离出来,制成神经-肌肉标本,这时如果在神经游离端一侧轻轻地触动神经,或通以适当的电流,那么在经过一个极短的潜伏期后,可以看到肌肉出现一次快速的缩短和舒张;如把刺激直接施加于肌肉,也会引起类似的收缩反应;而且只要刺激不造成组织的损伤,上述反应可以重复出现。这就是神经和肌肉组织具有兴奋性能证明。实际上,几乎所有活组织或细胞都具有某种程度的对外界刺激发生反应的能力,只是反应的灵敏度和反应的表现形式有所不同。在各种动物组织中,一般以神经和肌细胞,以及某些腺细胞表现出较高的兴奋性;这就是说它们只需接受较小的程度的刺激,就能表现出某种形式的反应,因此称为可兴奋细胞或可兴奋组织。不同组织或细胞受刺激而发生反应时,外部可见的反应形式有可能不同,如各种肌细胞表现机械收缩,腺细胞表现分泌活动等,但所有这些变化都是由刺激引起的,因此把这些反应称之为兴奋(excitation)。人和高等动物的细胞和组织一样具有兴奋性,但在离体情况下要保持它们的兴奋性,需要严格的环境条件,因此在研究组织的兴奋性时,常用较低等动物的组织作为观察对象。
随着电生理技术的发展和资料的积累,兴奋性和兴奋的概念有了新的含义。大量事实表明,各种可兴奋细胞处于兴奋状态时,虽然可能有不同的外部表现,但它们都有一个共同的、最先出现的反应,这就是受刺激处的细胞膜两侧出现一个特殊形式的电变化(它由细胞本身所产生,不应与作为刺激使用的外加电刺激相混淆),这就是动作电位;而各种细胞所表现的其他外部反应,如机械收缩和分泌活动等,实际上都是由细胞膜的动作电位进一步触发和引起的。在神经细胞,特别是它的延续很长、起着信息传送作用的轴突(神经纤维),在受刺激而兴奋时并无肉眼可见的外部反应,其反应只是用灵敏的电测量仪器才能测出的动作电位。在多数可兴奋细胞(以神经和骨骼肌、心肌细胞为主),当动作电位在受刺激部位产生后,还可以沿着细胞膜向周围扩布,使整个细胞膜都产生一次类似的电变化。既然动作电位是大多数可兴奋细胞受刺激时共有的特征性表现,它不是细胞其他功能变化的伴随物,而是细胞表现其他功能的前提或触发因素,因此在近代生理学中,兴奋性被理解为细胞在受刺激时产生动作电位的能力,而兴奋一词就成为产生动作电位的过程或动作电位的同义语了。只有那些在受刺激时能出现动作电位的组织,才能称为可兴奋组织;只有组织产生了动作电位时,才能说组织产生了兴奋。这样的理解显然比原定义更严格些。
据此定义,可以对上述神经-肌标本的现象描述如下:当刺激作用于坐骨神经某一点时,由于神经纤维具有兴奋性而出现兴奋,即产生了动作电位,此动作电位(常称为神经冲动)沿着神经纤维传向它们所支配的骨骼肌纤维,通过神经-肌接头处的兴奋传递(即ACh参加的跨膜信号转换),再引起骨骼肌细胞兴奋而产生动作电位,以后是动作电位沿整个肌细胞膜传遍整个肌细胞,并触发了细胞内收缩蛋白质的相互作用,表现出肌肉一次快速的收缩和舒张。
(二)刺激引起兴奋的条件和阈刺激
具有兴奋性的组织和细胞,并不对任何程度的刺激都能表现兴奋或出现动作电位。刺激可以泛指细胞所处环境因素的任何改变;亦即各种能量形式的理化因素的改变,都可能对细胞构成刺激。但实验表明,刺激要引起组织细胞发生兴奋,必须在以下三个参数达到某一临界值:刺激的强度、刺激的持续时间以及刺激强度对于时间的变化率(即强度对时间的微分);不仅如此,这三个参数对于引起某一组织和细胞的兴奋并不是一个固定值,它们存在着相互影响的关系。在实验室中,常用各种形式的电刺激作为人工刺激,用来观察和分析神经或各种肌肉组织的兴奋性,度量兴奋性在不同情况下的改变。这是因为电刺激可以方便地由各种电仪器(如电脉冲和方波发生器等)获得,它们的强度、作用时间和强度-时间变化率可以容易地控制和改变;并且在一般情况下,能够引起组织兴奋的电刺激并不造成组织损伤,因而可以重复使用。
为了说明刺激的各参数之间的相互关系,可以先将其中一个参数固定于某一数值,然后观察其余两个的相互影响。例如,当使用方波刺激时,由于不同大小和持续时间的方波上升支都以同样极快的增加速率达到某一预定的强度值,因而可以认为上述第三个参数是固定不变的,而每一方波电刺激能否引起兴奋,就只决定于它所达到的强度和持续的时间了。在神经和肌组织进行的实验表明,在强度-时间变化率保持不变的情况下,在一定的范围内,引起组织兴奋所需的最小刺激强度,与这一刺激所持续的时间呈反变的关系;这就是说,当刺激的强度较大时,它只需持续较短的时间就足以引进组织的兴奋,而当刺激的强度较弱时,这个刺激就必须持续较长的时间才能引起组织的兴奋。但这个关系只是当所用强度或时间在一定限度内改变时是如此。如果将所用的刺激强度减小到某一数值时,则这个刺激不论持续多么长也不会引起组织兴奋;与此相对应,如果刺激持续时间逐资助缩短时,最后也会达到一个临界值,即在刺激持续时间小于这个值的情况下,无论使用多么大的强度,也不能引起组织的兴奋。
上述情况给比较不同组织的兴奋性高低或测量同一组织在不同生理或病理情况下的兴奋性改变时造成了许多困难。如果不仔细思考,可以认为那些用较小的刺激强度就能兴奋的组织具有较高的兴奋性;据上述,这个强度小的程度,还要决定这个刺激的持续时间和它的强度-时间变化率。因此,简单地用刺激强度这一个参数表示不同组织兴奋性的高低或同一组织兴奋性的波动,就必须使所用刺激的持续时间和强度-时间变化率固定某一(应是中等程度的)数值;这样,才能把引起组织兴奋、即产生动作电位所需的最小刺激强度,作为衡量组织兴奋性高低的指标;这个刺激强度称为阈强度或阈刺激,简称阈值(threshold)。强度小于阈值的刺激,称为阈下刺激;阈下刺激不能引起兴奋或动作电位,但并非对组织细胞不产生任何影响。
(三)组织兴奋及其恢复过程中兴奋性的变化
体内不同组织具有不同的兴奋性;而且同一组织在不同生理和病理情况下,强环境中离子成分特别是钙离子、酸碱度、温度的改变,以及存在着特殊毒物或药物等情况,都可以引起兴奋性的改变。但一个普遍存在于各种可兴奋细胞的现象是,在细胞接受一次刺激而出现兴奋的当时和以后的一个短时间内,它们的兴奋性将经历一系列有次序的变化,然后才恢复正常。这一特性说明,在细胞或组织接受连续刺激时,有可能由于它们接受前一刺激而改变了对后来刺激的反应能力,因而是一个有重要功能意义的生理现象。
为了示证这一特性,可以让两个刺激连续作用于组织,这时让第一个刺激的强度相当于阈强度,以便使它能引起组织兴奋,并以此阈强度的值作为该组织兴奋性的“正常”对照值;对于第二个刺激,在实验中要能任意地选定它们和第一刺激的间隔,并且可以按需要改变它们的强度。这样,可以检查组织在因第一个刺激后的不同时间内,接受新刺激的能力是否发生了改变。实验证明,在组织接受前面一个刺激而兴奋后一个较短的时间内,无论再受到多么强大的刺激,都不能再产生兴奋;即在这一时期内出现的任何刺激均“无效”;这一段时期,称为绝对不应期。在绝对不应期之后,第二个刺激有可能引起新的兴奋,但使用的刺激强度必须大于该组织正常的阈强度;这个时期称为相对不应期。上述绝对和相对不应期的存在,反映出组织在一次兴奋后所经历的兴奋性改变的主要过程;即在绝对不应期内,由于阈强度成为无限大,故此时的兴奋性可认为下降到零;在相对不应期内,兴奋性逐渐恢复,但仍低于正常值,此时需使用超过对照阈强度的刺激强度,才能引起组织的兴奋;到相对不应期结束时,兴奋性才逐渐恢复到正常。用更精密的实验发现,在相对不应期内之后,组织还经历了一段兴奋性先是轻度增高,继而又低于正常的时期,分别称为超常期和低常期。以上各期的长短,在不同细胞可以有很大差异;一般绝对不应期较短,相当于或略短于前一刺激在该细胞引起的动作电位主要部分的持续时间,如它在神经纤维或骨骼肌只有0.5~2.0ms左右,在心肌细胞可达200~400ms;其他各期的长短变化较大,易受代谢和温度等因素的影响。在神经纤维,相对不应期约持续数毫秒,超常期和低常期可达30~50ms。
组织在每次兴奋后都要发生一系列兴奋性的改变,如果在这期间组织受到新的刺激,它的反应能力将异于“正常”。既然绝对不应期的持续时间相当于前次刺激所引起的动作电位主要部分的持续时间,那么在已有动作电位存在的时期就不可能产生新的兴奋或动作电位,亦即细胞即便受到连续的快速刺激,也不会出现两次动作电位在同一部位重合的现象;由于同样的理由,不论细胞受到频率多么高的连续刺激,它在这一细胞所能引起的兴奋或动作电位的次数,总不会超过某一个最大值;因为落于前一刺激所产生的绝对不应期内的后续刺激将“无效”,因此这个最大值理论上不可能超过该细胞和组织的绝对不应期的倒数。例如,蛙的有髓神经纤维的绝对不应期或动作电位的持续时间约为2ms,那么此纤维每秒钟内所能产生的动作电位的次数不可能超过500;实际上神经纤维在体内自然情况下所能产生和传导的神经冲动的频率,远远低于它们理论上可能达到的最大值。
二、细胞的生物电现象及其产生机制
(一)生物电现象的观察和记录方法
前已指出,神经在接受刺激时,虽然不表现肉眼可见的变化,在受刺激的部位产生了一个可传导的电变化,以一定的速度传向肌肉,这一点可以用阴极射线示波器为主的生物电测量仪器测得,如图2-10所示。图中由射线管右侧电子枪形成的电子束连续射向荧光屏,途中经过两对板状的偏转电极;当电子束由水平偏转板两极之间通过时,由于板上有来自扫描发生器装置的锯齿形电压变化,使射向荧光屏的电子束以一定的速度作水平方向的反复扫动;这时,如果把由两个测量电极引导来的生物电变化经放大器放大后加到垂直偏转板的两极,那么电子束在作横扫的同时又作垂直方向的移动。这样,根据移动电子束在荧光屏上形成的光点的轨迹,就能准确地测量出组织中的微弱电变化的强度及其随时间变化的情况。如果神经干在右端受到刺激,神经纤维将产生一个传向左端的动作电位,当它传导到同放大器相导到同放大器相连的第一个引导电极处时,该处的电位暂时变得相对地较负,于是在一对垂直偏转板上再现电位差,在荧光屏上可看到一次相应的光点波动;当动作电位传导到第二个引导电极处时,该处也将变得较负,于是荧光屏上会出现另一次方向相反的光点波动;这样记到的两次电位波动,称作双相动作电位。把神经标本作一些特殊处理,如将第二个记录电极下方的神经干损伤(如图2-10所示),使该处不能产生兴奋,那么再刺激神经右端时,在示波器上只能看到一次电位波动,这称为单相动作电位。另外,用其他技术方法还可使记录电极中的一个电极处的电位保持恒定或经常处于零电位状态,亦即使此电极成为参考或无关电极,于是在实验中记录到的电变化就只反映与另一电极(称为有效电极)接触处的组织或细胞的电变化,这称为单极记录法。
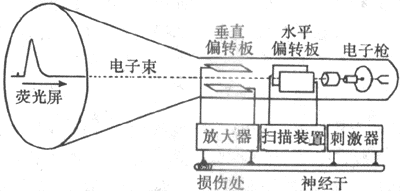
图2-10 用阴极射线示波器及有关设备观察生物电现象的基本实验布置
(二)细胞的静息电位和动作电位
双相或单相动作电位,是在神经干或整块肌肉组织上记录到的生物电现象,是许多在结构和功能上相互独立的神经纤维或肌细胞的电变化的复合反映;由于测量电极和组织有较大的接触面积,而且组织本身又是导电的,许多细胞产生的电变化可被同一电极所引导,所以记录和测量出的电变化是许多单位的电变化和代数叠加。但目前已经确知,生物电现象是以细胞为单位产生的,是以细胞膜两侧带电离子的不均衡分布和选择性离子跨膜转运为基础的。因此,只有在单一神经或肌细胞进行生物电的记录和测量,才能对它的数值和产生机制进行直接和深入的分析。由于一般的细胞纤小脆弱,单一细胞生物电是通过以下方法测量的:一是利用某些无脊椎动物特有的巨大神经或肌细胞,如枪乌贼的神经轴突,其直径最大可达100μm左右,便于单独剥出进行实验观察,脊椎动物的单一神经纤维也可以设法剥出,但它们的直径最粗也不过20μm左右,方法上较为困难。另一种方法是进行细胞内微电极记录,即用一个金属或细玻璃管制成的充有导电液体而尖端直径只有1.0μm或更细的微型记录电极(凌宁和Gerard,1949),由于它只有尖端导电,可用它刺入某一个在体或离体的细胞或神经纤维的膜内,测量细胞在不同功能状态时膜内电位和另一位于膜外的参考电极之间的电位差(即跨膜电位),这样记录到的电变化,只与该细胞有关而几乎不受其他细胞电变化的影响。
细胞水平的生物电现象主要有两种表现形式,这就是它们在安静时具有的静息电位和它们受到刺激时产生的动作电位。体内各种器官或多细胞结构所表现的多种形式的生物电现象,大都可以根据细胞水平的这些基本电现象来解释。
静息电位指细胞未受刺激时存在于细胞内外两侧的电位差。测量细胞静息电位的方法如图2-11所示。R表示测量仪器如示波器,和它相连的一对测量电极中有一个放在细胞的外表面,另一个连了微电极,准备刺入膜内。当两个电极都处于膜外时,只要细胞未受到刺激或损伤,可发现细胞外部表面各点都是等电位的;这就是说,在膜表面任意移动两个电极,一般都不能测出它们之间有电位差存在。但如果让微电极缓慢地向前推进,让它刺穿细胞膜进入膜内,那么在电极尖端刚刚进入膜内的瞬间,在记录仪器上将显示出一个突然的电位跃变,这表明细胞膜内外两侧存在着电位差。因为这一电位差是存在于安静细胞的表面膜两侧的,故称为跨膜静息电位,简称静息电位。
在所有被研究过的动植物细胞中(少数植物细胞例外),静息电位都表现为膜内较膜外为负;如规定膜外电位为0,则膜内电位大都在-10~-100mV之间。例如,枪乌贼的巨大神经轴突和蛙骨骼肌细胞的静息电位为-50~-70mV,哺乳动物的肌肉和神经细胞为-70~-90mV,人的红细胞为-10mV,等等。静息电位在大多数细胞是一种稳定的直流电位(一些有自律性的心肌细胞和胃肠平地滑肌细胞例外),只要细胞未受到外来刺激而且保持正常的新陈代谢,静息电位就稳定在某一相对恒定的水平。
在近代生理学文献中,一些过去单纯用来描述膜两侧电荷分布状态的术语,仍被用来说明静息电位的存在及其可能出现的改变。例如,人们常常把静息电位存在时膜两侧所保持的内负外正状态称为膜的极化(polarization),原意是指不同极性的电荷分别在膜两侧的积聚;当静息电位的数值向膜内负值加大的方向变化时,称作膜的超级化(hyperpolarization);相反,如果膜内电位向负值减少的方向变化,称作去极化或除极(depolarization);细胞先发生去极化,然后再向正常安静时膜内所处的负值恢复,则称作复极化(repolarization)。
现通过图2-11中的实验布置,观察单一神经纤维动作电位的产生和波形特点,由图中可见,当神经纤维在安静状况下受到一次短促的阈刺激或阈上刺激时,膜内原来存在的负电位将迅速消失,并且进而变成正电位,即膜内电位在短时间内可由原来的-70~-90mV变到+20~+40mV的水平,由原来的内负外正变为内正外负。这样,整个膜内外电位变化的幅度应是90~130mV,这构成了动作电位变化曲线的上升支;如果是计算这时膜内电位由零值变正的数值,则应在整个幅值中减去膜内电位由负上升到零的数值,在图2-11中约为35mV,即动作电位上升支中零位线以上的部分,称为超射值。但是,由刺激所引起的这种膜内外电位的倒转只是暂时的,很快就出现膜内电位的下降,由正值的减小发展到膜内出现刺激前原有的负电位状态,这构成了动作电位曲线的下降支。由此可见,动作电位实际上是膜受刺激后在原有的静息电位基础上发生的一次膜两侧电位的快速而可逆的倒转和复原;在神经纤维,它一般在0.5~2.0ms的时间内完成,这使它在描记的图形上表现为一次短促而尖锐的脉冲样变化,因而人们常把这种构成动作电位主要部分的脉冲样变化,称之为锋电位。在锋电位下降支最后恢复到静息电位水平以前,膜两侧电位还要经历一些微小而较缓慢的波动,称为后电位,一般是先有一段持续5~30ms的负后电位,再出现一段延续更长的正后电位,如图2-11下所示(这里负后和正后电位两个术语仍沿用动作电位细胞外记录时的命名;确切地说,负后电位应称为去极化后电位,而正后电位应称为超极化后电位)。锋电位存在的时期就相当于绝对不应期,这时细胞对新的刺激不能产生新的兴奋;负后电位出现时,细胞大约正处于相对不应期和超常期,正后电位则相当于低常期。
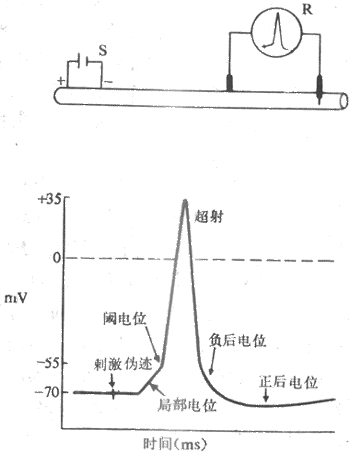
图 2-11 单一神经纤维静息电位和动作电位的实验模式图
R表示记录仪器,S是一个电刺激器。当测量电极中的一个
微电极刺入轴突内部时可发现膜内持续处于较膜外低70mV的负电位状态。
当神经受到一次短促的外加刺激时,膜内电位快速上升到+35mV的水平,
约经0.5~1.0ms后再逐渐恢复到刺激前的状态。其他说明见正文
动作电位或锋电位的产生是细胞兴奋的标志,它只在刺激满足一定条件或在特定条件下刺激强度达到阈值时才能产生。但单一神经或肌细胞动作电位产生的一个特点是,只要刺激达到了阈强度,再增加刺激并不能使动作电位的幅度有所增大;也就是说,锋电位可能因刺激过弱而不出现,但在刺激达到阈值以后,它就始终保持它某种固有的大小和波形。此外,动作电位不是只出现在受刺激的局部,它在受刺激部位产生后,还可沿着细胞膜向周围传播,而且传播的范围和距离并不因原初刺激的强弱而有所不同,直至整个细胞的膜都依次兴奋并产生一次同样大小和形式的动作电位。图2-11的实验布置中,神经受刺激部位和记录部位之间有一段距离;但不论记录电极在职一神经纤维上如何移动(除非是在纤维末梢处有了纤维形态的改变,或纤维的离子环境等因素发生了改变),我们一般都能记录到同样大小和波形的锋电位,所不同的只是刺激伪迹和锋电位之间的间隔有所变化,这显然与动作电位在神经纤维上“传导”到记录电极所在部位时所消耗的时间长短有关。这种在同一细胞上动作电位大小不随刺激强度和传导距离而改变的现象,称作“全或无”现象,其原因和生理意义将在下面讨论。
在不同的可兴奋细胞,动作电位虽然在基本特点上类似,但变化的幅值和持续时间可以各有不同。例如,神经和骨骼肌细胞的动作电位的持续时间以一个或几个毫秒计,而心肌细胞的动作电位则可持续数百毫秒;虽然如此,这些动作电位都表现“全或无”的性质。
(三)生物电现象的产生机制
早在1902年,Bernstein就提出膜学说,他根据当时关于电离和电化学的理论成果提出了经典的膜学说来解释当时用粗劣的电测量仪器记录到的生物电现象。他认为细胞表面膜两侧带电离子的不同分布和运动,是产生物电的基础。但在当时和以后相当长的一段时期内,还没有测量单一细胞电活动的手段和其他有关技术,因此他的学说长期未能得到证实。直到本世纪40~50年代,Hodgkin 和Huxley等开始利用枪乌贼的巨大神经轴突和电生理学技术,进行了一系列有意义的实验,不仅对经典膜学说关于静息电位产生机制的假设予以证实,而且对动作电位的产生作了新的解释和论证。通过这一时期的研究,对于可兴奋细胞静息电位和动作电位的最一般原理已得到阐明,即细胞生物电现象的各种表现,主要是由于某些带电离子在细胞膜两侧的不均衡分布,以及膜在不同情况下对这些离子的通透性发生改变所造成的。但是由于当时对细胞膜的分子结构和膜中蛋白质的存在形式和功能还知之甚少,因此Hodgkin等对生物电的理解只能是宏观的,对微细过程只能用数学模型来说明。随着70年代以来蛋白质化学和分子生物学技术的迅速发展,蛋白质分子从膜结构中克隆出来,并从它们的分子结构的特点来说明通道的功能特性;特别是70年代中期发展起来的膜片钳(patch clamp)技术,可以观察和记录单个离子通道的功能活动,使宏观的所谓膜对离子通透性或膜电导的改变,得到了物质的、可测算的证明。
1.静息电位和K+平衡电位Bernstein最先提出,细胞内外钾离子的不均衡分布和安静状态下细胞膜主要对K+有通透性,可能是使细胞能保持内负外正的极化状态的基础。已知所有正常生物细胞细胞内的K+浓度超过细胞外K+很多,而细胞外Na+浓度超过细胞内Na+浓度很多,这是Na+泵活动的结果;在这种情况下,K+必然会有一个向膜外扩散的趋势,而Na+有一个向膜内扩散趋势。假定膜在安静状态下只对K+有通透的可能,那么只能有K+移出膜外,这时又由于膜内带负电荷的蛋白质大分子不能随之移出细胞,于是随着K+移出,出现膜内变负而膜外变得较正的状态。K+的这种外向扩散并不能无限制地进行,这是因为移到膜外的K+所造成的外正内负的电场力,将对K+的继续外移起阻碍作用,而且K+移出的愈多,这种阻碍也会愈大。因此设想,当促使K+外移的膜两侧K+浓度势能差同已移出K+造成的阻碍K+外移的电势能差相等,亦即膜两侧的电-化学(浓度)势代数和为零时,将不会再有K+的跨膜净移动,而由已移出的K+形成的膜内外电位差,也稳定在某一不再增大的数值。这一稳定的电位差在类似的人工膜物理模型中称为K+平衡电位。Bernstein用这一原理说明细胞跨膜静息电位的产生机制。不难理解,K+平衡电位所能达到的数值,是由膜两侧原初存在K+浓度差的大小决定的,它的精确数值可根据物理化学上著名的Nernst公式(1889)算出:
![]() (1)
(1)
(1)式中Ek表示K+平衡电位,R是通用气体常数,Z是离子价,F是Farady常数,T是绝对温度;式中只有[K+]o和[K+]i是变数,分别代表膜两侧的K+浓度。如果把有关数值代入,室温以27°С计算,再把自然对数化为常用对数,则式(1)可简化为;(2)
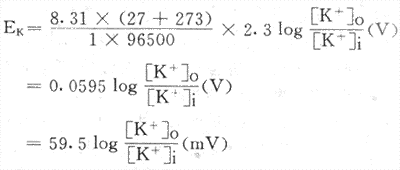 (2)
(2)
如果,Bernstein应用当时物理化学最新成果说明细胞静息电位产生机制的理论是正确的,那么在细胞实际测得的静息电位的数值,应相当于把当时细胞内外K+浓度值代入式(2)时计算所得的Ek值。1939年Hodgkin等利用了枪乌贼的巨大神经纤维和较精密的示波器等测量仪器,第一次精确地测出此标本的静息电位值,结果发现此值和计算所得的K+平衡电位值非常接近而略小于后者;如在一次实验中测得的静息电位值为-77mV,而按当时[K+]o和[K+]i值算出的Ek为-87mV,基本上符合膜学说关于静息电位产生机制的解释。
为了进一步证实这一理论,Hodgkin等又用人工地改变标本浸溶液中K+浓度即[K+]o,因而也改变了[K+]o/[K+]i值的实验方法,观察到所记录的静息电位的什也随[K+]o的改变而改变,而改变的情况基本上同根据式(2)计算出的预期值相一致。随后用微电极细胞内记录法在纤细的哺乳类标本也进行了类似的实验,得到类似的结果,如在骨骼肌细胞测得的静息电位为-90mV,而计算所得的Ek值为-95mV。这些实验都说明,大多数细胞的静息电位的产生,是由于正常细胞的细胞内液高K+而膜在安静时又主要对K+有通透能力的结果;至于静息电位的数值为何略小于理论上的Ek值,一般认为是由于膜在静息时对Na+也有极小的通透性(大约只有K+通透性的1/50~1/100)的缘故;由于膜外Na+浓度大于膜内,即使小量的Na+逸入膜内也会抵消一部分K+外移造成的膜内负电位。
2.锋电位和Na+平衡电位Hodgkin等根据兴奋时膜内不仅出现负电位的消失,而且出现一定数值的正电位(相当于前面提到的超射值)的事实,因而认为对动作电位上升支的出现,不能像Bernstein那样简单地解释为膜对K+通透性的消失,因为这样最多也只能使膜内原有的负电位回升到零。他们据此设想膜在受到刺激时可能出现了膜对Na+通透性的突然增大,超过了K+的通透性,由于细胞外高Na+,而且膜内静息时原已维持着的负电位也对Na+的内流起吸引作用,于是Na+迅速内流,结果先是造成膜内负电位的迅速消失;而且由于膜外Na+的较高的浓度势能,Na+在膜内负电位减小到零电位时仍可继续内移,直至内移的Na+在膜内形成的正电位足以阻止Na+的净移入时为止。不难设想,这时膜内所具有的电位值,理论上应相当于根据膜内外Na+浓度差代入Nernst公式时所得出的Na+平衡电位值(可写为ENa)。实验数据证明,动作电位所能达到的超射值,即膜内正电位的数值,正相当于计算所得的ENa;而且实验中随着标本浸溶液中Na+被同等数目的葡萄糖分子所代替(使[Na+]o逐渐减小),可以看到所能记录到的动作电位的超射值和整个动作电位的幅度也逐渐减小,其程度也同按Nernst公式算出的预期值基本一致。
但是,膜内电位停留在ENa水平的时间极短;随后很快出现膜内电位向静息时的状态恢复,亦即出现复极,造成了锋电位曲线的快速下降支。如后来的实验证明,这下降支的出现是由于Na+通透性的消失,并伴随出现了K+通透性的增大。
细胞每兴奋一次或产生一次动作电位,总有一部分Na+在去极化时进入膜内,一部分K+在复极时逸出膜外,但由于离子移动受到各该离子的平衡电位的限制,它们的实际进出量是很小的;据估计,神经纤维每兴奋一次,进入膜内的Na+量大约只能使膜内的Na+浓度增大约八万分之一,复极时逸出的K+量也类似这个数量级;即便神经连续多次产生兴奋,短时间内也不大可能明显地改变膜内高K+和膜外高Na+这种基本状态,而只要这种不均衡离子分布还能维持,静息电位就可以维持,新的兴奋就可能产生。细胞膜两侧K+、Na+离子的不均衡分布,主要是靠钠泵蛋白质消耗代谢能建立起来的,而由此形成的势能贮备却可供细胞多次产生兴奋而不需当时耗氧供能。不过实际上钠泵的活动又受膜内外Na+、K+浓度的调控,它对膜内Na+浓度增加十分敏感,Na+的轻微增加就能促使钠泵的活动,因此在每次兴奋后的静息期内,都有钠泵活动的一定程度的增强,将兴奋时多进入膜内的Na+泵出,同时也将复极时逸出膜外的K+泵入,使兴奋前原有的离子分布状态得以恢复。这时由于两种离子的转运同时进行,出入的离子总数又近于相等,故一般不伴有膜两侧电位的明显改变。但在膜内Na+蓄积过多而使钠泵的活动过度增强时,上述的定比关系可以改变,结果是泵出的Na+量有可能明显超过泵入的K+量,这就可能使膜内负电荷相对增多,使膜两侧电位向超极化的方向变化;这时的钠泵,就称为生电性钠泵。有人认为,锋电位以后出现的正后电位,是由于生电性钠泵作用的结果。至于负后电位,则一般认为是在复极时迅速外流的K+蓄积在膜外侧附近,因而暂时阻碍了K+外流的结果。
3.经典的电压钳(或电压固定)实验从上述可知,Hodgkin等对于动作电位产生机制的说明,关键在于膜受刺激时对Na+、K+的通透性发生了有选择而时间亦有先后的改变,但这只是根据所测得的膜内外电位改变对照Nernst公式进行的推论,实验并没有对膜的通透性进行直接的测量和动态描述。为此,他们又应有当时最先进的电子学技术,设计和进行了著名的电压钳(voltage clamp)实验。实验的设计根据是:离子作跨膜移动时形成了跨膜离子电流(I),而通透性亦即离子通过膜的难易程度,就是膜的电阻(R)或其倒数电导(G),因此所谓膜对某种离子通透性增大时,实际是膜对该离子的电导加大;对于带电离子来说,膜电导就是膜通透性的同义语。根据欧姆定律,I=VG,可知在膜两侧电位差(V)固定不变的条件下,测出的跨膜电流I的变化,就可作为膜电导变化的度量。测定膜在受刺激时跨膜电流的改变在技术上是容易的,但在这过程中要保持膜电位固定不变却不容易;因为当存在跨膜离子电流时,离子的进出膜会使不导电而有电容(C)特性的脂质膜充电或放电,因而根据V=Q/C的关系(其中Q为电量,相当于I和时间t的乘积),跨膜离子的移动必然要引起跨膜电位的改变;实际上记录到的动作电位就是这种改变。正因为如此,Hodgkin等自行设计了一种应用负反馈原理的电子学装置,使它们能在跨膜电位维持恒定(恒定的数值可由实验者通过实验装置预先设定)的情况下,测量跨膜离子电流的强度改变,并由此计算出膜电导即膜通透性的变化情况。电压钳实验的基本原理模式图如图2-12所示。图中电极1插入巨大神经轴突内一定距离,用来测量和监察这一段轴突膜内的电位,此电极先连到一个电压放大器,再在一个示波器上显示;电极1测得电位值经放大后同时输给一个负反馈放大器(FBA),这是整个仪器设计的关键部分,它可把测得的膜内电位同来自一个电压源的、由实验者预先设定的要求保持恒定的电位值进行比较,如果二者有差值,FBA就会通过电极2向轴突膜内输出相应强度和方向的电流,由于仪器线路的精密设计和快速反应,电极2输出电流的改变正足以补偿标本由于跨膜离子电流使膜充放电而引起的跨膜电位的变动,于是与电极1相边的示波器上显示出膜内电位固定在设定的数值,而在电流放大器IA上测得的跨膜离子电流的变化,就反映了膜电导的变化。
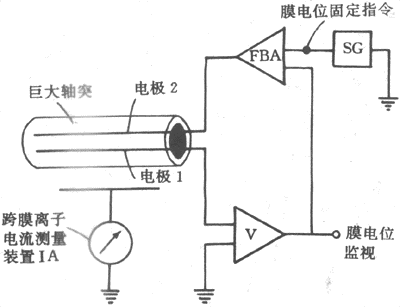
图 2-12 电压钳实验布置模式图
电压固定实验获得了许多有意义的结论。首先一点是,只有设定的膜内电位固定在去极化水平时,才有可能出现膜的Na+电导(GNa)和K+电导(Gk)的增大,并且设定电位愈接近零值,电导的增大也愈明显;相反,如果设定的膜内电位值是超极化的,则不可能引起跨膜离子电流和膜电导的改变,这一点以后还要谈到。以图2-13的记录曲线为例,分析不同离子的电导在一次兴奋过程中的变化情况。图中最上方曲线表示在一次电压钳实验中,把膜内电位由静息时的-65mV突然固定(这就是(clamp)的意思)在-9mV,结果很快引起一次如曲线A的跨膜电流变化曲线,这曲线的开始部分是内向的,以后逐渐转变为外向电流。只记录到内向或外向电流还不能说明电荷的携带者是何种离子,根据过去的实验者有理由认为,先出现的内向电流可能是Na+电流(INa),外向电流则可能是K+电流(Ik)。用附加的实验观察证明了这点:假定把标本浸浴液中的NaCI用相同摩尔数的氯化胆碱来代替,则在同样的条件下只能记录到较晚出现的曲线B,它是外向的,这显然是因为不能出现内向的INa的结果;把曲线A和B逐点相减,就能得到曲线C,它就是内向的INa;由INa、Ik两条曲线,就可算出GNa和Gk的变化曲线,其特点是:(1)GNa和Gk都是电压依从性的,只能由跨膜电位的去极化所激活,但GNa被激活得早,是动作电位上升支出现的基础,而Gk激活出现缓慢,是动作电位复极到静息电位水平的基础;(2)GNa有失活(inactivation)状态而Gk没有此特性,其证明是图2-13中曲线C只存在1~2ms,以后跨膜电压虽仍固定在-9mV的水平,但GNa早已恢复到原初水平,而代表Gk的曲线B虽然出现较晚,但它在设定电位持续期间一直维持在较同的水平。GNa失活的出现和Gk的激活是造成神经纤维和骨骼肌细胞表现短促的锋电位的原因;在膜复极以后GNa的失活状态才能消失,这时GNa才能因膜的去极化而再出现增大。
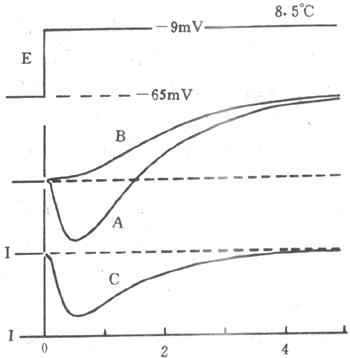
图2-13 电压钳实验结果示意图
将巨大神经纤维的膜电位由原来的-65mv突然上升并固定于-9mv的水平时,
膜的离子电流的变化情况(曲线A、B、C的意义见正文)
根据图2-13中INa和Ik两条电流曲线,即可计算出同这两者相对应的GNa和Gk曲线,再根据这一段膜所具有的电容的数值(有人测得每cm2的枪乌贼轴突膜的电容约为1μF),就可算出如果“允许”每一瞬间的离子移动在电容上形成电位改变时,有可能造成怎样的跨膜电位的改变,这正是不进行“电压固定”时的情况,而由此作出的电位变化曲线正好同在一般实验中记录到的动作电位的波形特点一致,如图2-14所示。这进一步说明了电压钳实验证明动作电位产生机制的正确性。
4.膜片钳实验和单通道离子电流的记录通过上节关于电压门控通道的特性分析已知,所谓膜对某种离子通透性的改变,实际上决定于膜结构中有关离子通道蛋白质分子的功能状态;例如,Hodgkin等测出的GNa的变化,实际是那一段轴突膜上众多的电压门控式Na+通道因膜的去极化而开放的结果。在Hodgkin等当时进行的膜电导改变的数学模拟中,已经明确提示,GNa和Gk的改变不是均匀地发生在整个膜平面上,而是与膜上某些特定的“点”有关,不久又发现,有些药物可以选择性地阻断某种离子的跨膜移动,如河豚毒可以单独阻断GNa而不影响Gk,四乙基铵可以单独阻断Gk而不影响GNa;以同位素标记的河豚毒只能与膜上某些特殊的“点”作特异性结合,而标记的四乙基铵只能与另一些“点”结合。这些实验以及兴奋过程中离子移动数目之多与快,逐渐使人们推断膜结构中有特殊的蛋白质离子通道的存在。这说明,“通道”概念的提出,远在通道的实质被阐明以前,是前者促进了对后者的进一步探索。70年代中期由Neher和Sakmann等发展出一种能够记录膜结构中单一的离子通道蛋白质分子的开放和关闭、亦即测量单通道离子电流和电导的技术,称为膜片钳实验。
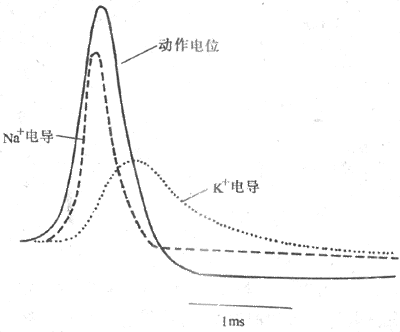
图 2-14 电导变化与电位变化的关系示意图
根据电压钳实验中测得的Na+电导(GNa)和K+电导(Gk)的变化过程,
可以算出在膜电位不进行人为固定时,相应的Na+、K+离子电流在膜电容
上引起的电位变化(实线),其形状正同在标本上记录到的动作电位的波形一致
膜片钳实验的基本原理如图2-15A所示:用一个尖端光洁、直径约0.5~3μm的玻璃微电极同神经或肌细胞的膜接触而不刺入,然后在微电极另一端开口施加适当的负压,将与电极尖端接触的那一小片膜轻度吸入电极尖端的纤细开口,这样在这小片膜周边与微电极开口处的玻璃边沿之间,会形成紧密的封接,在理想的情况下其电阻可达数个或数十千兆欧(其物理过程目前尚不清楚),这实际上把吸附在微电极尖端开口处的那一小片膜同其余部分的膜在电学上完全隔离开来;如果在这一小片膜中只包含了一个或少数几个通道蛋白质分子,那么通过此微电极就可能测量出单一通道开放时的离子电流和电导,并能对单通道的其他功能特性进行分析。

图2-15 膜片钳实验布置示意图
A:图中Ip为记录到的单通道电流,VCMD决定设定的膜电位数值
B:在大鼠胚胎骨骼肌细胞膜片上记录到的由ACH激活的单通道
离子电流,强度为pA(皮安)级
从Neher等最初用膜片钳技术观察骨骼肌终板膜处的单一ACh-门控通道机能特性开始,已经对多种通道进行了观察,发现它们一般有如下共同特性:(1)不论是化学门控或电压门控通道,它们的开放和关闭都是突然的,使描绘出的电流曲线呈方波状,说明相应的蛋白质分子可以从一种构象快速地跃变到另一种构象;(2)每种通道开放时具有恒定的电导,即在恒定的情况下,只能看到“开”或“关”两种状态,很少看到“半开”或“部分开”的情况;(3)即使是同一通道分子,每次开放的持续时间长短也不一致,似乎说明蛋白质分子可在开放和关闭两种构象之间“摆动”,停留在某种状态的长短具有随机的性质;(4)在化学门控通道结合了相应的化学信号分子,或电压门控通道所在膜两侧处于特定的电位差的情况下,“摆动”的次数增多,开放的机率增大,而“失活”使开放的机率减小。
用单通道记录可说明在自然情况下整段膜的离子电导和离子电流的形成机制;以上述GNa增大为例,它显然是该段膜中众多的Na+通道在去极化的影响下出现开放的机率增加所决定的,而在每一瞬间同时出现的各通道的电导或离子电流相互叠加,于是如图2-16B所示,这种叠加形成的Na+电流曲线,正好和图2-13中的曲线C相似。
膜片钳实验可用于各种细胞,由于微电极不刺入细胞,即使用于纤小的细胞也不致造成损伤。膜片钳实验已有各种变式,如吸着在微电极尖端的小膜片可以随电极而同原细胞脱离,把它们浸入人工浸浴液中,就可以观察某些因素在膜的胞浆侧怎样影响通道功能;也可以形成膜的胞浆侧面向微电极尖端开口而膜表面侧面向浸浴液的实验模式,等等。膜片钳实验也已用于细胞生物电以外的功能研究,如细胞的分泌过程等。
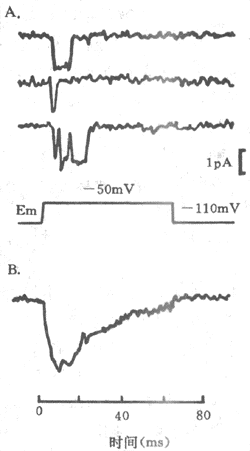
图2-16 电压门控Na+通道的膜片钳记录A:
随着静息电位(Em)由-110mV突然固定到-50mV,
在3次膜片钳实验记录到的离子电流 B:将144次膜片钳记录
到的离子电流曲线进行平均叠加,得到一条类似图
2-13中曲线C的Na+电流曲线,说明后者是多数Na+通道激活的结果
三、兴奋的引起和兴奋的传导机制
(一)阈电位和锋电位的引起
膜内负电位必须去极化到某一临界值时,才能在整段膜引发一次动作电位,这个临界值大约比正常静息电位的绝对值小10~20mV,称为阈电位。例如,巨大神经轴突的静息电位为-70mV,它的阈电位约为-55mV。这不是由于小于阈电位的去极化不引起GNa的增加,实际情况是这时也有一定数目的Na+通道开放,但由于膜对K+的通透性仍大于Na+,因而少量的Na+内流及其对膜内电位的影响随即被K+的外流所抵消,因而去极化不能继续发展下去,不能形成动作电位。只有当外来刺激引起的去极化达到阈电位水平时,由于较多量Na+通道的开放造成了膜内电位较大的去极化,而此去极化已不再能被K+外流所抵消,因而能进一步加大膜中Na+通道开放的机率,结果又使更多Na+内流增加而造成膜内进一步的去极化,如此反复促进,就形成一种正反馈的过程,称为再生性循环,其结果使膜内去极化迅速发展,形成动作电位陡峭的升支,直至膜内电位上升到近于Na+平衡电位的水平。由此可见,阈电位不是单一通道的属性,而是在一段膜上能使Na+通道开放的数目足以引起上面描述的再生性循环出现的膜内去极化的临界水平。由此也不难理解,只要刺激大于能引起再生性循环的水平,膜内去极化速度就不再决定于原刺激的大小;整个动作电位上升支的幅度也只决定于原来静息电位的值和膜内外的Na+浓度差,而与引起此次动作电位的刺激大小无关。此即动作电位所以能表现“全或无”现象的机制。
阈电位是用膜本身去极化的临界值来描述动作电位的产生条件。所谓阈强度,是作用于标本时能使膜的静息电位去极化到阈电位的外加刺激的强度;这就是阈强度和阈电位在概念上的区别。
(二)局部兴奋及其特性
一个阈下刺激会对可兴奋细胞产生何种影响?可通过图2-17中的实验回答。在巨大神经轴突放置一对刺激电极,但其中一个电极穿入膜内,再在附近放置一个作膜内电反应记录的记录电极。假定先把膜内的刺激电极连到电源正极,那么电路接通时将会产生去极化;如果这个去极化未能达到阈电位,则说明所用电刺激强度属于阈下刺激。但如前所述,阈下刺激虽未能膜电位达到阈电位的去极化,也能引起该段膜中所含Na+通道的少量开放,只是开放的机率少,于是少量内流的Na+和电刺激造成的去极化叠加起来,在受刺激的膜局部出现一个较小的膜的去极化反应,称为局部反应或局部兴奋,局部兴奋由于强度较弱,且很快被外流的K+所抵消,因而不能引起再生性循环而发展成真正的兴奋或动作电位。图2-17B就记录了一组这样的实验曲线,说明在阈下刺激的范围内,刺激强度愈强,引起的膜的去极化即局部兴奋的幅度愈大(由表示静息电位水平的线段上方的各条曲线表示),延续的时间也愈长;只有当局部兴奋的幅度大到足以引发再生性循环的水平时,膜的去极化的速度才突然加大,这样局部兴奋就发展成为动作电位。
局部兴奋有以下几个基本特性:(1)不是“全或无”的,而是随着阈下刺激的增大而增大;(2)不能在膜上作远距离的传播,虽然由于膜本身有电阻特性且膜内外都是电解质溶液,发生在膜的某一点的局部兴奋,可以使邻近的膜也产生类似的去极化,但随距离加大而迅速减小以至消失,这个局部兴奋所波及的范围在一般神经细胞膜上不超过数十乃至数百微米,但有的细胞本身也不很大,如神经元细胞体,局部兴奋的这种电紧张性扩布(eletrotonic propagation)还是有重要生理意义的;(3)局部兴奋是可以互相叠加的,也就是说,当一处产生的局部兴奋由于电紧张性扩布致使邻近处的膜也出现程度较小的去极化,而该处又因另一刺激也产生了局部兴奋,虽然两者(当然不一定限于两者)单独出现时都不足以引发一次动作电位,但如果遇到一起时可以叠加起来,以致有可能达到阈电位而引发一次动作电位。称为兴奋的空间性总和;局部兴奋的叠加也可以发生在连续受数个阈下刺激的膜的某一点,亦即当前面刺激引起的局部兴奋尚未消失时,与后面刺激引起的局部兴奋发生叠加,称为时间性总和。总和现象在神经元细胞的功能活动中十分重要和常见。另外,由图示2-17B中还可看到,当刺入膜内的刺激电极和电源负极相连时,通电时只能引起膜的超级化(图中水平线下方的那组曲线);刺激愈强,超极化程度愈大,但不引起Na+通道开放,更不能引发锋电位。事实上,这时由于膜内电位和阈电位之间差值加大,因而该处膜变得更不容易兴奋了。体内某些感受器细胞、部分腺细胞和平滑肌细胞,以及神经细胞体上的突触后膜和骨骼肌细胞的终板膜,它们在受刺激时不产生“全或无”形式的动作电位,而只出现原有静息电位的微弱而缓慢的变动,分别称为感受器电位、慢电位、突触后电位和终板电位。这些电位也具有类似局部兴奋的特性。这些形式的电变化,实际是使另一细胞或同一细胞的其他部分的膜产生“全或无”式动作电位上的过渡性电变化。
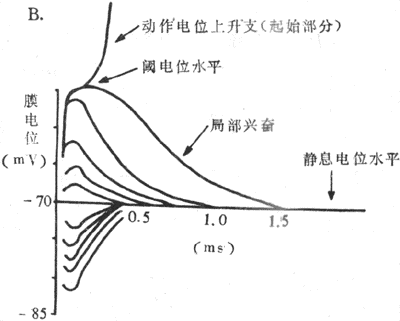
图2-17 局部兴奋的实验布置(A)和实验结果(B)示意图说明见正文
(三)兴奋在同一细胞上的传导机制
可兴奋细胞的特征之一是它任何一处的膜产生的动作电位,都可沿着细胞膜向周围传播,使整个细胞的膜都经历一次类似于被刺激部位的离子电导的改变,表现为动作电位沿整个细胞膜的传导。传导的机制实际已包含在兴奋膜的上述特性之中。设想一条枪乌贼的无髓神经纤维的某一小段,因受到足够强的外加刺激而出现了动作电位(图2-18,B左端),即该处出现了膜两侧电位的暂时性倒转,由静息时的内负外正变为内正外负,但和该段神经相邻接的神经段仍处于安静时的极化状态;由于膜两侧的溶液都是导电的,于是在已兴奋的神经段和与它相邻的未兴奋的神经段之间,将由于电位差的存在而有电荷移动,称为局部电流。它的运动方向是:膜外有正电荷由未兴奋段移向已兴奋段,膜内有正电荷由已兴奋段移向未兴奋段。这样流动的结果,是造成未兴奋段膜内电位升高而膜外电位降低,亦即引起该处膜的去极化;这一过程开始时,就相当于电紧张性扩布。根据上述关于兴奋产生的机制的分析,当任何原因使膜的去极化达到阈电位的水平时,都会大量激活该处的Na+通道而导致动作电位的出现。因此,当局部电流的出现使邻接的未兴奋的膜去极化到阈电位时,也会使该段出现它自己的动作电位。所谓动作电位的传导,实际是已兴奋的膜部分通过局部电流“刺激”了未兴奋的膜部分,使之出现动作电位;这样的过程在膜表面连续进行下去,就表现为兴奋在整个细胞的传导。由于锋电位产生期间电位变化的幅度和陡度相当大,因此在单一细胞局部电流的强度超过了引起邻近膜兴奋所必需的阈强度数倍以上,因而以局部电流为基础的传导过程是相当“安全”的,亦即一般不易因某处动作电位不足以使邻接的膜产生兴奋而导致传导“阻滞”,这一点与一般化学性突触处的兴奋传递有明显的差别。
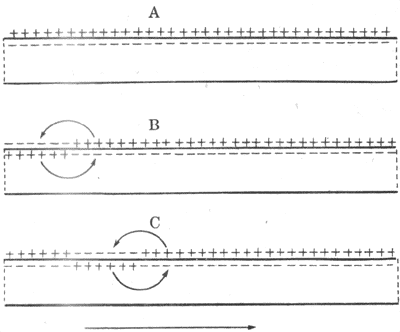
图2-18 神经纤维传导机制的模式图弯箭头表示膜内外
局部电流的流动方向,下方直箭头表示冲动传导方向。
A:静息时 B:发生兴奋后 C:传导过程中
兴奋传导机制虽然以无髓神经纤维为例,但在其他可兴奋细胞(如骨骼肌细胞)的兴奋传导,基本上遵循同样的机制。有髓神经纤维在轴突外面包有一层相当厚的髓鞘,髓鞘主要成分的脂质是不导电或不允许带电离子通过的,因此只有在髓鞘暂时中断的朗飞结处,轴突膜才能和细胞外液接触,使跨膜离子移动得以进行。因此,当有髓纤维受到外加刺激时,动作电位只能在邻近刺激点的朗飞结处产生,而局部电流也只能发生在相邻的朗飞结之间,其外电路要通过髓鞘外面的组织间液,因此,动作电位表现为跨过每一段髓鞘而在相邻朗飞结处相继出现,这称为兴奋的跳跃式传导。
跳跃式传导时的兴奋传导速度,显然比上述无髓纤维或一般细胞的传导速度快得多;而且由于跳跃式传导时,单位长度内每传导一次兴奋所涉及的跨膜离子运动的总数要少得多,因此它还是一种“节能”的传导方式。看来,神经髓鞘的出现是进化过程中既能增加神经纤维传导速度、又能减少生物能量消耗的一种方式。无脊椎动物没有有髓神经纤维,而无髓纤维增加增加传导速度的一个可能途径是增大轴突的直径,因为这样可以减少膜内液体的电阻而增加局部电流的强度,使动作电位的传导速度加快;这大概就是需要进行快速神经反应的枪乌贼在进化中出现巨大的无髓神经纤维的道理所在。但徐科(1993)等人指出,某些无脊椎动物的神经纤维也可以一种特殊的方式进行跳跃式传导。
如果一条神经纤维在它的中间部受到刺激,将会有动作电位由中间向纤维两端传送,这是由于局部电流可以出现在原兴奋段两侧之故。由此可以理解,兴奋在同一细胞上的传导,并不限于朝向某一方向;体内神经纤维所以有传入和传出之分,只是由于在整体的自然条件下,传入纤维只能在它们和感受器相连接的外周端被刺激,而传出纤维只能在它们的细胞体产生冲动而传向外周,并非是由于这些纤维本身只能单方向传导兴奋的缘故。以动作电位为兴奋出现的指标,可以测定兴奋在各种细胞的传导速度。例如,人体一些较粗的有髓神经纤维的传导速度,最快可达每秒100m以上,而一些细胞的无髓纤维每秒传导距离还不到1m;构成心脏内部传导系统的浦肯野细胞,每秒传导速度约4~5m,是心肌细胞中传导速度最快的。
第四节 肌细胞的收缩功能
人体各种形式的运动,主要是靠一些肌细胞的收缩活动来完成的。例如,躯体的各种运动和呼吸动作由骨骼肌的收缩来完成;心脏的射血活动由心肌的收缩来完成;一些中空器官如胃肠、膀胱、子宫、血管等器官的运动,则由平滑肌的收缩来完成。不同肌肉组织在功能和结构上各有特点,但从分子水平来看,各种收缩活动都与细胞内所含的收缩蛋白质,主要与肌凝蛋白和肌纤蛋白的相互作用有关;收缩和舒张过程的控制,也有某些相似之处。本节以研究最充分的骨骼肌为重点,说明肌细胞的收缩机制。
骨骼肌是体内最多的组织,约占体重的40%。在骨和关节的配合下,通过骨骼肌的收缩和舒张,完成人和高等动物的各种躯体运动。骨骼肌由大量成束的肌纤维组成,每条肌纤维就是一个肌细胞。成人肌纤维呈细长圆柱形,直径约60μm,长可达数毫米乃至数十厘米。在大多数肌肉中,肌束和肌纤维都呈平行排列,它们两端都和由结缔组织构成的腱相融合,后者附着在骨上,通常四肢的骨骼肌在附着点之间至少要跨过一个关节,通过肌肉的收缩和舒张,就可能引起肢体的屈曲和伸直。我们的生产劳动、各种体力活动等,都是许多骨骼肌相互配合的活动的结果。每个骨骼肌纤维都是一个独立的功能和结构单位,它们至少接受一个运动神经末梢的支配,并且在体骨骼肌纤维只有在支配它们的神经纤维有神经冲动传来时,才能进行收缩。因此,人体所有的骨骼肌活动,是在中枢神经系统的控制下完成的。
一、神经-骨骼肌接头处的兴奋传递
运动神经纤维在到达神经末梢处时先失去髓鞘,以裸露的轴突末梢嵌入到肌细胞膜上称作终板的膜凹陷中,但轴突末梢的膜和终板膜并不直接接触,而是被充满了细胞外液的接头间隙隔开,其中尚含有成分不明的基质;有时神经末梢下方的终板膜还有规则地再向细胞内凹入,形成许多皱褶,其意义可能在于增加接头后膜的面积,使它可以容纳较多数目的蛋白质分子,它们最初被称为N-型乙酰胆碱受体,现已证明它们是一些化学门控通道,具有能与ACh特异性结合的亚单位。在轴突末梢的轴浆中,除了有许多线粒体外还含有大量直径约50nm的无特殊构造的囊泡(图2-19)。用组织化学的方法可以证明,囊泡内含有ACh;此ACh首先在轴浆中合成,然后贮存在囊泡内。据测定,每个囊泡中贮存的ACh量通常是相当恒定的,且当它们被释放时,也是通过出胞作用,以囊泡为单位“倾囊”释放,被称为量子式释放。在神经末梢处于安静状态时,一般只有少数囊泡随机地进行释放,不能对肌细胞产生显著影响。但当神经末梢处有神经冲动传来时,在动作电位造成的局部膜去极化的影响下,大量囊泡向轴突膜的内侧面靠近,通过囊泡膜与轴突膜的融合,并在融合处出现裂口,使囊泡中的ACh全部进入接头间隙。据推算,一次动作电位的到达,能使大约200~300个囊泡的内容排放,使近107个ACh分子被释放。轴突末梢处的电位变化引起囊泡排放的过程十分复杂,但首先是轴突末梢膜的去极化,引起了该处特有的电压门控式Ca2+通道开放,引起细胞间隙液中的Ca2+进入轴突末梢,触发了囊泡移动以至排放的过程。Ca2+的进入量似乎决定着囊泡释放的数目;细胞外液中低Ca2+或(和)高Mg2+,都可阻碍ACh的释放而影响神经-肌接头的正常功能。已故冯德培院士在30年代对神经-肌接头的化学性质传递进行过重要的研究。
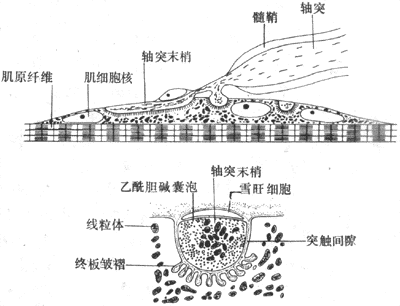
图2-19 神经-肌接头处的超微结构示意图
当ACh分子通过接头间隙到达终板膜表面时,立即同集中存在于该处的特殊通道蛋白质的两个α-亚单位结合,每分子的通道将结合两个分子的ACh,由此引起的蛋白质分子内部构象的变化会导致它的通道结构的开放。这种通道开放时,孔道的横截面比前面提到的Na+通道的面积为大,可允许Na+、K+甚至少量Ca2+同时通过;由于这几种离子正常时在膜内处的分布特点,实际出现的是Na+的内流和K+的外流,其总的结果是使终板膜处原有静息电位减小,向零值靠近,亦即出现膜的去极化;这一电变化,称为终板电位,它的出现约较神经冲动到达接头前膜处晚0.5~1.0ms。有人曾在运动神经无冲动到达末梢时,记录到由于个别囊泡的自发释放在终板膜上引起的微小的电变化,称为微终板电位。终板电位与前述的局部兴奋电反应有类似的性质:不表现“全或无”特性,其大小与接头前膜释放的ACh的量成比例;无不应期,可表现总和现象等,如我们一般记录到的终板电位就是多数微终板电位总和的结果。终板电位产生时,它将以电紧张性扩布的形式影响终板膜周围一般肌细胞膜。一般的肌细胞膜与神经轴突的膜性质类似,其中主要含电压门控式Na+通道和K+通道;因而当同终板膜邻接的肌细胞膜的静息电位由于终板电位的影响而去极化到该处膜的阈电位水平时,就会引发一次向整个肌细胞膜作“全或无”式传导的动作电位,后者再通过所谓“兴奋-收缩耦联”,引起肌细胞出现一次机械收缩。
正常情况下,一次神经冲动所释放的ACh以及它所引起的终板电位的大小,大约超过引起肌细胞膜动作电位所需阈值的3~4倍,因此神经肌接头处的兴奋传递通常是1对1的,亦即运动纤维每有一次神经冲动到达末梢,都能“可靠地”使肌细胞兴奋一次,诱发一次收缩;这一点与将来要讲的神经元之间的兴奋传递有明显不同(见第十章)。接头传递能保持1对1的关系,还要靠每一次神经冲动所释放的ACh能够在它引起一次肌肉兴奋后被迅速清除,否则它将持续作用于终板而使终板膜持续去极化,并影响下次到来的神经冲动的效应。已知,ACh的清除主要靠胆硷酯酶的降解作用来完成,此酶主要分布在接头间隙中和接头后膜上,它们大约可以在2.0ms的时间内将一次神经冲动所释放的ACh清除掉。许多药物可以作用于接头传递过程中的不同阶段,影响正常的接头功能。例如,美洲箭毒和α-银环蛇毒可以同ACh竞争终板膜的ACh受体亚单位,因而可以阻断接头传递而使肌肉失去收缩能力;有类似作用的药物称为肌肉松弛剂;有机磷农药和新斯的明对胆碱酯酶有选择性的抑制作用,可造成ACh在接头和其他部位的大量积聚,引起种种中毒症状。重症肌无力是由于体内骨骼肌终板处的ACh门控通道数量不足或功能障碍所引起。
二、骨骼肌细胞的微细结构
骨骼肌细胞在结构上最突出之点,是含有大量的肌原纤维和丰富的肌管系统,且其排列高度规则有序。肌细胞是体内耗能作功,完成机体多种机械运动的功能单位。
(一)肌原纤维和肌小节
每个肌纤维含有大量直径1~2μm的纤维状结构,称为肌原纤维,它们平行排列,纵贯肌纤维全长,在一个细胞中可达上千条之多(图2-20)。每条肌原纤维的全长都呈现规则的明、暗交替,分别称为明带和暗带;而且在平行的各肌原纤维之间,明带和暗带又都分布在同一水平上;暗带的长度比较固定,不论肌肉处于静止、受到被动牵拉或进行收缩时,它都保持1.5μm的长度;在暗带中央,有一段相对透明的区域,称为H带,它的长度随肌肉所处状态的不同而有变化;在H带中央亦即整个暗带的中央,又有一条横向的暗线,称为M线。明带的长度是可变的,它在肌肉安静时较长,并且在一定范围内可因肌肉所受的被牵引而变长;但明带在肌肉收缩时可变短。明带中央也有一条横向的暗线,称为z线(或z盘)。目前已经肯定,肌原纤维上每一段位于两条z线之间的区域,是肌肉收缩和舒张的最基本单位,它包含一个位于中间部分的暗带和两侧各1/2的明带,合称为肌小节(sarcomere)。由于明带的长度可变,肌小节的长度在不同情况下可变动于1.5~3.5μm之间;通常在体骨骼肌安静时肌小节的长度约为2.0~2.2μm。
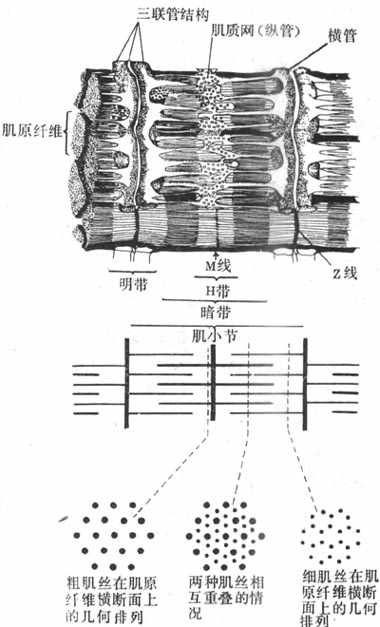
图2-20 骨骼肌细胞的肌原纤维和肌管系统
注意肌小节能的组成和不同部位肌小节横断布粗、细肌丝的几何排列
用X-线衍射等更精密的方法进一步发现,肌小节的明带和暗带包含有更细的、平行排列的丝状结构,称为肌丝。暗带中含有的肌丝较粗,直径约10nm,称为粗肌丝,其长度与暗带相同;实际上暗带的形成就是由于粗肌丝的存在,M线则是把成束的粗肌丝固定在一定位置的某种结构。明带中的肌丝较细,直径约5nm,称为细肌丝;它们由z线结构向两侧明带伸出,每侧的长度都是1.0nm,它的游离端在肌小节总长度小于3.5nm的情况下,必然有一段要伸入暗带,和粗肌丝处于交错和重叠的状态;如果由两侧z线伸入暗带的细肌丝未能相遇而隔有一段距离,这就形成了H带。肌肉被拉长时,肌小节长度增大,这时细肌丝由暗带重叠区拉出,使明带长度增大,H带也相应地增大(图2-20,下)。
粗、细肌丝相互重叠时,在空间上也呈现规则排列,这可从肌原纤维的横断面上看出。图2-20下方表示在肌小节的不同位置将肌原纤维横节时,断面上所能看到的两种肌丝的分布情况。在通过明带的横断面上只有细肌丝,它们的所在位置相当于一个正六边形的各顶点;在通过H带的横断面上只有粗肌丝,它们都处于正三边形的各顶点上;而在H带两侧的暗带的横断面上,则可看到粗、细肌丝交错存在的情况,这里,每一条粗肌丝正处在以六条细肌丝为顶点的正六边形的中央,而这就为收缩时粗细肌丝之间的相互作用准备了条件。
(二)肌管系统
肌管系统指包绕在每一条肌原纤维周围的膜性囊管状结构,由来源和功能都不相同的两组独立的管道系统组成。一部分肌管的走行方向和肌原纤维相垂直,称为横管系统或称T管,是由肌细胞的表面膜向内凹入而形成;它们穿行在肌原纤维之间,并在z线水平(有些动物是在暗带和明带衔接处的水平)形成环绕肌原纤维的管道;它们相互交通,管腔通过肌膜凹入处的小孔与细胞外液相通。将标记物加入到细胞的浸浴液中,这些物质可以很快在每一条环绕肌小节的横管系统中出现,但不能进入肌浆和肌浆网中去。肌原纤维周围还有另一组肌管系统,就是肌浆网,它们的走行方向和肌小节平行,称为纵管系统或称为L管;纵管系统或肌浆网主要包绕每个肌小节的中间部分,这是一些相互沟通的管道,但是在接近肌小节两端的横管时管腔出现膨大,称为终末池,它使纵管以较大的面积和横管相靠近。每一横管和来自两侧肌小节的纵管终末池,构成了三联管结构(图2-20,上)。据研究,横管和纵管的膜在三联管结构处并不接触,中间尚隔有约12nm的间隙,说明两组管道的内腔并不直接沟通,但这样的结构显然有利于细胞内外之间某种形式的信息传递。目前普遍承认的看法是,横管系统的作用是将肌细胞兴奋时出现在细胞膜上的电变化沿T管膜传入细胞内部,肌浆网和终末池的作用是通过对钙离子的贮存、释放和再积聚,触发肌小节的收缩和舒张;而三联管结构是把肌细胞膜的电变化和细胞内的收缩过程衔接或耦联起来的关键部位。
三、骨骼肌的收缩机制和兴奋-收缩耦联
Huxley等在50年代初期就提出了用肌小节中粗、细肌丝的相互滑行来说明肌肉收缩的机制。这一被称为滑行理论(sliding theory)的主要内容是:肌肉收缩时虽然在外观上可以看到整个肌肉或肌纤维的缩短,但在肌细胞内并无肌丝或它们所含的分子结构的缩短,而只是在每一个肌小节内发生了细肌丝向粗肌比之间的滑行,亦即由z线发出的细肌丝在某种力量的作用下主动向暗带中央移动,结果各相邻的z线都互相靠近,肌小节长度变短,造成整个肌原纤维、肌细胞乃至整条肌肉长度的缩短。滑行现象最直接的证明是,肌肉收缩时并无暗带长度的变化,而只能看到明带长度的缩短;并且与此同时也看到暗带中央H带相应地变窄。这只能说明,细肌丝在肌肉收缩时也没有缩短,只是它们更向暗带中央移动,和粗肌丝发生了更大程度的重叠。这种变化只能用粗、细肌丝之间出现了相对运动即滑行现象来解释。滑行理论需要进一步说明的问题是:肌肉收缩时究竟是什么力量促使细肌丝向粗肌丝之间滑行,以及怎样把这些过程和肌肉膜的兴奋过程联系起来。近年来,由于肌肉生物化学及其他细胞生物学技术的发展,肌丝滑行的机制已基本上从组成肌丝的蛋白质分子结构的水平得到阐明,对于与滑行的开始和终止有关的各种控制因素,也有了较较深入的了解。
(一)肌丝的分子组成和横桥的运动
滑行现象的引起与组成肌丝的蛋白质分子结构和它们的特性有直接的关系。粗肌丝主要由肌凝蛋白(亦称肌球蛋白)所组成,它们的分子在粗肌丝中呈独特的有规则的排列。一条粗肌丝大约含有200~300个肌凝蛋白分子,每个分子长150nm,呈长杆状而在一端有球状膨大部。在组成粗肌丝时,各杆状部朝向M线而聚合成束,形成粗肌丝的主干,球状部则有规则地裸露在M线两侧的粗肌丝主干的表面,形成横桥(图2-21,左)。当肌肉安静时,横桥与主干的方向相垂直,由粗肌丝表面突出约6nm。用X-线衍射法证明,横桥在粗肌丝表面的分布位置也是严格有规则的,即在粗肌丝的同一周径上只能有两个相隔180°的横桥突出;在与此周径相隔14.3nm的主干上又有一对横桥突出,但与前一对有60°的夹角;如此反复,到第四对横桥出现时,其方向正好与第一对横桥相平行,且与第一对横桥相隔42.9nm(图示-21,右)。上述横桥的分布情况,正好与一条粗肌丝为6条细肌丝所环绕的情况相对应,亦即在所有横桥出现的位置,正好有一条细肌丝与之相对;而对于每条细肌丝来说,粗肌丝表面每隔42.9nm就伸出一个横桥与之相对(图2-21)。这种对应关系,对于粗、细肌丝之间的相互作用显然是十分有利的。
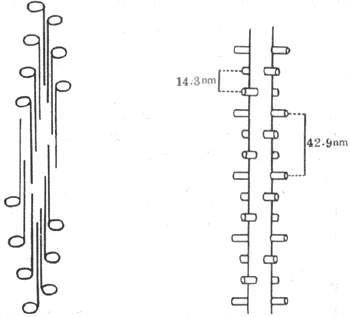
图2-21 粗肌丝中肌凝蛋白分子的排列示意图
左、肌凝蛋白分子的长杆状部横向聚合,形成粗肌丝主干,球状部裸露在表面,形成横桥
右、横桥在粗肌丝表面的几何排列
现已证明,横桥所具有的生物化学特性对于肌丝的滑行有重要意义。横桥的主要特性有二:一是横桥在一定条件下可以和细肌丝上的肌纤蛋白分子呈可逆性的结合,同时出现横桥向M线方向的扭动,继而出现横桥和细肌丝的解离、复位,然后再同细肌丝上另外的点结合,出现新的扭动,如此反复,使细肌丝继续向M线方向移动;二是横桥具有ATP酶的作用,可以分解ATP而获得能量,作为横桥摆动和作功的能量来源。由此可见,横桥和细肌丝的相互作用,是引起肌丝滑行的必要条件。
细肌丝至少由三种蛋白质组成,其中60%是肌纤蛋白(亦称肌动蛋白)。肌纤蛋白与肌丝滑行有直接的关系,故和肌凝蛋白一同被称为收缩蛋白质。肌纤蛋白分子单体呈球状,但它们在细肌丝中聚合成双螺旋状,成为细肌丝的主干(图2-22)。细肌丝中另外有两种蛋白质,它们不直接参与肌丝间的相互作用,但可影响和控制收缩蛋白质之间的相互作用,故称为调节蛋白质;其中一种是原肌凝蛋白,也呈双螺旋结构,在细肌丝中和肌纤蛋白双螺旋并行,但在肌肉安静时原肌凝蛋白的位置正好在肌纤蛋白和横桥之间(图2-23,左)这就起了阻碍两者相互结合的作用;另一种调节蛋白质称为肌钙蛋白(亦称原宁蛋白),肌钙蛋白在细肌丝上不直接和肌纤蛋白分子相连接,而只是以一定的间隔出现在原肌凝蛋白的双螺旋结构之上。肌钙蛋白的分子呈球形,含有三个亚单位(图2-22):亚单位C中有一些带双负电荷的结合位点,因而对肌浆中出现的Ca2+(以及其他可能出现的两价正离子和H+)有很大的亲和力;亚单位T作用是把整个肌钙蛋白分子结合于原肌凝蛋白;而亚单位I的作用是在亚单位C与Ca2+结合时,把信息传递给原肌凝蛋白,引起后者的分子构象发生改变,解除它对肌纤蛋白和横桥相互结合的阻碍作用。
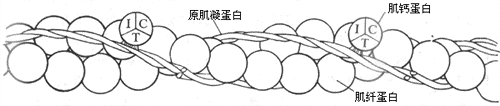
图 2-22 细肌丝的分子组成
I、T、C分别代表肌钙蛋白的三个亚单位
肌丝滑行的基本过程一般公认为:当肌细胞上的动作电位引起肌浆中Ca2+浓度升高时,作为Ca2+受体的肌钙蛋白结合了足够数量的Ca2+,这就引起了肌钙蛋白分子构象的某些改变,这种改变“传递”给了原肌凝蛋白,使后者的构象也发生某些改变,其结果是使原肌凝蛋白的双螺旋结构发生了某种扭转,这就把安静时阻止肌纤蛋白和横桥相互结合的阻碍因素除去,出现了两者的结合。在横桥与肌纤蛋白的结合、扭动、解离和再结合、再扭动构成的横桥循环过程中,使细肌丝不断向暗带中央移动;与此相伴随的是ATP的分解消耗和化学能向机械能的转换,完成了肌肉的收缩(图示2-23)。
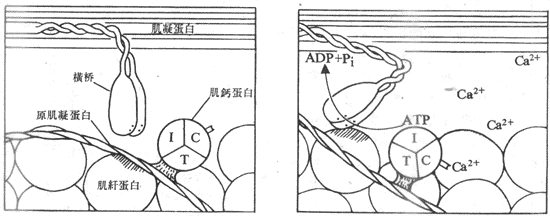
图2-23 Ca2+通过和肌钙蛋白的结合,诱发横桥和肌纤蛋白之间的相互作用
上述的横桥循环在一个肌小节以至整个肌肉中都是非同步地进行的,这样才可能使肌肉产生恒定的张力和连续的缩短。至于能参与循环的横桥数目以及横桥循环的进行速率,则是决定肌肉缩短程度、缩短速度以及所产生张力的关键因素,这将在分析肌肉收缩的力学表现时再作讨论。下面叙述正常条件下引发和终止横桥循环的肌浆中Ca2+浓度的突然升高和快速下降是怎样发生的。
(二)骨骼肌的兴奋-收缩耦联
在整体情况下,骨骼肌总是在支配它的躯体传出神经的兴奋冲动的影响下进行收缩的;直接用人工刺激作用无神经支配的骨骼肌,也可引起收缩。但不论何种情况,刺激在引起收缩之前,都是先在肌细胞膜上引起一个可传导的动作电位,然后才出现肌细胞的收缩反应。这样,在以膜的电变化为特征的兴奋过程和以肌丝的滑行为基础的收缩过程之间,必然存在着某种中介性过程把两者联系起来,这一过程,称为兴奋-收缩耦联。目前认为,它至少包括三个主要步骤:电兴奋通过横管系统传向肌细胞的深处;三联管结构处的信息传递;肌浆网(即纵管系统)对Ca2+释放和再聚积。
横管系统对正常肌细胞的兴奋-收缩耦联是十分必要的。用含有甘油的高渗任氏液浸泡肌肉一段时间,再把它放回到一般任氏液中,这样的处理可以选择性地破坏肌细胞的横管系统;这时如果再给肌肉以外加刺激,虽然仍可在完好的肌细胞膜上引起动作电位,但不再能引起细胞收缩。近年来证明,横管膜和一般肌细胞膜有类似的特性,又是后者的延续部分,因而它也可以产生以Na+内流为基础的膜的去极化甚或动作电位;当一般细胞膜因兴奋而产生动作电位时,这一电变化可沿着凹入细胞内部的横管膜传导,深入到三联管结构和每个肌小节的近旁。
实际测定还证明,肌肉安静时肌浆中的Ca2+浓度低于10-7mol/L,但在膜开始去极化的很短时间内,可以在1~5ms内升高到10-5mol/L的水平,亦即增高100倍之多。这样多的Ca2+由何而来?用放射性45Ca自显影等技术证明,肌肉安静时Ca2+主要停留和聚积在z线附近,相当于肌浆网的终末池部位;肌肉收缩时,Ca2+由这里向暗带区扩散,触发横桥循环。这样问题就归结为:当肌膜上的电变化沿横管系统到达三联管部分时,一定有某种因子把横管膜上发生的变化传递给了相距不远的肌浆网膜上的类似Ca2+通道的结构,引起后者分子的变构作用,使通道开放,于是肌浆网内高浓度的Ca2+就不需耗能而靠易化扩散进入肌浆,到达肌丝区。传递这一信号的因子,有人认为是横管膜上存在的一种特殊蛋白,平时对肌浆网Ca2+通道外侧开口有机械堵塞作用,但在横管膜有电变化时发生变构作用,使原来的堵塞作用解除;也有人认为横管膜可因电变化而产生了第二信使类物质IP3(见本章第二节),由后者作用于Ca2+通道使之开放。由于三联管外有关的膜和膜中蛋白质几乎可以相互接触,因而第一种控制形式还是有可能的。
释放到肌浆中的Ca2+怎样被迅速除去,目前已证明是由于肌浆网膜结构中存在的一种特殊的离子转运蛋白质即钙泵活动的结果。钙泵是一种Ca2+依赖式ATP酶,目前已被分离提纯,它占肌浆网膜蛋白质总量的60%;在肌浆中Ca2+增高情况上,它可以分解ATP获得能量,将Ca2+在逆浓度差的情况下由肌浆转运到肌浆网内腔中去;由于肌浆中Ca2+浓度的降低,和肌钙蛋白结合的Ca2+也解离,引起肌肉舒张。
四、骨骼肌收缩的外部表现和力学分析
骨骼肌在体内的功能,就是它们在受刺激时能产生缩短或(和)张力,藉以完成躯体的运动或(和)抵抗外力的作用。当肌肉克服某一外力而缩短,或肌肉因缩短而牵动某一负荷时,肌肉就完成了一定量的机械功,其数值等于它所克服的阻力(或负荷)和肌肉缩短长度的乘积;如以缩短速度乘以负荷,则得出肌肉的输出功率。但肌肉在收缩时究竟以产生张力为主或缩短为主,以及收缩时能作多少功,则要看肌肉收缩时所遇到的负荷条件和肌肉本身的功能状态。
肌肉在体内或实验条件下可能遇到的负荷主要有两种:一种是在肌肉收缩前就加在肌肉上的,如把一条肌肉顺着它的肌原纤维的走行方向悬挂起来而把上端固定,再在另一端悬挂一定数量的重物,后者就是前负荷。前负荷使肌肉在收缩前就处于某种程度的被拉长状态,使它具有一定的长度,这称为初长度;这样由于前负荷的不同,同一肌肉就要在不同的初长度条件下进行收缩。另一种负荷称为后负荷,它是在肌肉开始收缩时才能遇到的负荷或阻力,它不增加肌肉的初长度,但能阻碍收缩时肌肉的缩短。可以理解,对于某一具体的肌肉来说,实验中所加负荷、特别是前负荷不应当过大,因为后者在肌肉收缩前就可能因过度的牵拉而损伤肌肉本身的结构;至于后负荷,它在大到一定程度就足以抵抗肌肉收缩所产生的最大张力,因而肌肉不再表现缩短,出现等长收缩,亦即这时肌肉虽进行了收缩,并未有长度改变;在这种情况下继续增加后负荷,显然不会对肌肉的收缩有什么影响。
据上述,能影响肌肉收缩时作功能力或其力学表现的因素至少有三个,即前负荷、后负荷和肌肉本身的功能状态(即肌肉收缩能力)。要分析某一因素影响的最简单办法,就是使其他因素保持在某一恒定值而改变要观察因素的值,得到一组数据并作成一条座标曲线来进行分析。
(一)前负荷或肌肉初长度对肌肉收缩的影响枣长度-张力曲线
为了保持在实验过程中肌肉本身的功能状态基本保持不变,通常选用代谢速度较慢的两栖类如蛙腓肠或缝匠进行实验,实验布置如图2-24A所示。肌肉在下方被固定,并且边了一个灵敏的张力换能器来记录肌肉收缩前和收缩后的张力产生情况;肌肉的上方连一个可移动的按钮,可以上下移动而改变肌肉的初长度,但不论初长度固定在什么长度,同旋钮相连的固定杆是不能动的,这就意味着把后负荷固定在无限大时的位置,肌肉在收缩时不可能缩短而只能产生张力(即前面所说的等长收缩),于是就可以观察初长度不同时对同一肌肉所能产生张力的影响了。
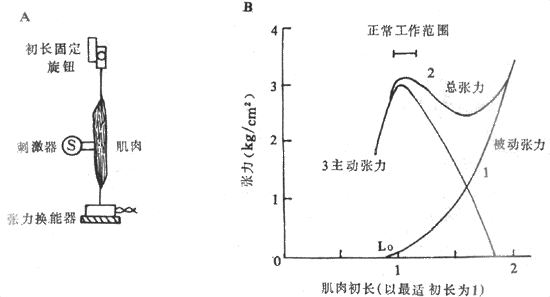
图2-24 肌肉初长度对肌肉收缩的影响
A:在实验布置中,下方是张力换能器,它位置固定,能把所受张力转变为相应的
电信号,上方旋钮可将肌肉初长在给肌肉刺激前固定于预定长度
B:3条曲线分别代表肌肉在初长度改变时的张力改变情况,被动张力指改变初长而尚未收缩的
肌肉的张力改变,总张力是在已有被动张力的基础上肌肉收缩时产生的主动张力与前者之和
图2-24B的长度-张力曲线反映了在依次改变肌肉的初长度时(横座标)在张力换能器上记录到的肌肉的张力产生的情况(纵座标)。曲线1是只改变肌肉初长度并不刺激肌肉收缩时肌肉所受的拉力,称为被动张力曲线,它反映安静肌肉具有某种弹性,在受到牵拉时产生某种回弹力,但牵拉超过某种程度,达到了弹性限度,被动张力急速增大,有可能造成组织损坏,其过程有如拉长一个弹簧时类似。曲线2是肌肉在具有不同前负荷即已具有被动张力的条件下进行一次收缩时记录到的张力变化,曲线的每一点都代表那个初长度时肌肉已有的被动张力和收缩时新产生的张力之和,故整个曲线称为总张力曲线;因此,由曲线2代表的不同初长度时的总张力减去同一初长度时的被动张力,就能得到曲线3,它表示肌肉在不同前负荷时进行收缩所能产生的张力,故称为主动张力曲线,它反映了本实验中要观察的内容,即不同前负荷或初长度对肌肉收缩所能产生的张力影响:当前负荷开始增加时,每次收缩所产生的主动张力也相应地增大,但在超过某一限度后,再增加前负荷反而使主动张力越来越小,以致于为零,如曲线3右端所示。这个结论也可以表达为,对于肌肉在等长度收缩条件下所产生的主动张力大小,存在着一个最适前负荷和与之相对应的最适初长度,相当于图2-24B横座标上Lo的位置,在这样的初长度情况下进行收缩,产生的张力最大。
肌肉在最适初长度条件下进行收缩何以能产生最大的张力,很容易根据肌肉被前负荷拉长时对每一肌小节中粗、细肌丝相互关系的改变来解释。已知,肌肉产生张力和缩短,靠的是粗肌丝表面的横桥和细肌丝之间的相互作用;肌肉初长度的大小,决定着每个肌小节的长度,亦即细肌丝和粗肌丝重叠的程度,而后者又决定于肌肉收缩时有多少横桥可以与附近的细肌丝相互作用。从理论上分析,粗肌丝的长度是1.5μm,但在M线两侧各为0.1μm的范围内正常时没有横桥,因此在M线两侧有横桥的粗肌丝长度各为0.65μm,这样当每侧细肌丝伸入暗带0.65μm(尚未于明带的细肌丝长度为0.35μm),亦即肌小节总长度为2.2μm时,粗肌丝上的每个横桥都能与细肌丝作用,因而收缩时能出现最佳的效果。当肌肉处于最适前负荷或最适初长度时,每个肌小节的长度正是2.2μm,如图2-25箭头3所示。如果稍稍减少前负荷使肌小节长度2.0μm(箭头2),尽管每侧细肌丝又多伸入暗带0.1μm(这时两侧细肌丝正好相遇),但这一段正是粗肌丝上无横桥伸出的部分,因而肌肉收缩时起作用的横桥数目并未增多(相当于图2-25中的箭头2)。至于再减小肌小节的长度,则细肌丝可能穿过M线或两侧肌丝相互重合和卷屈,因而造成收缩张力下降(图中箭头1)。反之,如果前负荷超过最适前负荷,收缩前肌小节的长度将大于2.2μm,细肌丝和粗肌丝相互重合的程度逐渐变小,使得肌肉收缩时起作用的横桥数也减少,造成所产生张力的下降;当前负荷使肌小节长度增加到3.5μm时,细肌丝将全部由暗带拉出,这时肌肉受刺激时不再产生主动张力(图中箭头4)。由此可见,通过前负荷对肌小节中粗、细肌丝重合程度的影响,可以说明骨骼肌长度-张力曲线的特点。
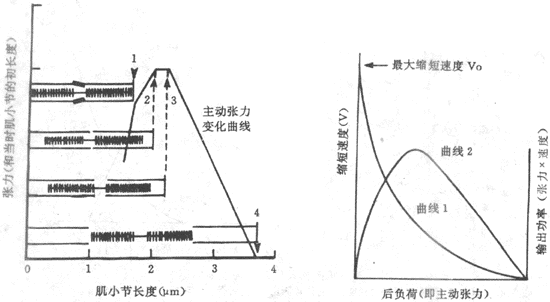
图2-25 不同初长度时粗、细肌丝重合程度和产生张力的关系示意图
用肌小节在不同前负荷时粗、细肌丝相对位置的改变,来说明不同前负荷时所产生的主动张力的不同:在箭头1所指的初长度时,每个肌小节中两侧细肌丝伸入暗带过多,互相重叠或发生卷屈,不利于与横桥间的相互作用;在箭头2和3所指的情况下,肌小节中全部横桥都可与细肌丝相互作用,产生出最大主动张力;在箭头4的情况下,细肌丝相互全部由暗带被拉出,失去产生张力的条件
图2-26 肌肉的张力-速度关系曲线
在肌肉前负荷固定在适当值的条件下,改变后负荷对肌肉产生张力(横座标)
和速短速度(左侧纵座标)相互关系的影响,这称为张力-速度关系曲线(曲线1),
由此曲线可以算出不同后负荷时的输出功率(右侧纵座标),组成了曲线2
(二)肌肉后负荷对肌肉收缩的影响-张力-速度曲线
据前述,如在实验室装置的设计中使一条骨骼肌的前负荷固定不变而可以人为地改变后负荷,即可观察不同后负荷对肌肉收缩的影响。一般情况下,可以把肌肉的前负荷固定在它的最适前负荷(这时出现的被动张力极小(见图2-24B),然后,在逐次改变后负荷的情况下观察肌肉收缩时的情况。不论在任何前负荷的情况下,如果所加后负荷超过了肌肉收缩时所能产生的最大张力(注意负荷的重量值和肌肉产生的张力的值可以用相同的物理单位度量,而且有相同的值),那么肌肉收缩时将只产生张力而不出现肌肉长度的改变。因此,在改变后负荷的实验中所加的后负荷都应小于这个最大张力,那么肌肉在收缩时产生的主动张力超过这个后负荷的值时,它将会出现一定程度的长度缩短,使移动酌相同的距离,并且由于后者也可以算出一个缩短速度来(可以是初速度或平均速度)。后负荷愈小,肌肉产生的张力将较早地超过这个负荷,并且出现较大的缩短长度和缩短速度,但相当于负荷值的肌肉张力却在缩短的过程中保持不变。这样就得到了改变后负荷时,肌肉产生张力和其缩短速度变化的关系曲线(为了计算输出功率,一般只分析缩短速度和张力的关系),即图2-26曲线1,称为张力-速度曲线。该曲线类似一条双曲线,横座标表示肌肉所产生的张力,纵座标表示收缩速度,双曲线的性质则说明这二者大致呈反比的关系,即后负荷减小时,使肌肉产生的张力减小,但可得到一个较大的缩短速度;在曲线同纵轴相交的点,说明后负荷理论上为零时,可以得到该肌肉在当时的功能状态下的最大收缩速度,在图2-26中用V表示;但这时因无负荷,肌肉并未作功,亦无功率输出。在曲线同横轴相交的点,后负荷的值相当于肌肉所能产生的最大张力,这时不能移动负荷,也没有作功和功率输出;在这两个极端之间,在不同的后负荷时都能看到肌肉在产生与负荷相同的张力的情况下使负荷移动一定距离,这种类型的收缩,称为待张收缩,都可作功和有功率输出,但以后负荷相当于最大张力的30%左右时,肌肉的输出功率最大,如图2-26中曲线2所示。
(三)肌肉收缩能力的改变对肌肉收缩的影响
上述的前、后负荷的改变对肌肉收缩时张力产生、缩短速度以及作功能力等力学表现的影响,显然是在肌肉功能状态恒定的情况下对所处负荷条件改变所作的不同反应。但肌肉的状态也是可以改变的,它也可以影响肌肉收缩的效率。例如,缺氧、酸中毒、肌肉中能源物质缺乏,以及其他原因引起的兴奋-收缩耦联、肌肉蛋白质或横桥功能特性的改变,都可能降低肌肉收缩的效果,而钙离子、咖啡因、肾上腺素等体液因素则可能通过影响肌肉的收缩机制而提高肌肉的收缩效果。将影响肌肉收缩效果的肌肉内部功能状态的改变,定义为肌肉收缩能力(contractility)的改变,以区别于肌肉收缩时外部条件即前、后负荷改变所导致的收缩效果的改变。这样的区分虽然在概念上比较容易,但在具体情况下要区分哪些改变是由于肌肉收缩能力的改变所引起,哪些是由于负荷条件的改变所引起,常常十分困难。例如,一个肌肉的最大张力变大了,可能是由于肌肉收缩能力的提高,但也可能是由于在这次收缩前它处于最适初长度;一个肌肉等张收缩时的收缩速度增大了,可能是由于后负荷的减小,也可能是它处于最适初长,但也可能是由于肌肉收缩能力的提高,或三者兼而有之。这就是说,很难简单地根据肌肉某项力学指标的改变,确定是否发生了肌肉收缩能力的改变。从理论上讲,肌肉收缩能力的改变对肌肉收缩的各力学表现的影响是“非选择性”的,即收缩能力的提高可使图2-24B中的长度-张力曲线(主动张力)和图2-26中的张力-速度曲线的位置都升高。显然,为了检查收缩能力是否改变而再绘制一条条座标曲线是十分复杂的;为了简便,如果能让同一肌肉所处的前、后负荷条件不变而发现有肌肉收缩速度的改变,或使肌肉维持最适初长度而有最大张力的改变,则都表示肌肉收缩能力发生了改变;因为在这些条件下可以肯定,这些收缩效果的改变并不是由于前、后负荷的改变所引起,因而只能是由于肌肉的内在性能的改变引起的。
(四)肌肉的单收缩和单收缩的复合
整块骨骼肌或单个肌细胞受到一次短促的刺激时,先是产生一次动作电位,紧接着出现一次机械收缩,后者称为单收缩;根据收缩时肌肉所处的负荷条件不同,单收缩可以是等长的,也可以是等张的。前面叙述的肌肉收缩时各种力学表现,就是以单收缩为观察对象而进行分析的。但在正常体内,当骨骼肌在运动神经的支配下进行自然收缩时,几乎是无例外地接受来自神经的连续刺激,因此有必要进一步分析肌肉有受到不同频率的连续刺激时可能发生的情况。为了便于分析,先观察一下肌肉单收缩时电变化和机械变化在时间上的关系。图2-27是猫胫前有一次等长单收缩时张力变化的全过程,同时记录了肌肉的动作电位。注意图中电反应的开始要较张力增加的开始为早,而且电变化在张力达到顶点以前早已结束;以张力最高点为界,收缩全过程可分为收缩期和舒张期,前者持续时间较后者为短。整个单收缩的时间因肌肉不同而有显著差异,如人的眼外肌的一次单收缩不超过10ms,而腓肠肌可达100ms以上。
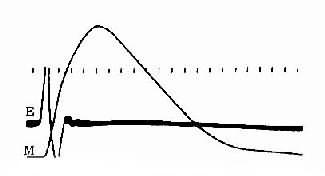
图2-27 猫胫前肌的等长单收缩曲线
M:肌肉收缩时的张力变化曲线
E:肌肉的双相动作电位记录时标每 格相当于0.01s
如果给肌肉以连续的脉刺激,肌肉的收缩情况将随刺激的频率而有不同。如图2-28所示,在刺激的频率较低时,因每一个新的刺激到来时由前一次刺激引起的单收缩过程(包括舒张期)已经结束,于是每次刺激都引起一次独立的单收缩;当刺激频率增加到某一限度时,后来的刺激有可能在前一次收缩的舒张期结束前即到达肌肉,于是肌肉在自身尚处于一定程度的缩短或张力存在的基础上进行新的收缩,发生了所谓收缩过程的复合,这样连续进行下去,肌肉就表现为不完全强直收缩,其特点是每次新的收缩都出现在前次收缩的舒张期过程中,在描记曲线上形成锯齿形;如果刺激频率继续增加,那么肌肉就有可能在前一次收缩的收缩期结束以前或在收缩期的顶点开始新的收缩,于是各次收缩的张力或长度变化可以融合而叠加起来,使描记曲线上的锯齿形消失,这就是完全强直收缩。
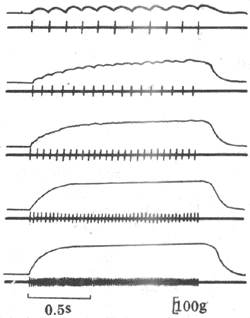
图2-28 不同频率的连续刺激对骨骼肌收缩的影响
每组曲线中,上方曲线是肌肉等长收缩的记录,下方曲线是肌肉动作电位的记录,
注意由上而下当刺激频率依次增加时,机械收缩可出现逐渐融合,
表现不完全和完全强直收缩,但动作电位始终彼此分离,不发生融合和叠加
由于正常体内由运动神经传到骨骼肌的兴奋冲动都是快速连续的,体内骨骼肌收缩几乎都属于完全强直收缩,只不过强直收缩的持续时间可长可短。强直收缩显然可以产生更大的收缩效果,例如,强直收缩所能产生的最大张力可达单收缩的4倍左右。这是因为肌肉在只接受一次刺激时,释放到肌浆中的Ca2+很快被肌浆网上的Ca2+泵回收入肌浆网,而连续刺激可使肌浆中的Ca2+维持在一个饱和的高浓度水平。不同肌肉单收缩的持续时间不同,因而能引起肌肉出现完全强直收缩的最低临界频率在不同肌肉也不同,例如,单收缩快速的眼球内直肌需要每秒约350次的高频刺激才能产生完全强直收缩,而收缩缓慢的比目鱼肌只需每秒约30次的频率就够了。但不论在不完全强直收缩或完全强直收缩,伴随每次刺激出现的肌肉动作电位只出现频率加快,却始终各自分离而不会发生融合或总和;这是由于肌肉的动作电位只持续1~2ms,当刺激频率加速到下一次刺激落于前一次刺激引进起的动作电位持续期间时,组织又正好处于兴奋的绝对不应期,这时新的刺激将无效,既不能引起新的动作电位产生,也不引起新的收缩。
五、平滑肌的结构和生理特性
平滑肌广泛分布于人体消化道、呼吸道以及血管和泌尿、生殖等系统;它和骨骼肌不同,不是每条肌纤维(即肌细胞)的两端都通过肌腱同骨骼相连;平滑肌细胞互相连接,形成管状结构或中空器官;在功能上可以通过缩短和产生张力使器官发生运动和变形,也可产生连续收缩或紧张性收缩,使器官对抗所加负荷而保持原有的形状,前者如胃和肠,后者如动脉血管、括约肌等。此外,也不能像在骨骼肌和心肌那样,把分布在不同器官的平滑肌看作具有相同功能特性和调节机制的组织,例如有些器官的平滑具有和心脏一样的自动节律性,有些则像骨骼肌那样,只有在支配它的神经纤维有神经冲动到来时才出现收缩,而在这两个极端之间,还存在着各种的过渡形式,致使平滑肌的分类困难。
(一)平滑肌的微细结构和收缩机制
平滑肌虽然也具有同骨骼肌类似的肌丝结构,但由于它们不存在像骨骼肌那样平行而有序的排列(平滑肌的肌丝有它自己的“有序的”排列),图2-29是根据最近观察提出的平滑肌细胞内部假想结构图,它的特点是细胞内部存在一个细胞骨架,包含一些卵圆形的称为致密体的结构,它们也间隔地出现于细胞膜的内侧,称为致密区,并且后者与相邻细胞的类似结构相对,而且两层细胞膜也在此处连结甚紧,因而共同组成了一种机械性耦联,藉以完成细胞间张力的传递;细胞间也存在别的连接形式,如缝隙连接,它们可以实现细胞间的电耦联和化学耦联。在致密体和致密区中发现有同骨骼肌Z带中类似的蛋白成分,故认为这两种结构可能是与细肌丝连接的部位。另外,在致密体和致密区之间还有一种直径介于粗、细肌丝之间的丝状物存在,它们是一种称为结蛋白(desmin)的聚合体。这样由丝状物联结起来的致密体和膜内侧的致密区就形成了完整的细胞内构架。
平滑肌细胞中的细肌丝有同骨骼肌类似的分子结构,但不含肌钙蛋白;同一体积的平滑肌所含肌纤蛋白的量是骨骼肌的2倍,推测平滑肌肌浆中有大量细肌丝存在,它们的排列大致与细胞长轴平行。与此相反,胞浆中肌凝蛋白的量却只有骨骼肌的1/4。估计连接在致密体上的3~5根细肌丝会被较少数目粗肌丝包绕,形成相互交错式的排列,这可能就是类似于骨骼肌中肌小节的功能单位。
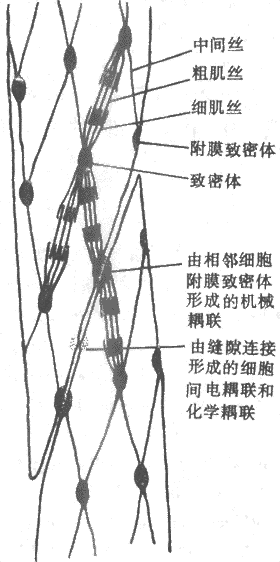
图2-29平骨肌内部结构和相互关系模式图
一般平滑肌细胞呈梭形,直径2~5μm;其长度可变性很大,大约长度为400μm时是产生张力的最适长度。它们没有骨骼肌(和心肌)那样发达的肌管系统。肌细胞膜只有一些纵向排列的袋状凹入,但其功能尚不清楚,不过这使得细胞膜表面积和细胞体积之比更为加大,因此和肌丝靠近的不是横管或肌浆网系统,而是肌膜。细胞被激活时,细胞外Ca2+进入膜内,但平滑肌细胞中靠近膜的肌浆网也构成了细胞内Ca2+贮存库。一些兴奋性递质、激素或药物同肌膜受体结合时,通过G-蛋白在胞浆中产生第二信使,引起Ca2+库中的Ca2+释出。因平滑肌的细肌丝中不存在肌钙蛋白,因而Ca2+引起平滑肌细胞中粗、细肌丝相互滑行的横桥循环的机制与骨骼肌不同。目前认为,横桥的激活开始于它的磷酸化,而这又依赖一称为肌凝蛋白激酶的活化;其过程是Ca2+先结合于胞浆中一种称为钙调蛋白(calmodulin)的特殊蛋白质,后者结合4个Ca2+之后才使肌凝蛋白激酶活化,使ATP分解,由此产生的磷酸基结合于横桥并使横桥处于高自由状态。比起平滑肌来,平滑肌横桥激活的机制需要较长的时间,这和平滑肌收缩的缓慢相一致。
(二)平滑肌在功能上的分类
尽管体内各器官所含平滑肌在功能特性上判别很大,但一般可分为两大类:一类称为多单位(multi-unit)平滑肌,其中所含各平滑肌细胞在活动时各自独立,类似骨骼肌细胞,如竖毛肌、虹膜肌、瞬膜肌(猫)、以及大血管平滑肌等,它们各细胞的活动受外来神经支配或受扩散到各细胞的激素的影响;另一类称为单位(single-unit)平滑肌,类似心肌组织,其中各细胞通过细胞间的电耦联而可以进行同步性活动,这类平滑肌大都具有自律性,在没有外来神经支配时也可进行近于正常的收缩活动(由于起搏细胞的自律性和内在神经丛的作用),以胃肠、子宫、输尿管平滑肌为代表。还有一些平滑肌兼有两方面的特点,很难归入哪一类,如小动脉和小静脉平滑肌一般认为属于多单位平滑肌,但又有自律性;膀胱平滑肌没有自律性,但在遇到牵拉时可作为一个整体起反应,故也列入单位平滑肌。
(三)平滑肌活动的控制和调节
和平滑肌本身的特性具有多样性一样,它们活动所受的调控也是多种多样的,不象骨骼肌那样单纯。大多数平滑肌接受神经支配,包括来自自主神经系统的外来神经支配,其中除小动脉一般只接受交感系统一种外来神经支配外,其他器官的平滑肌通常接受交感和副交感两种神经支配。平滑肌组织、特别是消化管平滑肌肌层中还有内在神经丛存在,后者接受外来神经的影响,但其中还发现有局部传入性神经元,可以引起各种反射。平滑肌的神经-肌接头有些类似骨骼肌,但不具有后者那样特殊结构形式。支配平滑肌的外来神经纤维在进入靶组织时多次分支,分支上每隔一定距离出现一个膨大,呈念珠状,称为曲张体,其中含有分泌囊泡,它们在神经冲动到达时可以释放其中递质或其他神经活性物质;每个曲张体和靶细胞的距离亦不固定,平均约为80~100nm,这说明由神经末梢释放出来的递质分子要扩散较远距离才能达到靶细胞,而靶细胞和神经末梢的关系也不可能是固定的;凡是递质分子可以到达而又具有该递质受体的平滑肌细胞,都可能接受外来神经的影响。
平滑肌细胞约保持-55~-60mV的静息电位,产生机制和骨骼肌类似。单位平滑肌细胞有产生动作电位的能力,而且通过细胞间通道可使相邻细胞也产生动作电位。
参考资料
· 乔健天,关于兴奋膜的一些基本概念和新进展.生理科学进展1980;11:193~200
· 董政,朱培闳.肌醇磷脂信使系统调控骨骼肌兴奋收缩耦联.生理科学进展1994;25:121~125
· BerneRM,Levy MN.Physiology 3rd ed,CV Mosby Co,StLouis,1993
· WrightFM.The intestinal Na/glucose cotransporter .Ann Rev Physiol 1993;55:575~589
· ThorensR.Facilitated glucose transporters in epithelial cells.Ann Rev Physiol1993;55:591~608
· Simon.MI,Strathman MP,Gautam N.Diversity of G-protein in signaltransduction .Science 1991;252:802
· FantlWJ,Johnson DE,Williams LT.Signaling by receptor tyrosine kinases.Ann RevBiochem 1993;62:453~481
· StraderC,Fang TM,Dixon RAT.Structure and function of G-protein-coupled receptors.AnnRev Biochem 1994;63:101~132
· ClaphamD,Found M,Rochester MN.Direct G-protein activation of ion channel?Ann RevNeurosci 1994;17:441~464
· HodgkinAL.The ionic basis of nervous conduction.Science 1964;145:1148~1153
· BezanillaF,Steffani E.Voltage-dependent gating of ionic channels .Ann Rev Biophy andBiomol Structure 1994;23:819~846
· NeherE.Ion channels for communication between and within cells.Neuron 1992;8:605~612
· SakmannB.Elementary steps in synaptic transmission revealed by currents through singleion channels .Neuron 1992;8:613~629
· FrenchAS.Mechanotransduction.Ann Rev Physiol 1992;54:135~152
· ShengM,Greenberg MF.The regulation and function of c-fos and other immediate earlygenes in the nervous system .Neuron 1990;4:477~485
· FrancisArmstrong C,Jorgensen AO.Structure and development of E-C coupling units inskeletal muscle .Ann Rev Physiol 1994;56:509~534
· MottaPM.Ultrastructure of Smooth Muscle .Kluwer Academic Publishers.Lancaster,England,1990
· GalziJL,Revah F,Bessis A,Changeux JP.Functional architecture of the nicotinicacetylcholine receptor:from electric organ to brain .Ann Rev Pharmacol1991;31:37~72
· CatterallWA.Structure and function of voltage-gated ion channels.TINS 1993;16:500
· Huxley HE.The mechanism of muscle contraction .Science 1969;164:1355~1366
第三章 血液
血液是一种流体组织,充满于心血管系统中,在心脏的推动下不断循环流动。如果流经体内任何器官的血流量不足,均可能造成严重的组织损伤;人体大量失血或血液循环严重障碍,将危及生命。血液在医学诊断上有重要价值,因为很多疾病可导致血液组成成分或性质发生特征性的变化。
血液与内环境的演化血液是在动物进化过程中出现的。生命最初出现在海洋中。当在远古的海洋中出现比较复杂的多细胞生物时,机体的部分细胞已不可能与浸浴着整个机体的海洋环境直接接触;这时,机体内开始出现了细胞外液,它一方面作为细胞直接生活的内环境,同时又是机体与外环境进行物质交换的媒介。可以认为在进化中,最初的细胞外液可能是由包绕在机体内部的那部分海水形成的,因而它主要是一种盐溶液,其基本成分可能与远古的海水十分相似。以后,机体内出现了循环系统,细胞外液也进一步分化成为血管内的血浆和血管外的组织间隙液(简称组织液)。组织液仍然主要是盐溶液,是直接浸浴着绝大部分机体细胞的液体环境;而血管内的液体,则又溶入了多种蛋白质,并逐步出现了各种血细胞,于是形成了血液。
人体内除细胞外液外,尚有更多的液体(约为前者的2倍)存在于细胞内部,称为细胞内液。细胞外液和细胞内液总称为体液,约占机体总重量的60%。一般来说,细胞内液是细胞内各种生物化学反应得以进行的场所,细胞外液则是细胞直接生活的液体环境。因此,如果大气是整个人体的外环境,细胞外液就是第一个细胞生活的具体环境,故称为内环境。在细胞外液中,4/5在血管外构成组织液,1/5在血管内成为血浆的组成部分;而后者由于能在血管中不断循环流动,是内环境中最为活跃的部分,成为沟通各部分组织液以及和外环境进行物质交换的中间环节。
血液与内环境稳态在一些疾病情况下,常可出现内环境理化性质的较大变化,如高热、酸中毒、缺氧等,均将引起机体细胞功能的严重紊乱。在灌流离体器官的动物实验中,所用灌流液的化学成分、含氧量、Ph、温度与渗透压等,必须与这些动物的血浆十分相近(表3-1),离体器官才能在一段时间内保持接近于正常功能的活动状态。这些都说明,内环境理化性质的相对稳定,对于维持整个人体和体内所有细胞的正常功能都是非常必要的。这就是为什么早在19世纪时,Claude Bernard就指出:内环境的稳定是“机体自由独立生活的必要条件”。
然而在机体生活期间,干扰内环境理化性质的因素是不断出现的。机体细胞与细胞外液的物质交换,经常改变着内环境的理化性质;一些外环境因素的急剧变化也倾向于直接或间接(通过机体活动变化)改变内环境的理化性质。但与此同时,消化道不断补充营养物质,肺不断补充氧和排出二氧化碳,肾不断排出各种代谢尾产物、调整水与各种无机盐及小分子物质的排泄量,皮肤也不断散失代谢所产生的热量;而且,这些活动都处于整体的神经和体液调节之下,从而使内环境的理化性质只能作较小幅度的波动,保持着动态平衡。这一状态称为稳态(homeostasis)。
表3-1 血浆与生理盐溶液成分对照
| 任(Ringer)氏液(用于蛙心灌流)(g) | 蛙血浆(g) | 乐(Locke)氏液(用于哺乳动物)(g) | 哺乳动物血浆(g) | |
| NaCI | 0.65 | 0.55 | 0.9 | 0.7 |
| KCI | 0.014 | 0.023 | 0.042 | 0.038 |
| CaCI2 | 0.012 | 0.025 | 0.024 | 0.028 |
| NaHCO3 | 0.02 | 0.1 | 0.02 | 0.23 |
| NaH2PO4 | 0.001 | 0.002 | - | 0.036 |
| 葡萄糖 | - | 0.04 | 0.1-0.25 | 0.07 |
| 水 | 到100ml | 100ml | 到100ml | 100ml |
血液对内环境某些理化性质的变化有一定的“缓冲”作用。例如血液中含有多种缓冲物质,可以减轻酸性代谢产物引起的pH变化;血液中的水比热较大,可以吸收大量的热量而使温度升高不多。这类缓冲作用虽然有限,但在血液运输各种物质的过程中可防止其理化性质发生太大的变化。
血量人体内血液的总量称为血量,是血浆量和血细胞量的总和;但是,除红细胞外,其它血细胞数量很少,常可忽略不计。血浆量和红细胞量均可按稀释原理分别测定。例如可由静脉注射一定量不易透出血管的大分子染料(通常用T1824)或131I标记的血浆蛋白,待与体内血浆混匀后,再抽血测定T1824或131I被稀释的倍数,即可计算出血浆量。同样,可由静脉注射一定量用51Cr或32P标记的红细胞,待与体内的红细胞混匀后,抽血以测定标记的红细胞稀释的倍数,即可计算出红细胞总量。但由于标记的血浆白蛋白可逸出血管,因而从血流中“消失”较快,会影响测定结果,因此一般可先测出红细胞总量后,再按红细胞在血液中所占容积的百分比来推算血液总量。正常成年人的血液总量约相当于体重的7%~8%,或相当于每公斤体重70~80ml,其中血浆量为40~50ml。幼儿体内的含水量较多,血液总量占体重的90%。
第一节 血液的组成与特性
一、血液的组成
人类的血液由血浆和血细胞组成。1L血浆中含有900~910g水(90%~91%)。65~85g蛋白质(6.5%~8.5%)和20g低分子物质(2%).低分子物质中有多种电解质和小分子有机化合物,如代谢产物和其他某些激素等。血浆中电解质含量与组织液基本相同(表3-2)。由于这些溶质和水分都很容易透过毛细血管与组织液交流,这一部分液体的理化性质的变化常与组织液平行。在血液不断循环流动的情况下。血液中各种电解质的浓度,基本上代表了组织液中这些物质的浓度。
血浆蛋白的浓度是血浆和组织液的主要区别所在(表3-2),因为血浆蛋白的分子很大,不能透过毛细血管管壁。在生物化学研究中,曾经用盐析法将血浆蛋白分为白蛋白、球蛋白与纤维蛋白原三大类。以后,用电泳法又将白蛋白区分为白蛋白和前白蛋白,将球蛋白区分为a1-、a2-、a3-、β-、γ-球蛋白等。用其他方法,如免疫电泳,还可以将血浆蛋白作更进一步的区分。这说明血浆蛋白包括了很多分子大小和结构都不相同的蛋白质。
表3-2 人体各部分体液中电解质含量
| 血浆 | 组织液 | 细胞内液 | |||
| MEq/L(血浆) | MEq/L(水) | MEq/L(水) | MEq/L(水) | ||
| 正离子 | Na+ | 142.0 | 153.0 | 140.7 | 10 |
| K+ | 5.0 | 5.4 | 4.0 | 140 | |
| Ca2+ | 5.0 | 5.4 | 2.5 | 5 | |
| Mg2+ | 3.0 | 3.2 | 2.0 | 27 | |
| 总计 | 155.0 | 167.0 | 155.5 | 182 | |
| 负离子 | HCO3- | 27.0 | 29.0 | 30.0 | 10 |
| CI- | 103.0 | 111.0 | 114.0 | 25 | |
| HPO42- | 2.0 | 2.2 | 2.0 | 80 | |
| SO42- | 1.0 | 1.1 | 1.0 | 20 | |
| 有机酸 | 6.0 | 6.5 | 7.5 | ?/FONT> | |
| 蛋白质 | 16.0 | 17.2 | 1.0 | 47 | |
| 总计 | 155.0 | 167.0 | 155.5 | 182 | |
引自Koushanpour,E.1976(血浆依含水93%计算)
各种血浆蛋白具有不同的生理机能,主要有以下六下方面:
1、营养功能 每个成人3L左右的血浆中约含有200g蛋白质,它们起着营养贮备的功能。虽然消化道一般不吸收蛋白质,吸收的是氨基酸,但是,体内的某些细胞,特别是单核吞噬细胞系统,吞饮完整的血浆蛋白,然后由细胞内的酶类将吞入细胞的蛋白质分解为氨基酸。这样生成的氨基酸扩散进入血液,随时可供其它细胞合成新的蛋白质之用。
2、运输功能蛋白质巨大的表面上分布有众多的亲脂性结合位点,它们可以与脂容性物质结合,使之成为水溶性,便于运输;血浆蛋白还可以与血液中分子较小的物质(如激素、各种正离子)可逆性的结合,即可防止它们从肾流失,又由于结合状态与游离状态的物质处于动态平衡之中,可使处于游离状态的这些物质在血中的浓度保持相对稳定。
3、缓冲功能血浆白蛋白和它的钠盐组成缓冲对,和其它无机盐缓冲对(主要是碳酸和碳酸氢钠)一起,缓冲血浆中可能发生的酸碱变化,保持血液pH的稳定。
4、形成胶体渗透压,调节血管内外的水份分布(详见血液的理化特性一节)。
5、参与机体的免疫功能在实现免疫功能中有重要作用的免疫抗体、补体系统等,都是由血浆球蛋白构成的。
6、参与凝血和抗凝血功能绝大多数的血浆凝血因子、生理性抗凝物质以及促进血纤维溶解的物质都是血浆蛋白(参见本章第三节)。
二、血液的理化特性
(一)血液的比重
血液的比重为1.050~1.060,血浆的比重约为1.025~1.030。血液中红细胞数愈多则血液比重愈大,血浆中蛋白质含量愈多则血浆比重愈大。血液比重大于血浆,说明红细胞比重大于血浆。
红细胞的悬浮稳定性将与抗凝剂混匀的血液静置于一支玻璃管(如分血计)中,红细胞由于比重较大,将因重力而下沉,但正常时下沉十分缓慢。通常以红细胞在一小时内下沉的距离来表示红细胞沉降的速度,称为红细胞沉降率。正常男性的红细胞沉降率第一小时不超过3mm,女性不超过10mm。红细胞下降缓慢,说明它有一定的悬浮稳定性;红细胞沉降率愈小,表示悬浮稳定性愈大。
红细胞因比重较大而在血浆中下沉时,红细胞与血浆之间的磨擦则阻碍其下沉,特别是双凹碟形的红细胞,表面积与容积之比较大,因而所产生的磨擦也较大。红细胞沉降率在某些疾病时(如活动性肺结核、风湿热等)加快,这主要是由于许多红细胞能较快地互相以凹面相贴,形成一叠红细胞,称为叠连;红细胞叠连起来,其外表面积与容积之比减小,因而磨擦力减小,下沉加快。叠连形成的快慢主要决定于血浆的性质,而不在于红细胞自身。若将血沉快的病人的红细胞,置于正常人的血浆中,则形成叠连的程度和红细胞沉降的速度并不加大;反过来,若将正常人的红细胞置于这些病人的血浆中,则红细胞会迅速叠连而沉降。这清楚地说明促使红细胞发生叠连的因素在于血浆中。一般血浆中白蛋白增多可使红细胞沉降减慢;而球蛋白与纤维蛋白原增多时,红细胞沉降加速。其原因可能就在于白蛋白可使红细胞叠连(或聚集成其他形式有团粒)减少,而球蛋白与纤维蛋白原则可促使叠连(或其他形式的聚集)增多,但其详细作用机制尚不清楚。
(二)血液的粘滞性
通常是在体外测定血液或血浆与水相比的相对粘滞性,这时血液的相对粘滞性为4-5,血浆为1.6-2.4。全血的粘滞性主要决定于所含的红细胞数,血浆的粘滞性主要决定于血浆蛋白质的含量。水、酒精等在物理学上所谓“理想液体”的粘滞性是不随流速改变的,而血液在血流速度很快时类似理想液体(如在动脉内),其粘滞性不随流速而变化;但当血流速度小于一定限度时,则粘滞性与流速成反变的的关系。这主要是由于血流缓慢时,红细胞可叠连或聚集成其他形式的团粒,使血液的粘滞性增大。在人体内因某种疾病使微环境血流速度显着减慢时,红细胞在其中叠连和聚集,对血流造成很大的阻力,影响循环的正常进行;这时可以通过输入血浆白蛋白或低分子右旋糖酐以增加血流冲刷力量,使红细胞分散。
(三)血浆渗透压
血浆渗透压约为313mOsm/kgH2O,相当于7个大气压708.9kPa(5330mmHg)。血浆的渗透压主要来自溶解于其中的晶体物质,特别是电解质,称为晶体渗透压。由于血浆与组织液中晶体物质的浓度几乎相等,所以它们的晶体渗透压也基本相等。血浆中虽含有多量蛋白质,但蛋白质分子量大,所产生的渗透压甚小,不超过1.5mOsm/kgH2O,约相当于3.3kPa(25mmHg),称为胶体渗透压.由于组织液中蛋白质很少,所以血浆的胶体渗透压高于组织液.在血浆蛋白中,白蛋白的分子量远小于球蛋白,故血浆胶体渗透压主要来自白蛋白。若白蛋白明显减少,即使球蛋白增加而保持血浆蛋白总含量基本不变,血浆胶体渗透压也将明显降低。
血浆蛋白一般不能透过毛细血管壁,所以血浆胶体渗透压虽小,但对于血管内外的水平衡有重要作用(参见第四章第三节)。由于血浆和组织液的晶体物质中绝大部分不易透过细胞膜,所以细胞外液的晶体渗透压的相对稳定,对于保持细胞内外的水平衡极为重要。
等渗溶液与等张溶液在临床或生理实验使用的各种溶液中,其渗透压与血浆渗透压相等的称为等渗溶液(如0.85%NaCI溶液),高于或低于血浆渗透压的则相应地称为高渗或低渗溶液。将正常红细胞悬浮于不同浓度的NaCI溶液中即可看到:在等渗溶液中的红细胞保持正常大小和双凹圆碟形;在渗透压递减的一系列溶液中,红细胞逐步胀大并双侧凸起,当体积增加30%时成为球形;体积增加45%~60%则细胞膜损伤而发生溶血,这时血红蛋白逸出细胞外,仅留下一个双凹圆碟形细胞膜空壳,称为影细胞(ghost cell)。正常人的红细胞一般在0.42%NaCI溶液中时开始出现溶血,在0.35%NaCI溶液中时完全溶血.在某些溶血性疾病中,病人的红细胞开始溶血及完全溶血的NaCI溶液浓度均比正常人高,即红细胞的渗透抵抗性减小了,渗透脆性增加了。不同物质的等渗溶液不一定都能使红细胞的体积和形态保持正常;能使悬浮于其中的红细胞保持正常体积和形状的盐溶液,称为等张溶液.所谓“张力”实际是指溶液中不能透过细胞膜的颗粒所造成的渗透压。例如NaCI不能自由透过细胞膜,所以0.85%NaCI既是等渗溶液,也是等张溶液;但如尿素,因为它是能自由通过细胞膜的,1.9%尿素溶液虽然与血浆等渗,但红细胞置入其中后立即溶血。所以不是等张溶液。
(四)血浆的pH值
正常人的血浆的pH值约为7.35-7.45。血浆pH值主要决定于血浆中主要的缓冲对,即NaHCO3/H2CO3的比值。通常NaHCO3/H2CO3比值为20。血浆中NaHCO3/H2CO3外,尚有其他缓冲对。在血浆中有蛋白质钠盐/蛋白质、Na2HPO4/NaH2PO4,在红细胞内尚有血红蛋白钾盐/血红蛋白、氧合血红蛋白钾盐/氧合血红蛋白、Na2HPO4/NaH2PO4、KH2PO4、KHCO3/H2CO3等缓冲对,都是很有效的缓冲对系统。一般酸性或碱性物质进入血液时,由于有这些缓冲系统的作用,对血浆pH值的影响已减至很小,特别是在肺和肾不断的排出体内过多的酸或碱的情况下,通常血浆pH值的波动范围极小。
第三节 血细胞及其功能
血细胞包括红细胞、白细胞和血小板三类细胞,它们均起源于造血干细胞。在个体发育过程中,造血器官有一个变迁的程序。在胚胎发育的早期,是在卵黄囊造血,从胚胎第二个月开始,由肝、脾造血;胚胎发育到第五个月以后,肝、脾的造血活动逐渐减少,骨髓开始造血并逐渐增强;到婴儿出生时,几乎完全依靠骨髓造血,但在造血需要增加时,肝、脾可再参与造血以补充骨髓功能的不足。因此,此时的骨髓外造血具有代偿作用。儿童到4岁以后,骨髓腔的增长速度已超过了造血组织增长的速度,脂肪细胞逐步填充多余的骨髓腔。到18岁左右,只有脊椎骨、肋骨、胸骨、颅骨和长骨近端骨骺处才有造血骨髓,但造血组织的总量已很充裕。成年人如果出现骨髓外造血,已无代偿的意义,而是造血功能紊乱的表现。
造血过程,也就是各类血细胞的发育、成熟的过程,是一个连续而又区分为阶段的过程。首先是造血干细胞(hemopietic stem cells)阶段,处于这一阶段的造血细胞为干细胞,它们既能通过自我复制(self renewal)以保持本身数量的稳定,又能分化形成各系定向祖细胞(committedprogenitors);第二个阶段是定向祖细胞阶段,处于这个阶段的造血细胞,进一步分化方向已经限定,它们可以区分为:红系祖细胞,即红系集落形成细胞(CFU-E),粒-单核系祖细胞(CFU-GM),巨核系祖细胞( CFU-MK)和 TB淋巴系祖细胞(CFU-TB);第三个阶段是形态可辩认的前体细胞(precursors)阶段,此时的造血细胞已经发育成为形态上可以辨认的各系幼稚细胞,这些细胞进一步分别成熟为具有特殊细胞功能的各类终末血细胞,然后释放进入血液循环。造血细胞在经历上述发育成熟过程中,细胞自我复制的能力逐渐降低,而分化、增殖的能力逐渐增强,细胞数量逐步增大(图3-1)
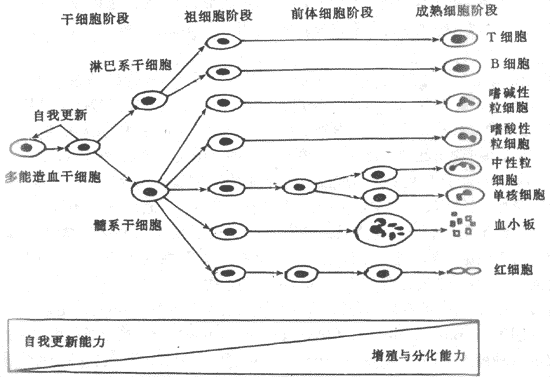
图3-1造血细胞发育模式图
一、红细胞生理
1.红细胞的数量、形态和功能红细胞(erythuocyte)是血液中数量最多的一种血细胞,正常男性每微升血液中平均约500万个(5.0×1012/L),女性较少,平均约420万个(4.2×1012/L).红细胞含有血红蛋白,因而使血液呈红色.红细胞在血液的气体运输中有极重要的作用.在血液中由红细胞运输的氧约为溶解于血浆的70倍;在红细胞参与下,血浆运输二氧化碳的能力约为直接溶解于血浆的18倍(详见第五章第三节).正常红细胞呈双凹圆碟形,平均直径约8μm,周边稍厚.这种细胞开头的表面积与体积之比,较球形时为大,因而气体可通过的面积也较大;由细胞中心到大部分表面的距离较短,因此气体进出红细胞的扩散距离也较短.这种形状也有利于红细胞的可塑性变形.红细胞在全身血管中循环运行,常要挤过口径比它小的毛细血管和血窦间隙,这时红细胞将发生卷曲变形,在通过后又恢复原状,这种变形称为塑性变形.表面积与体积的比值愈大,变形能力愈大,故双凹圆碟形红细胞的变形能力远大于异常情况下可能出现的球形红细胞.红细胞保持双凹圆碟形需要消耗能量。
红细胞膜是以脂质双分子层为骨架的半透膜。氧和二氧化碳等脂溶性气体可以自由通过,尿素也可以自由透入。在电解质中,负离子(如CI-、HCO3-)一般较易通过红细胞膜,而正离子却很难通过。红细胞内Na+浓度远低于细胞外,而细胞内K+浓度远高于细胞外,这种细胞内外的Na+、K+浓度差主要是依靠细胞膜上Na+泵的活动来维持的。低温贮存较久的血液,血浆内K+浓度升高,就是由于低温下代谢几乎停止,Na+泵不能活动的缘故。
红细胞结合和携带氧的过程并不消耗能量,血红蛋白中的Fe2+也不被氧化,若Fe2+被氧化成Fe3+成为高铁血红蛋白,即失去携氧能力。红细胞消耗葡萄糖,主要是通过糖酵解和磷酸戊糖旁路,所产生的能量(以结合于ATP的形式)主要是用于供应细胞膜上Na+泵的活动,用于保持低铁血红蛋白不致被氧化,也用于保持红细胞膜的完整性和细胞的双凹圆碟形。
2.红细胞比容红细胞在血液中所占的容积百分比,称为红细胞比容(hematocritvalue),可以用分血计(hematocrit)来测定。通常是将一定量的血液与抗凝剂混匀,置于用直径2.5mm的平底玻璃管制成的分血计中,以每分钟3000转的速度离心半小时,使血细胞下沉压紧,即可测出红细胞比容。正常成年人的红细胞比容,男性为40%-50%,女性为37%-48%。但这是从手臂等处浅静脉抽血测定的数值,并且这时在压紧的红细胞之间有很少量血浆;同时,全身各类血管中,血液的红细胞比容值也不尽相同。
3.正常红细胞生成所必需的原料和其它因素在幼红细胞的发育成熟过程中,细胞核的存在对于细胞分裂和合成血红蛋白有着重要的作用。在这些阶段,合成细胞核的主要构成物质—DNA必须有维生素B12和叶酸作为辅酶。
维生素B12是含钴的有机化合物,多存在于动物性食品中。机体对维生素B12的吸收必须要有内因子(intrinsic factor)和R结合蛋白(Rprotein)参与。内因子是由胃腺的壁细胞所分泌的一种糖蛋白,分子量在50000-60000之间,而R(rapid)蛋白是一种电泳速度很快的血浆蛋白。在酸性的胃液中,维生素B12主要与R蛋白结合,到了小肠上段处胰蛋白酶将这种结合断裂,维生素B12转而与内因子结合。内因子有两个活性部位,一个部位可与维生素B12结合,另一个部位则可与回肠上皮细胞膜上的特异受体结合。在正常情况下,内因子-B12复合物在小肠上段可保护维生素B12不受小肠内蛋白水解酶的破坏。当复合物运行至回肠段,便与回肠粘膜受体结合而被吸收进入门脉系统血流,一部分贮存在肝,一部分又与运输维生素B12的转钴蛋白Ⅱ(transcobalamineⅡ)结合,沿血液输送到造血组织,参与红细胞生成过程。当胃的大部分被切除或胃腺细胞受损伤,机体缺乏内因子,或体内产生抗内因子的抗体时,即可发生维生素B12吸收障碍,影响幼红细胞的分裂和血红蛋白合成,出现巨幼红细胞性贫血,即大细胞性贫血。
叶酸是以蝶酰单谷氨酸的形式吸收的。吸收之后,在双氢叶酸还原酶的催化下,形成四氢叶酸。存在于血浆中的叶酸几乎全是四氢叶酸的单谷氨酸盐。但进入组织细胞后,又通过酶促作用,再转变为多谷氨酸盐,才具有活性。叶酸缺乏时也引起与维生素B12缺乏时相似的巨幼红细胞性贫血。只是在维生素B12缺乏时,还可伴有神经系统和消化道症状。
合成血红蛋白还必须有铁作为原料,每亳升红细胞需要1mg铁,每天需要20-25mg铁用于红细胞生成,但人每天只需从食物中吸收1mg(约5%)以补充排泄的铁,其余95%均来自人体铁的再利用。机体贮存的铁主要来自于破坏了的红细胞。衰老的红细胞被巨噬细胞吞噬后,血红蛋白被消化而释出血红素中的Fe2+。这样释出的铁即与铁蛋白(ferritin)结合,此时的铁为Fe3+,聚集成铁黄素颗粒而沉淀于巨噬细胞内。血浆中有一种运铁蛋白(transferrin),可以来往运行于巨噬细胞与幼红细胞之间,以运送铁。贮存于铁蛋白中的Fe3+,先还原成Fe2+再脱离铁蛋白,而后与运铁蛋白结合。每分子运铁蛋白可以运送两个Fe2+,运送到幼红细胞后,又可反复作第二次运输。此外,还可以通过巨噬细胞与红母细胞直接接触,以提供合成血红蛋白所需的铁。由于慢性出血等原因,体内贮存的铁减少,或造血功能增强而供铁不够,均可引起小细胞性贫血,这主要是合成血红蛋白不足。此外,红细胞生成还需要氨基酸和蛋白质、维生素B6、B2、C、E,微量元素铜、锰、钴和锌等。
4.红细胞生成的调节每个成年人体内约有25×1012个红细胞,每24小时便有0.8%的红细胞进行更新,也就是说每分钟约有160×106个红细胞生成;当机体有需要时,如失血或某些疾病使红细胞寿命缩短时,红细胞的生成率还能在正常基础上增加数倍。目前已经证明有两种调节因子分别调制着两个不同发育阶段红系祖细胞的生长。一种是早期的红系祖细胞,称为爆式红系集落形成单位(burst forming unit-erythroid,BFU-E),这是因为它们在体外培养中能形成很大的细胞集落,组成集落的细胞散布成物体爆炸的形状,这种早期祖细胞的生长和在体外形成集落都依赖于一种称为爆式促进因子(burst promoting activitor,BPA)的刺激作用。BPA是一类分子量为25000-40000的糖蛋白,以早期红系祖细胞BFU-E为作用的靶细胞,可能是促进更多的BFU-E从细胞周期中的静息状态(G期)进入DNA合成期(S期),因而使早期祖细胞加强增殖活动。另一种是晚期的红系祖细胞,称为红系集落形成单位(colony forming unit-erythroid,CFU-E),它们在体外培养中只能形成较小的集落。晚期红系祖细胞对BPA不敏感,但主要接受促红细胞生成素(erythropoietin,EPO)的调节。促红细胞生成素是一种热稳定(80℃)的糖蛋白,分子量为34000。当组织中氧分压降低时,血浆中的促红细胞生成素的浓度增加,它促进红系祖细胞向前体细胞分化,又加速这些细胞的增殖,结果使骨髓中能合成血红蛋白的幼红细胞数增加,网织红细胞加速从骨髓释放。早在本世纪50年代,动物实验已显示了促红细胞生成素活性的存在,以后又确定促红细胞生成素主要由肾组织产生。切除双肾后,血浆中促红细胞生成素的浓度急剧降低。用分子生物学手段进一步证明,从肾组织细胞中已提取出编码促红细胞生成素的Mrna 和Cdna,还确定促红细胞生成素和mRNA和cDNA,还确定促红细胞生成素基因定位在7号染色体上。近年来有迹象提示人类的某些血液病,如再生障碍性贫血是红系祖细胞促红细胞生成素受体有缺陷所致(图3-2)。
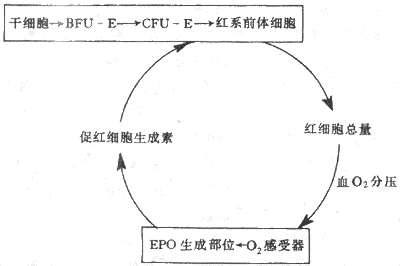
图3-2 EPOA调节红细胞生成的反馈调节环
促红细胞生成素主要由肾组织产生,但肾外,如肝脏,也有小量生成。晚期肾病患者,肾脏产生EPO已基本停止,但体内仍有小量EPO促使骨髓继续产生红细胞。
其他一些激素,包括雄激素、甲状腺激素和生长激素,都可增强促红细胞生成素的作用;雌激素则有抑制红细胞生成的作用。这可能是男性的红细胞数和血红蛋白量高于女性的原因。
二、白细胞生理
白细胞(leukocyte)是一类有核的血细胞。正常成年人白细胞总数是4000-10000/μ1,每日不同的时间和机体不同的功能状态下,白细胞在血液中的数目是有较大范围变化的。当每微升超过10000个白细胞时,称为白细胞增多,而每微升少于4000个白细胞时,称为白细胞减少。机体有炎症时常出现白细胞增多。
白细胞不是一个均一的细胞群,根据其形态、功能和来源部位可以分为三大类:粒细胞、单核细胞和淋巴细胞(表3-3)。白细胞与红细胞和血小板一样都起源于骨髓中的造血干细胞,在细胞发育过程中又都是经历定向祖细胞、前体细胞,而后成为具有各种细胞功能的成熟白细胞。
表3-3 血液中各类白细胞计数(细胞/μL)
| 均数 | % | 范围 | |
| 粒细胞 | |||
| 中性粒细胞 | 4150 | (59) | 1712~7588 |
| 嗜酸性粒细胞 | 165 | (2) | 0~397 |
| 嗜碱性粒细胞 | 44 | (<1) | 0~112 |
| 单核细胞 | 456 | (7) | 66~846 |
| 淋巴细胞 | 2185 | (31) | 1029~3341 |
| 白细胞 | 7000 | 2800~11200 |
引自Schmidt,R.F.&Thews,G,1989
白细胞的分化和增殖受到一组造血76生长因子(hematopoietic growth factor,HGF)的调节。这些因子从淋巴细胞、单核细胞和成纤维细胞生成并分泌,是一类糖蛋白。由于有些造血生长因子在体外可刺激造血细胞生成集落,故又称为集落刺激因子(clony stimulating factor, CSF)。目前从结构到功能已经充分阐明的集落刺激因子有M-CSF、G-CSF、GM-CSF、Multi-CSF、Meg-CSFt 和EPO等6种,除了EPO是调节红细胞生成因子之外,其余因子均参与调节白细胞的生成。这些因子中有的作用是广谱的,如Multi-CSF和GM-CSF(G是粒细胞缩写,M是单核细胞的缩写)的作用可以影响多系造血祖细胞的生成和发育,而其它一些因子(如G-CSF、M-CSF、GM-CSF)作用较为局限,只作用于某一系的造血祖细胞。所有这些因子除了作用于祖细胞,还能影响成熟白细胞的功能。此外,还有一类抑制性因子,如粒细胞抑素、乳铁蛋白和转化生成因子-β等,它们或是直接抑制白细胞的增殖、生长、或是限制上述的一些生长因子的释放或作用。
淋巴细胞的生成过程与其它白细胞有一些不同。在干细胞分化的早期,淋巴干细胞首先从多能干细胞分化出来。这些淋巴干细胞随血流进入初级(或中枢)淋巴器官,即骨髓和胸腺,在这里它们发育成定向淋巴细胞(commmittde lymphocyte)。在骨髓中发育的称为B细胞;在胸腺中发育的称为T细胞。随后,B和T细胞均随血流转移到二级(或外周)淋巴器官,即淋巴结和脾,在那里它们与某种抗原接触后即分化和增殖成为真正具有免疫功能的细胞,如浆细胞和T效应细胞(t effector cell)。淋巴细胞在生长成熟过程中接受一组称为白细胞介素(interleukins,ILs)的细胞因子的调节,T细胞在胸腺中还接受胸腺激素的作用(图3-3)。
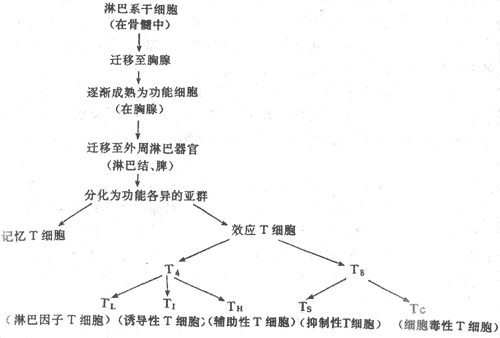
图3-3 T细胞的成熟和分化
所有的白细胞都能作变形运动,凭藉这种运动白细胞得以穿过血管壁,这一过程称作血细胞渗出(diapedisis)。白细胞具有趋向某些化学物质游走的特性,称为趋化性。体内具有趋化作用的物质包括:细菌毒素、细菌或人体细胞的降解产物,以及抗原-性体复合物等。白细胞按照这些物质的浓度梯度游走到这些物质的周围,把异物包围起来并吞入胞浆内,这称为吞噬作用。每类白细胞都具有某些酶类,如蛋白酶、多肽酶、淀粉酶、脂酶和脱氧核糖核酸酶等。在白细胞总数中,有一半以上存在于血管外的细胞间隙内,有30%以上贮存在骨髓内,其余的才是在血管中流动的。这些白细胞凭藉血液的运输,从它们生成的器官,即骨髓和淋巴组织,到达发挥作用的部位。
(一)粒细胞
约有60%的白细胞的胞质内具有颗粒,因而把它们称为粒细胞。又根据胞质中颗粒的染色性质不同将粒细胞区分为:中性、嗜酸性和嗜碱性粒细胞,这三类细胞的比例见表3-3。粒细胞在血流中停留时间很短暂,一般从数小时至2天。
1.中性粒细胞绝大部分的粒细胞属中性粒细胞。每微升血液中约有4500个中性粒细胞。由于这些细胞的细胞核的形态特殊,又称为多形核白细胞。中性粒细胞在血管内停留的时间平均只有6-8小时,它们很快穿过血管壁进入组织发挥作用,而且进入组织后不再返回血液中来。在血管中的中性粒细胞,约有一半随血流循环,通常作白细胞计数只反映了这部分中性粒细胞的情况;另一半则附着在小血管壁上。同时,在骨髓中尚贮备了约2.5×1012个成熟中性粒细胞,在机体需要时可立即动员大量这部分粒细胞进入循环血流。
中性粒细胞在血液的非特异性细胞免疫系统中起着十分重要的作用,它处于机体抵御微生物病原体,特别是在化脓性细菌入侵的第一线,当炎症发生时,它们被趋化性物质吸引到炎症部位。由于它们是藉糖酵解获得能量,因此在肿胀并血流不畅的缺氧情况下仍能够生存,它们在这里形成细胞毒存在破坏细菌和附近组织的细胞膜。由于中性粒细胞内含有大量溶酶体酶,因此能将吞噬入细胞内的细菌和组织碎片分解,这样,入侵的细菌被包围在一个局部,并消灭,防止病原微生物在体内扩散。当中性粒细胞本身解体时,释出各溶酶体酶类能溶解周围组织而形成脓肿。
中性粒细胞的细胞膜能释放出一种不饱和脂肪酸——花生四烯酸,在酶的作用下,由它再进一步生成一组旁分泌激素物质,如血栓素和前列腺素等,这类物质对调节血管口径和通透性有明显的作用,还能引起炎症反应和疼痛,并影响血液凝固(参看本章第三节及图3-8)。
2.嗜碱性粒细胞在白细胞中嗜碱性细胞占0.5%-1%,即50个细胞/μ1。平均循环时间是12小时。这类粒细胞的胞质中存在较大和碱性染色很深的颗粒。颗粒内含有肝素和组织胺。近年来发现嗜碱性粒细胞参与体内的脂肪低谢。当食物中的脂肪被肠吸收后,周围血液中的嗜碱性粒细胞数随即增加。嗜碱性粒细胞释放出肝素(heparin),激活在血浆中的脂肪分解。这是由于肝素作为脂酶的辅基增强了脂酶的作用。结果加快了由脂肪分解为游离脂肪酸的过程。
嗜碱性粒细胞释放的组胺与某些异物(如花粉)引起过敏反应的症状有关(请参考免疫学教材)。
此外,嗜碱性粒细胞被激活时还释放一种称为嗜酸性粒细胞趋化因子A(eosinophile chemotactic factor A)的小肽,这种因子能把嗜酸性粒细胞吸引过来,聚集于局部以限制嗜碱性粒细胞在过敏反应中的作用。
3.嗜酸性粒细胞血液中嗜酸性粒细胞占白细胞总数的2%-4%,即100-350个细胞/μ1。血液中嗜酸性粒细胞的数目有明显的昼夜周期性波动,清晨细胞数减少,午夜时细胞数增多。这种细胞数的周期性变化是与肾上腺皮质释放糖皮质激素量的昼夜波动有关的。当血液中皮质激素浓度增高时,嗜酸性粒细胞数减少;而当皮质激素浓度降低时,细胞数增加。嗜酸性粒细胞的胞质内含有较大的、椭圆形的嗜酸性颗粒。这类白细胞也具有吞噬功能。嗜酸性粒细胞在体内的作用是:①限制嗜碱性粒细胞在速发性过敏反应中的作用。当嗜碱性粒细胞被激活时,释放出趋化因子,使嗜酸性粒细胞聚集到同一局部,并从三个方面限制嗜碱性粒细胞的活性:一是嗜酸性粒细胞可产生前列腺素E使嗜碱性粒细胞合成释放生物活性物质的过程受到抑制;二是嗜酸性粒细胞可吞噬嗜碱性粒细胞所排出的颗粒,使其中含有生物活性物质不能发挥作用;三是嗜酸性粒细胞能释放组胺酶等酶类,破坏嗜碱性粒细胞所释放的组胺等活性物质。②参与对蠕虫的免疫反应。在对蠕虫的免疫反应中,嗜酸性粒细胞有重要的作用。这类粒细胞的细胞膜上分布有免疫球蛋白Fc片断和补体C3的受体。在已经对这种蠕虫具有免疫性的动物体内,产生了特异性的免疫球蛋白IgE。蠕虫经过特异性IgE和C3的调理作用后,嗜酸性粒细胞可借助于细胞表现的Fc受体和C3受体粘着于蠕虫上,并且利用细胞溶酶体内所含的过多氧化物酶等酶类损伤蠕虫体。在有寄生虫感染、过敏反应等情况时,常伴有嗜酸性粒细胞增多。
(二)单核细胞
第二类白细胞称为单核细胞,胞体较大,直径约为15-30酶μm,胞质内没有颗粒,它们约占血液中白细胞数的4%-8%。单核细胞来源于骨髓中的造血干细胞,并在骨髓中发育。当它们从骨髓进入血流时仍然是尚未成熟的细胞。与其他血细胞比较,单核细胞内含有更多的非特异性脂酶,并且具有更强的吞噬作用。单核细胞在血液中停留2-3天后迁移到周围组织中,细胞体积继续增大,直径可达50-80μm,细胞内所含的溶酶体颗粒和线粒体的数目也增多,成为成熟的细胞。固定在组织中的单核细胞称为组织巨噬细胞,它们经常大量存在于淋巴结、肺泡壁、骨髓、肝和脾等器官。激活了的单核细胞和组织巨噬细胞能生成并释放多种细胞毒、干扰素和白细胞介素,参与机体防卫机制,还产生一些能促进内皮细胞和平滑肌细胞生长的因子。在炎症周围单核细胞能进行细胞分裂,并包围异物。
(三)淋巴细胞
淋巴细胞是免疫细胞中的一大类,它们在免疫应答过程中起着核心作用。根据细胞成长发育的过程和功能的不同,淋巴细胞分成T细胞和B细胞两类。在功能上T细胞主要与细胞免疫有关,B细胞则主要与体液免疫有关。
1.T细胞在血液的淋巴细胞中,约占70%-80%,在血液和淋巴组织之间反复循环,还可以停留在外周淋巴器官如淋巴结中。淋巴细胞的寿命较长,一般为数月,有的长达一年以上。T细胞被特异性的抗原物质激活后,进行增殖和分化,形成在功能上各异的两类细胞,即T免疫效应细胞T记忆细胞(t memory cell)。
根据T效应细胞的细胞表面特征的不同可区分为T4和T8两个亚群,而这些亚群还可根据不同的功能再分为不同类型。属于T4亚群的有:淋巴因子T细胞(t lymphokine cell,TL)、诱导性T细胞(T inductor cells,T1)和辅助性T细胞(t helper cells, TH)。淋巴因子T细胞能通过释放淋巴因子激活巨噬细胞和造血干细胞;T诱导性细胞能释放白细胞介素-2(interlukin-2),促进其他T细胞的成熟分化,而辅助性T细胞能产生一种B细胞生长因子(b cell growth factor),促使B细胞分化为浆细胞,影响抗体的产生。
T8亚型细胞,根据其功能可以再分为能抑制B细胞和T细胞活性的抑制性T细胞(t suppressor cells,Ts),和对带有特异抗原的靶细胞具有杀伤作用的细胞毒性T细胞(t cytotoxic cells, Tc)。由此可见,T细胞除了具有细胞免疫作用外,它们还具有调节其他免疫细胞特别是B细胞的功能。
长寿命的记忆T细胞在血液中不断循环,当他们再次遇到曾经接触过的抗原时,即使相隔几年之久仍能加以“识别”。在第二次与抗原体接触时能激发一种继发反应,这种反应比原发反应更强烈的引起细胞增殖,在短时间内形成大量的效应T细胞。T细胞各亚群的关系见图3-3。
2.B细胞在血液中B细胞约占淋巴细胞总数的15%。固定在B细胞膜表面的免疫球蛋白(主要是单体IgM和IgD)是抗原的特异性受体。当它们初次与某一个抗原接触而被致敏时,一部分B细胞即分化成熟为浆细胞,浆细胞即开始生成对该抗原特异的免疫球蛋白并将它们释放到周围的组织液中,这就是免疫抗体。只有当某些调节性因子,如由辅助性T细胞所释放的淋巴因子和巨噬细胞释放的白细胞介素-1存在时,B细胞才能被抗原激活。浆细胞不再在血液中循环,在它们生存的2-3天时间里一直停留在组织中。
有小部分受抗原刺激的B细胞发展成为记忆性B细胞,寿命很长,且保持特异性,由它们增殖生成的后代细胞也保持着这种特异性。当它们再次接触具有同样特异性的抗原时,便能迅速被激活,成为特异B淋巴母细胞。由记忆性B细胞增殖生成的后代细胞愈多,被特异性抗原、激活的B细胞数也愈多。可见B细胞系统的“记忆”能力是取决于具有抗原特异性的记忆细胞数目的多少。
在血液中,除了T细胞和B细胞之外还有一类淋巴细胞,根据它们的细胞表面标志既不归属于B细胞,也不归属于T细胞。这类细胞称为裸细胞(null cell),约占血液中淋巴细胞总数的5%-10%。目前受注意的裸细胞有杀伤细胞(killer cell,K细胞)和自然杀伤细胞(matiral killer cell,NK细胞),K细胞上具有免疫球蛋白IgG的Fc片段受体,当表面覆盖有IgG(抗体)的靶细胞与K细胞接触时,IgG分子的Fc片段可与K细胞表面的Fc受体结合,激发K细胞的杀伤作用。由此可见,K细胞的杀伤作用是抗原依赖性的,但抗原是非特异的。至于NK细胞,虽然也是杀伤细胞,但其杀伤作用不依赖于抗原和抗体的存在。NK细胞广泛分布在血液和外周淋巴器官,对杀伤肿瘤细胞有重要作用。干扰素能活化NK细胞,而白细胞介素-2能刺激NK细胞的增殖和产生干扰素,因而增强NK细胞的杀伤作用。
三、血小板生理
血小板(platelets, thrombocyte)是从骨髓成熟的巨核细胞胞浆解脱落下来的小块胞质。巨核细胞虽然在骨髓的造血细胞中为数最少,仅占骨髓有核细胞总数的0.05%,但其产生的血小板却对机体的止血功能极为重要。每个巨核细胞均可产生1000-6000个血小板。
正常成年人的血小板数量是150000-350000个/μ/(150-350×109/L)。血小板有维护血管壁完整性的功能。当血小板数减少到50000个/μl(50×109/L)以下时,微小创伤或仅血压增高也使皮肤和粘膜下出现血瘀点,甚至出现大块紫癜。可能由于血小板能随时沉着于血管壁以填充内皮细胞脱落留下的空隙;而且,用同位素标记血小板示踪和电子显微镜观察,发现血小板可以融合入血管内皮细胞,因而可能对保持内皮细胞完整或对内皮细胞修复有重要作用。当血小板太少时,这些功能就难以完成而产生出血倾向。
循环血液中的血小板一般处于“静止”状态。但当血管受损伤时,通过表面接触和某些凝血因子的作用,血小板转入激活状态。激活了的血小板能释放一系列对止血过程必需的物质,关于血小板在止血过程中的作用可参看本章第三节。
生成血小板的巨核细胞也是从骨髓中的造血干细胞分化发展来的。造血干细胞首先分化生成巨核系祖细胞,也称巨核系集落形成单位(colony forming unit-megakaryocyte,CFU-Meg)。祖细胞阶段的细胞核内的染色体一般是2-3倍体。当祖细胞是2倍体或4倍体时,细胞具有增殖能力,因此这是巨核细胞系增加细胞数量的阶段。当巨核系祖细胞进一步分化为8-32倍体的巨核细胞时,胞质开始分化,内膜系统逐渐完备。最后有一种膜性物质把巨核细胞的胞质分隔成许多小区。当每个小区被完全隔开时即成为血小板,一个个血小板通过静脉窦窦壁内皮间的空隙从巨核细胞脱落,进入血流。
巨核细胞增殖、分化的调节机制类似于红细胞系生成的调节,至少受两种调节因子分别对两个分化阶段进行调节。这两种调节因子是:巨核系集落刺激因子(Meg-CSF)和促血小板生成素(thrombopoietin,TPO)。
巨核系集落刺激因子是主要作用于祖细胞阶段的调节因子,它的作用是调节巨核系祖细胞的增殖。骨髓中巨核细胞总数减少时促使该调节因子的生成增加,Meg-CSF是一种低分子糖蛋白,分子量约为46000,它与促血小板生成素具有完全不同的免疫学性质。
促血小板生成素也是一种糖蛋白,当血流中血小板减少时,促血小板生成素在血液中的浓度即增加。该调节因子的作用包括:①增强祖细胞的DNA合成和增加细胞多倍体的倍数;②刺激巨核细胞合成蛋白质;③增加巨核细胞的总数,结果增加了血小板的生成。根据去肾大鼠出现血小板减少时血液中促血小板生成素的浓度不增加的事实,推测肾是产生促血小板生成素的部位。
四、血细胞的破坏
血细胞常因衰老而被破坏,但也可因意外和各种病理原因而被破坏。破坏的方式各种血细胞不尽相同,这与各种细胞功能不同有关。
(一)红细胞的破坏
红细胞的平均寿命约为120天。在这期间,平均每个红细胞血管内循环流动约27km,在“旅途”中常常需要挤过去比它小的毛细血管及孔隙,因而不得不变形。当红细胞逐渐衰老时,细胞变形能力减退而脆性增加,在血流湍急处可因机械冲击而破损,在通过微小孔隙时也发生困难,因而特别容易停滞在脾和骨髓中被巨噬细胞所吞噬。事实上,任何组织都能使红细胞解体,这可从皮下出血的青紫块都会逐渐消失的事实证明。
红细胞在血管内破损而发生溶血,所释出的血红蛋白立即与一种血浆a2-球蛋白—触珠蛋白结合;但溶血严重达到每100m1血浆有100mg血红蛋白时,血浆中的触珠蛋白已不够用,未能与触珠蛋白结合的血红蛋白将经肾从尿中排出。与触珠蛋白结合的血红蛋白虽不致被排出,但将被肝摄取,脱铁血红素转变为胆色素,铁则以铁黄素的形式沉着于肝细胞内。在脾内被吞蚀的衰老红细胞,经消化后,铁可再利用,而脱铁血红素也转变为胆色素,运送到肝处理。
(二)血小板的破坏
血小板进入血液后,只在开始两天具有生理功能,但平均寿命可有7-14天。
在生理止血活动中,血小板聚集后本身将解体并释出全部活性物质;它也可能融入血管内皮细胞。这样看来,血小板除衰老破坏外,还可能在发挥其生理功能时被消耗。但是用51Cr或32P标记血小板观察其破坏的情况,发现血小板的破坏也随血小板的“日龄”而增多,即主要是衰老尔后被破坏。曾经对一些接受了抗凝处理的人进行观察,发现这时血凝过程虽被阻止,但血小板的寿命也不延长。这些事实,倾向于说明了平时的血管内凝血并不强(也可能这种凝血程度很小),不致影响全部血小板的平均寿命。衰老的血小板是在脾、肝和肺组织中被吞噬的。
白细胞的寿命较难准确判断。因为,粒细胞和单核细胞主要是在组织中发挥作用的;淋巴细胞则往返循环于血液-组织液-淋巴之间,而且尚可增殖分化。一般来说,中性粒细胞在循环血液中停留8小时左右即进入组织,一般三、四天后将衰老死亡;若有细菌入侵,粒细胞在吞噬活动中可释出溶酶体酶过多而发生“自我溶解”,与破坏的细菌和组织片共同构成脓。
第三节 生理止血、血液凝固与纤维蛋白溶解
小血管损伤后血液将从血管流出,但在正常人,数分钟后出血将自行停止,称为生理止血。用一个小撞针或注射针刺破耳垂或指尖使血液流出,然后测定出血延续的时间,这一段时间称为出血时间(bleeding time)。出血时间的长短可以反映生理止血功能的状态。正常出血时间为1-3分钟。血小板减少,出血时间即相应延长,这说明血小板在生理止血过程中有重要作用;但是血浆中一些蛋白质因子所完成的血液凝固过程也十分重要。凝血有缺陷时常可出血不止。
生理止血过程包括三部分功能活动。首先是小血管于受伤后立即收缩,若破损不大即可使血管封闭;主要是由损伤刺激引起的局部缩血管反应,但持续时间很短。其次,更重要的是血管内膜损伤,内膜下组织暴露,可以激活血小板和血浆中的凝血系统;由于血管收缩使血流暂停或减缓,有利于激活的血小板粘附于内膜下组织并聚集成团,成为一个松软的止血栓以填塞伤口。接着,在局部又迅速出现血凝块,即血浆中可溶的纤维蛋白源转变成不溶的纤维蛋白分子多聚体,并形成了由血纤维与血小板一道构成的牢固的止血栓,有效地制止了出血。与此同时,血浆中也出现了生理的抗凝血活动与纤维蛋白溶解活性,以防止血凝块不断增大和凝血过程漫延到这一局部以外。显然,生理止血主要由血小板和某些血浆成分共同完成。
一、血凝、抗凝与纤维蛋白溶解
血液离开血管数分钟后,血液就由流动的溶胶状态变成不能流动的胶冻状凝块,这一过程称为血液凝固(blood coagulation)或血凝。在凝血过程中,血浆中的纤维蛋白源转变为不溶的血纤维。血纤维交织成网,将很多血细胞网罗在内,形成血凝块。血液凝固后1-2小时,血凝块又发生回缩,并释出淡黄色的液体,称为血清。血清与血浆的区别,在于前者缺乏纤维蛋白原和少量参与血凝的其他血浆蛋白质,但又增添了少量血凝时由血小板释放出来的物质。
血浆内具备了发生凝血的各种物质,所以将血液抽出放置于玻璃管内即可凝血。血浆内又有防止血液凝固的物质,称为抗凝物质(anticoagulant)。血液在血管内能保持流动,除其他原因外,抗凝物质起了重要的作用。血管内又存在一些物质可使血纤维再分解,这些物质构成纤维蛋白溶解系统(简称纤溶系统)(fibrinloytic system)。
在生理止血中,血凝、抗凝与纤维蛋白溶解相互配合,既有效地防止了失血,又保持了血管内血流畅通。
(一)血液凝固
凝血因子血浆与组织中直接参与凝血的物质,统称为凝血因子(bloodclotting factors),其中已按国际命名法用罗马数字编了号的有12种(表3-4)。此外,还有前激肽释放酶、高分子激肽原以及来自血小板的磷脂等直接参与凝血过程。除因子Ⅳ与磷脂外,其余已知的凝血因子都是蛋白质,而且因子Ⅱ、Ⅶ、Ⅸ、Ⅹ、Ⅺ、Ⅻ以及前激肽释放酶都是蛋白酶。这些蛋白酶都属于内切酶,即每一种酶只能水解某两种氨基酸所形成的肽键。因而不能将某一知肽链分解成很多氨基酸,而只能是对某一条肽链进行有限的水解。通常在血液中,因Ⅱ、Ⅶ、Ⅸ、Ⅹ、Ⅺ、Ⅻ都是无活性的酶原,必须通过有限水解在其肽链上一定部位切断或切下一个片段,以暴露或形成活性中心,这些因子才成为有活性的酶,这个过程称为激活。被激活的酶,称为这些因子的“活性型”,习惯上于该因子代号的右下角加一“a”字来表示。如凝血酶原被激活为凝血酶,即由因子Ⅱ变成因子Ⅱa。因子Ⅶ是以活性型存在于血液中的,但必须有因子Ⅲ(即组织凝血激酶)同时存在才能起作用,而在正常时因子Ⅲ只存在于血管外,所以通常因子Ⅶ在血流中也不起作用。
表3-4 按国际命名法编号的凝血因子
| 编 号 | 同 义 名 |
| 因子Ⅰ | 纤维蛋白原(fibrinogen) |
| 因子Ⅱ | 凝血酶原(prothrombin) |
| 因子Ⅲ | 组织凝血激素(tissue thromboplastin) |
| 因子Ⅳ | Ca2+ |
| 因子Ⅴ | 前加速素(proaccelerin) |
| 因子Ⅶ | 前转变素(proconvertin) |
| 因子Ⅷ | 抗血友病因子(antihemophilic factor,AHF) |
| 因子Ⅸ | 血浆凝血激酶(plasma thromboplastin component,PTC) |
| 因子Ⅹ | Stuart-Prower因子 |
| 因子Ⅺ | 血浆凝血激酶前质(plasma thromboplastin antecedent,PTA) |
| 因子Ⅻ | 接触因子(contact factor) |
| 因子ⅩⅢ | 纤维蛋白稳定因子(fibrin-stabilizing factor) |
凝血过程凝血过程基本上是一系列蛋白质有限水解的过程,凝血过程一旦开始,各个凝血因子便一个激活另一个,形成一个“瀑布”样的反应链直至血液凝固。凝血过程大体
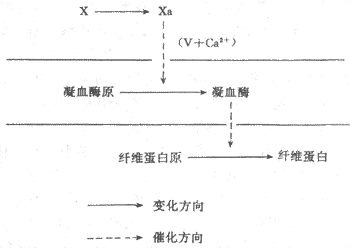
图3-4凝血过程的三个阶段简图
上可分为三个阶段(图3-4):即因子χ激活成χa;因子Ⅱ(凝血酶原)激活成Ⅱa(凝血酶);因子Ⅰ(纤维蛋白原)转变成Ⅰa(纤维蛋白)。
因子χ的激活可以通过两种途径。如果只是损伤血管内膜或抽出血液置于玻璃管内,完全依靠血浆内的凝血因子逐步使因子χ激活从而发生凝血的,称为径内源性激活途径(intrinsic route);如果是依靠血管外组织释放的因子Ⅲ来参与因子χ的激活的,称为外源性激活途径(extrinxic route),如创伤出血后发生凝血的情况。
1.内源性途径一般从因子Ⅻ的激活开始。血管内膜下组织,特别是胶原纤维,与因子Ⅻ接触,可使因子Ⅻ激活成Ⅻa。Ⅻa可激活前激肽释放酶使之成为激肽释放酶;后者反过来又能激活因子Ⅻ,这是一种正反馈,可使因子Ⅻa大量生成。Ⅻa又激活因子Ⅺ成为Ⅺa。由因子Ⅻ激活到Ⅺa形成为止的步骤,称为表面激活。表面激活过程还需有高分子激肽原*参与,但其作用机制尚不清楚。表面激活所形成的Ⅺa再激活因子Ⅸ生成Ⅸa,这一步需要有Ca2+(即因子Ⅳ)存在。Ⅸa再与因子Ⅷ和血小板3因子(PF3)及Ca2+组成因子Ⅷ复合物,即可激活因子Χ生成Χa。血小板3因子可能就是血小板膜上的磷脂,它的作用主要是提供一个磷脂的吸附表面。因子Ⅸa和因子χ分别通过Ca2+而同时连接于这个磷脂表面,这样,因子Ⅸa即可使因子χ发生有限水解而激活成为χa。但这一激活过程进行很缓慢,除非是有因子Ⅷ参与。因子Ⅷ本身不是蛋白酶,不能激活因子х,但能使Ⅸa激活因子χ的作用加快几百倍。所以因子Ⅷ虽是一种辅助因子,但是十分重要。遗传性缺乏因子Ⅷ将发生甲型血友病(hemophilia A),这时凝血过程非常慢,甚至微小的创伤也出血不止。先天性缺乏因子Ⅸ时,内源性途径激活因子χ的反应受阻,血液也就不易凝固,这种凝血缺陷称为B型血友病(hemophilia B)。
2.外源性途径由因子Ⅶ与因子Ⅲ组成复合物,在有Ca2+存在的情况下,激活因子χ生成χa。因子Ⅲ,原名组织凝血激酶,广泛存在于血管外组织中,但在脑、肺和胎盘组织中特别丰富。因子Ⅲ为磷脂蛋白质。Ca2+的作用就是将因子Ⅶ与因子χ都结合于因子Ⅲ所提供的磷脂上,以便因子Ⅶ催化因子χ的有限水解,形成χa。
Χa又与因子Ⅴ、PE3和Ca2+形成凝血酶原酶复合物,激活凝血酶原(因子Ⅱ)生成凝血酶(Ⅱa)。在凝血酶原酶复合物中的PF3也是提供磷脂表面,因子Χa和凝血酶原(因子Ⅱ)通过Ca2+而同时连接于磷脂表面,χa催化凝血酶原进行有限水解,成为凝血酶(Ⅱa)。因子Ⅴ也是辅助因子,它本身不是蛋白酶,不能催化凝血酶原的有限水解,但可使χa的作用增快几十倍。
因子χ与凝血酶原的激活,都是在PF3提供的磷脂表面上进行的,可以将这两个步骤总称为磷脂表面阶段。在这一阶段中,因子Ⅱ(凝血酶原)、因子Ⅶ、因子Ⅸ和因子χ,都必须通过Ca2+连接于磷脂表面。因此,在这些因子的分子上必须有能与Ca2+结合的部位。现已知,因子Ⅱ、Ⅶ、Ⅸ、х都是在肝中合成。这些因子在肝细胞的核糖体处合成肽链后,还需依靠维生素K的参与,使肽链上某些谷氨酸残基于γ位羧化成为γ-羧谷氨酸残基,构成这些因子的Ca2+结合部位。因此,缺陷维生素K,将出现出血倾向。
凝血酶(thrombin)有多方面的作用。它可以加速因子Ⅶ复合物与凝血酶原酶复合物的形成并增加其作用,这也是正反馈;它又能激活因子ⅩⅢ生成ⅩⅢa;但它的主要作用是催化纤维蛋白原的分解,使每一分子纤维蛋白原从N-端脱下四段小肽,转变成为纤维蛋白单体(fibrin monomer),然后互相连接,特别是在ⅩⅢa作用下形成牢固的纤维蛋白多聚体(fibrin polymers),即不溶于水的血纤维。上述凝血过程可见图3-5表示。
一般来说,通过外源性途径凝血较快,内源性途径较慢,但在实际情况中,单纯由一种途径引起凝血的情况不多。
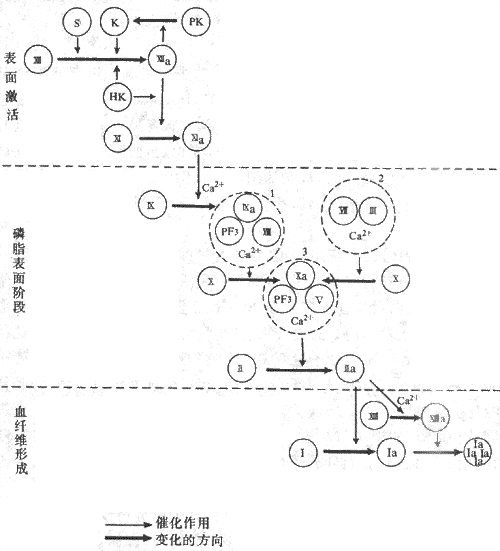
图3-5血液凝固过程示意图
S;血管内皮下组织 PF3:血小板3因子PK:前激肽释放酶 1:因子Ⅷ复合物
K:激肽释放酶 2:因子Ⅶ复合物 HK:高分子激肽原 3:凝血酶原酶复合物
在凝血的某些阶段,内源性途径与外源性途径之间存在着功能的交叉,也就是说,这两条途径之间具有某些“变通”的途径。例如,外源性的因子Ⅶa和Ⅲ可以形成复合物直接激活因子Ⅸ,从而部分代替了因子Ⅺ和Ⅻa的功能。这一机制得以解释为什么在因子Ⅸ缺乏时的出血倾向,较因子Ⅺ和Ⅻ缺乏时更为严重。另一方面,内源性因子Ⅻ的裂解产物和因子Ⅸa也能激活外源性的因子Ⅶ。
(二)抗凝系统的作用
正常人1ml血浆含凝血酶原约300单位,在凝血时通常可以全部激活。10ml血浆在凝血时生成的凝血酶就足以使全身血液凝固。但在生理止血时,凝血只限于某一小段血管,而且1ml血浆中出现的凝血酶活性很少超出8-10单位,说明正常人血浆中有很强的抗凝血酶活性。
现在已经查明,血浆中最重要的抗凝物质是抗凝血酶Ⅲ(antithrombinⅢ)和肝素,它们的作用约占血浆全部抗凝血酶活性的75%。抗凝血酶Ⅲ是血浆中一种丝氨酸蛋白酶抑制物(serine protease inhibitor)。因子 Ⅱa、Ⅶ、Ⅸa、χa、Ⅻa的活性中心均含有丝氨酸残基,都属于丝氨酸蛋白酶(serine protease)。抗凝血酶Ⅲ分子上的精氨酸残基,可以与这些酶活性中心的丝氨酸残基结合,这样就“封闭”了这些酶的活性中心而使之失活。在血液中,每一分子抗凝血酶Ⅲ,可以与一分子凝血酶结合形成复合物,从而使凝血酶失活。
肝素是一种酸性粘多糖,主要由肥大细胞和嗜碱性粒细胞产生,存在于大多数组织中,在肝、肺、心和肌组织中更为丰富。
肝素在体内和体外都具有抗凝作用,肝素抗凝的主要机制在于它能结合血浆中的一些抗凝蛋白,如抗凝血酶Ⅲ和肝素辅助因子Ⅱ(heparincofactorⅡ)等,使这些抗凝蛋白的活性大为增强。当肝素与抗凝血酶Ⅱ的某一个ε-氨基赖氨酸残基结合,则抗凝血酶Ⅲ与凝血酶的亲和力可增强100倍,使两者结合得更快,更稳定,使凝血酶立即失活。当肝素与肝素辅助因子Ⅱ结合而激活后者时,被激活的肝素辅助因子Ⅱ特异性地与凝血酶结合成复合物,从而使凝血酶失活,在肝素的激活作用下,肝素辅助因子灭活凝血酶的速度可以加快约1000倍。
肝素还可以作用血管内皮细胞,使之释放凝血抑制物和纤溶酶原激活物,从而增强对凝血的抑制和纤维蛋白的溶解。此外,肝素能激活血浆中的脂酶,加速血浆中乳糜微粒的清除,因而减轻脂蛋白对血管内皮的损伤,有助于防止与血脂有关的血栓形成。
天然肝素是一种分子量不均一的混合物,分子量为3000-57000不等。这种不均一是生物合成过程有差异所致。不同分子量肝素的生物作用也不完全相同。一般将分子量在7000以下肝素称为低分子量肝素。低分子量肝素只与抗凝血酶Ⅱ结合,而分子量较大的肝素除了能与抗凝血酶Ⅲ结合外,还能与血小板结合,结果不仅抑制血小板表面凝血酶的形成,而且抑制血小板的聚集与释放。由于分子量较大的肝素抗凝作用的环节较多,作用较为复杂,易引起出血倾向,而低分子时肝素具有半衰期较长,抗凝效果好和引起出血倾向少等优点,因而更适于作为外源性抗凝剂。
从化学本质看,凝血过程是一系列酶促反应链,其中主链是一系列丝氨酸蛋白酶的作用。组成抗凝系统的一类物质是血浆中存在的多种丝氨酸蛋白酶抑制物。上述抗凝血酶Ⅲ是其中最为重要的一种。属于丝氨酸蛋白酶抑制物的抗凝物质还有能抑制补体第1成分和因子Ⅻa、Ⅺa、的C1抑制物(C1inhibitor),广谱的蛋白酶抑制物a2-巨球蛋白(a2-macroglobulin)等。抗凝系统中的另一类物质是辅因子抑制物,这类抑制物通过对凝血辅因子如因子Ⅴ和Ⅷ活性的抑制而实现抗凝作用。下述的蛋白质C和凝血酶调制素都是属于这类抗凝物质。
蛋白质 C(proteinC)是近年来引起注意的另一种具有抗凝作用的血浆蛋白,分子量为62000,它由肝合成,并有赖于维生素K的存在。蛋白质C以酶原形式存在于血浆中,蛋白质C在凝血酶的作用下发生有限的酶解过程,从分子上裂解下一个小肽后即具有活性。激活的蛋白质C与血管内皮表面存在的辅因子凝血酶调制素(thrombomodulin)结合成复合物,在Ca2+存在的条件下这种复合物使蛋白质C的激活过程大大加快。激活的蛋白质C具有多方面的抗凝血、抗血栓功能,主要的作用包括:①灭活凝血因子Ⅴ和Ⅷ。这种灭活也是一种酶解过程,即是把因子Ⅴ和Ⅷ的重链进行水解,使他们与磷脂的结合力降低。这种灭活反应需要有Ca2+的存在,反应的速度是很快的;②限制因子Χa与血小板结合。存在于血小板表面的因子Ⅴa是因子Χa的受体。当因子Χa与这种受体结合后,可使因子Χa的活性大为增强。由于激活的蛋白质C能使因子Ⅴa灭活,使因子Χa与血小板的结合受到阻碍,结果可使因子Χa激活凝血酶原的作用大为减弱;③增强纤维蛋白的溶解。激活的蛋白质C能刺激溶酶原激活物的释放,从而增强纤溶活性。激活的蛋白质C的这一作用只有在内皮细胞存在的情况下才能实现。维生素K缺乏或患肝病可使蛋白质C的合成减少;某些病理情况造成血管内皮广泛损伤时使凝血酶调制减少,这种减少转而使蛋白质C的激活受阻。不论是蛋白质C减少或活化受阻都增加了形成血栓的倾向。
体外延缓或阻止血液凝固的因素:①降低温度,当反应系统的温度降低至10℃以下时,很多参与凝血过程的酶的活性下降,因些可延缓血液凝固,但不能完全阻止凝血的发生;②光滑的表面,也称不湿表面,可减少血小板的聚集和解体,减弱对凝血过程的触发,因而延缓了凝血酶的形成。例如,将血液盛放在内表面涂有硅胶或石蜡的容器内,即可延缓血凝,③去Ca2+,由于血液凝固的多个环节中都需要Ca2+的参加,因此如在体外向血液中加入某些能与钙结合形成不易解离但可溶解的络合物,从而减少了血浆中的Ca2+,防止了血液凝固。由于少量枸橼酸钠进入血液循环不致产生毒性,因此常用它作抗凝剂来处理输血用的血液。此外,实验室中可使用草酸铵、草酸钾和螯合剂乙二胺四乙酸(ECTA)作抗凝剂,它们能与Ca2+结合不易溶解的复合物。但它们对机体有害,因而不能进入体内。
(三)纤维蛋白溶解
在生理止血过程中,小血管内的血凝块常可成为血栓,填塞了这一段血管。出血停止、血管创伤愈合后,构成血栓的血纤维可逐渐溶解,先形成一些穿过血栓的通道,最后可以达到基本畅通。血纤维溶解的过程,称为纤维蛋白溶解(简称纤溶)。
纤维蛋白溶解(纤溶)系统包括四种成分,即纤维蛋白溶解酶原(plasminogen)(纤溶酶原,血浆素原)、纤维蛋白溶解酶(plasmin)(纤溶酶,血浆素)、纤溶原激活物与纤溶抑制物。纤溶的基本过程可分两个阶段,即纤溶酶原的激活与纤维蛋白(或纤维蛋白原)的降解(图3-6)。
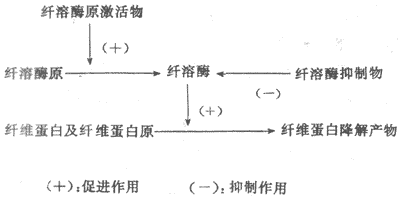
图3-6纤维蛋白溶解系统
1.纤溶酶原激活纤溶酶原很可能是在肝、骨髓、嗜酸性粒细胞与肾中合成的;在正常成年人每100ml血浆中约含10-20mg纤溶酶原,婴儿较少,妇女晚期妊娠时增多。
纤溶酶原激活物分布广而种类多,主要有三类:第一类为血管激活物,在小血管内皮细胞中合成后释放于血中,以维持血浆内激活物浓度于基本水平。血管内出现血纤维凝块时,可使内皮细胞释放大量激活物。所释放的激活物大都吸附于血纤维凝块上,进入血流的很少。肌肉运动、静脉阻断、儿茶酚胺与组胺等也可使血管内皮细胞合成和释放的激活物增多。第二类为组织激活物,存在于很多组织中,主要是在组织修复、伤口愈合等情况下,在血管外促进纤溶。肾合成与分泌的尿激酶就属于这一类激活物,活性很强,有助于防止肾小管中纤维蛋白沉着。第三类为依赖于因子Ⅻ的激活物,例如前激肽释放酶被Ⅻa激活后,所生成的激肽释放酶即可激活纤溶酶原。这一类激活物可能使血凝与纤溶互相配合并保持平衡。
血浆中的激活物的半衰期约13分钟,通常迅速被肝清除。
纤溶酶原的激活也是有限水解的过程,在激活物的作用下,脱下一段肽链成为纤溶酶。
2.纤维蛋白(与纤维蛋白原)的降解 纤溶酶和凝血酶一样,也是蛋白酶,但是它对纤维蛋白原的作用与凝血酶不同。凝血酶只是使纤维蛋白原从其中两对肽链的N-端各脱下一个小肽,使纤维蛋白原转变成纤维蛋白。纤溶酶却是水解肽链上各单位的赖氨酸-精氨酸键,从而逐步将整个纤维蛋白或纤维蛋白原分割成很多可溶的小肽,总称为纤维蛋白降解产物。纤维蛋白降解产物一般不能再出现凝固,而且其中一部分有抗血凝的作用。
纤溶酶是血浆中活性最强的蛋白酶,但特异性较小,可以水解凝血酶、因子Ⅴ、因子Ⅷ、激活因子Ⅻa;促使血小板聚集和释放5-羟色胺、ADP等;还能激活血浆中的补体系统;但它的主要作用是水解纤维蛋白原和纤维蛋白。血管内出现血栓时,纤溶主要局限于血栓,这可能是由于血浆中有大量抗纤溶物质(即抑制物)存在,而血栓中的纤维蛋白却可吸附或结合较多的激活物所致。正常情况下,血管内膜表面经常有低水平的纤溶活动,很可能血管内也经常有低水平的凝血过程,两者处于平衡状态。
3.抑制物及其作用血液中存在的纤溶抑制物主要是抗纤溶酶(antiplasmin) ,但其特异性不大,例如,a2-巨球蛋白能普遍抑制各种内切酶,包括纤溶酶、胰蛋白酶、凝血酶、激肽释放酶等。每一分子a2-巨球蛋白可结合一分子纤溶酶,然后迅速被吞噬细胞清除。血浆中a1-抗胰蛋白酶也对纤溶酶有抑制作用,但作用较慢,然而它分子量小,可渗出血管,控制血管外纤溶活动。看来这些抑制物的作用,是广泛控制在血凝与纤溶两个过程中起作用的一些酶类。这对于将血凝与纤溶局限于创伤部位,有重要意义。
(四)表面激活与血液的其他防卫功能
血管损伤后暴露出内膜下组织,通过表面激活使因子Ⅻ激活成因子Ⅻa,因子Ⅻa 又激活肽释放酶成为激肽释放酶,而激肽释放酶又可激活因子Ⅻ,从而形成一个正反馈环,可形成足够的Ⅻa 和激肽释放酶。这样,不但同时激活了血凝和纤溶系统,也激活了补体系统和激肽系统(图3-7)。补体激活的一些产物和激肽都是作用很强的趋化因子,能吸收吞噬细胞到受损伤的部位,产生非特异性免疫反应;这样使生理止血功能与免疫功能相配合,有效的保护机体,减少创伤带来的损害。
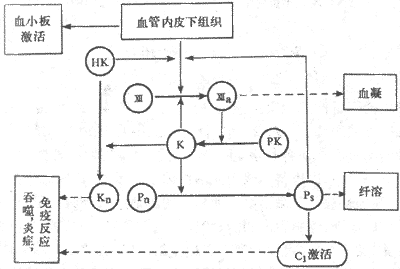
图3-7表面激活也血液各种防卫功能关系示意图
PK:前激肽释放酶 Pn:纤溶酶原 K:激肽释放酶 Pn:纤溶酶
HK:高分子激肽原 CI:补体 Kn:激肽 Ⅻ与Ⅻa,因子Ⅻ及其激活型
二、血小板的止血功能
因血管创伤而失血时,血小板在生理止血过程中的功能活动大致可以分为两段,第一段主要是创伤发生后,血小板迅速粘附于创伤处,并聚集成团,形成较松软的止血栓子;第二段主要是促进血凝并形成坚实的止血栓子。
(一)血小板粘附与聚集
止血中较松软的血小板止血栓子的形成,要经过血小板粘附与聚集两个过程。
血管损伤后,流经此血管的血小板被血管内皮下组织表面激活,立即粘附于损伤处暴露的胶原纤维上。参与血小板粘附过程的主要因素包括:血小板膜糖蛋白I(GPI)、vonWillebrand因子(vW因子)和内皮下组织中的胶原。当血小板缺乏GPI或胶原纤维变性时,血小板粘附(thrombocyte adhesion)功能便受损。发生血小板粘附过程的可能机制是vW因子再与血小板膜上的特异受体结合。此外,血小板膜上的糖苷移换酶活性和胶原蛋白分子的构型与粘附也有着密切关系。
粘附主要是一种表面现象,粘附一旦发生了,血小板的聚集过程(thrombocyteaggregation)也随即发生。聚集是指一些血小板相互粘连在一起的过程。聚集开始时,血小板由圆盘形变成球形,并伸出一些貌似小刺的伪足;同时血小板脱粒,即原来贮存于致密颗粒内的ADP、5-羟色胺等活性物质被释放。ADP释放和某些前列腺素的生成,对聚集的引起十分重要。
1.ADP的作用 在体外实验中看到,ADP是使血小板聚集最重要的物质,特别是从血小板释放出来的这种内源性ADP尤其重要。在血小板悬液中加入小量ADP(浓度在0.9μmol/L以下),能迅速引起血小板聚集,但很快又解聚;若加入中等剂量的ADP(1.0μmol/L左右),则在第一聚集时相结束和解聚后不久,又出现第二个不可逆的聚集时相,这是由于血小板释放的内源性ADP所引起的;若是加入大量ADP,则迅速引起不可逆的聚集,即直接进入聚集的第二时相.以不同剂量的凝血酶加入血小板悬液,也可使血小板发生聚集;而且与ADP相似,随着加入剂量的逐渐增加,可看到从只有第一时相可逆性聚集,到出现两个时相的聚集,再到直接进入第二时相的聚集.因为,用腺苷阻断内源性ADP的释放或用腺苷三磷酸双磷酸酶(apyrase)以破坏ADP,均可抑制凝血酶引起的聚集,说明凝血酶的作用可能是由于凝血酶与血小板细胞膜上的凝血酶受体结合后,引起内源性ADP释放所引起的。加入胶原也可引进悬液中的血小板聚集,然而只有第二时相的不可逆聚集,一般认为这也是由于胶原引起内源性的ADP释放所致。
一般能引起血小板聚集的物质均可使血小板内cAMP减少,而抑制血小板聚集的则使cAMP增多。因而目前认为,可能是cAMP减少引起血小板内Ca2+增加,促使内源性ADP释放。
ADP引起血小板聚集,还必须有Ca2+和纤维蛋白原存在,而且要消耗能量。将血小板悬浮于缺乏葡萄糖的溶液中数小时,或用药物阻断或减弱血小板产生ATP的代谢过程,均将抑制血小板的聚集。ADP也不能使洗净了的血小板聚集,除非加入纤维蛋白原;但凝血酶和胶原可使洗净了的血小板聚集。因为在这种情况下,可使血小板a 颗粒内的纤维蛋白原释放。
ADP是通过血小板膜上的ADP受体引起聚集的。目前认为,血小板膜上有表面ATP酶,这是防止血小板相互粘聚所必需的,而ADP可抑制表面ATP酶的活性;ADP还可使血小板暴露出磷脂表面,因而可以通过Ca2+“搭桥”而互相粘聚。
2.血小板前列腺素类物质的作用血小板质膜的磷脂中含有花生四烯酸,血小板细胞内有磷脂酸A2。在血小板被表面激活时,磷脂酶A2也被激活。在磷脂酶A2的催化作用下,花生四烯酸从质膜的磷脂中分离出来。花生四烯酸在血小板的环氧化酶作用下,产生前列腺素G2和H2(PGG2、PGH2)。PGG2和PGH2都是环内过氧化物,有很强的引起血小板聚集的作用。但是PGG2和PGH2都很不稳定,可以直接生成小量PGE2和PGF2。PGH2可以在血栓素合成酶的催化作用下,形成大量血栓素A2(thromboxane A2,TXA2)。TXA2使血小板内cAMP减少,因而有很强的聚集血小板的作用,也有很强的收缩血管的作用。TXA2也不稳定迅速转变成无活性的血栓素B2(TXB2)。咪唑(imidazole)可抑制血栓素合成酶,所以有防止血小板聚集的作用。此外,正常血管壁内皮细胞中有前列腺环素合成酶,可以催化血小板生成的PGH2生成前列腺环素(prostacyclin,PGI2)。PGI2可使血小板内cAMP增多,因而有很强抑制血小板聚集的作用,也有很强的抑制血管收缩的作用。PGI2也很不稳定,迅速变成无活性的6-酮-PGF1a。关于由花生四烯酸衍变成TXA2与PGI2的过程可参看图3-8。
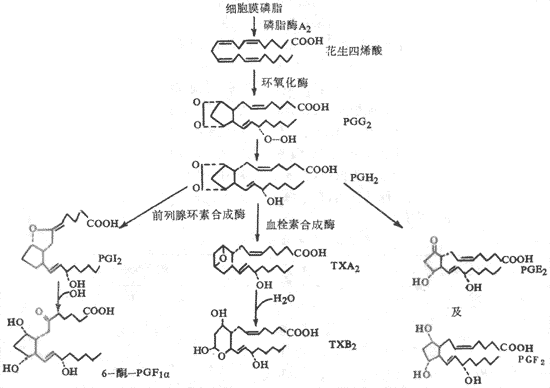
图3-8血小板前列腺素与血栓素的合成
在发现TXA2和PGI2之后,曾设想在正常情况下可能是血管壁的PGI2与血小板的TXA2之间保持了平衡,因而使血小板不致聚集。可以设想,血管损伤暴露内皮下组织时,一方面激活血小板和激活内源性凝血途径,损坏的血管组织释放凝血因子Ⅲ又激活外源性凝血途径,于是在此局部迅速形成凝血酶;另一方面血管损伤使局部血管壁PGI2减少。这样,由此血管通过的血小板即粘附于损伤处的胶原纤维上,随即血小板也发生变形、聚集,并激活磷脂酶A2,导致合成TXA2,TXA2可使血小板内cAMP减少而游离Ca2+增多,以致血小板脱粒释放内源性ADP,又使更多的血小板聚集,迅速形成松软的止血栓子。
(二)血小板与凝血
血小板对于血液凝固有重要的促进作用,如将血液置于管壁涂一薄层硅胶的玻璃管中,使血小板不易解体,虽然未加入任何抗凝剂,血液可保持液态达72小时以上;若加入血小板匀浆则立即发生凝血。这说明血小板破裂后的产物对于凝血过程有很强的促进作用。
血小板表面的质膜结合有多种凝血因子,如纤维蛋白原、因子Ⅴ、因子Ⅺ、因子ⅩⅢ等。a-颗粒中也含有纤维蛋白原、因子因子ⅩⅢ和一些血小板因子(PE),其中PF2和PF3都是促进血凝的。PF4可中和肝素,PF6则抑制纤溶。当血小板经表面激活后,它能加速凝血因子Ⅻ和Ⅺ的表面激活过程。血小板所提供的磷脂表面(PF3),据估计可使凝血酶原的激活加快两万倍。因子Ⅹa和因子Ⅴ连接于此磷脂表面后,还可以免受抗凝血酶Ⅲ和肝素对它们的抑制作用。
当血小板聚集形成止血栓时,凝血过程已在此局部进行,血小板已暴露大量磷脂表面,为因子Ⅹ和凝血酶原的激活提供了极为有利的条件。血小板聚集后,其α颗粒中的各种血小板因子释放出来,促进血纤维的形成和增多,并网罗其它血细胞形成凝块。因而血小板虽逐渐解体,止血栓子仍可增大。血凝块中留下的血小板有伪足伸入血纤维网中,这些血小板中的收缩蛋白收缩,使血凝块回缩,挤压出其中的血清而成为坚实的止血栓,牢牢地封住血管缺口。
在表面激活血小板和血凝系统时,同时也激活了纤溶系统。血小板内所含的纤溶酶及其激活物将释放出来。血纤维和血小板释放的5-羟色胺等,也能使内皮细胞释放激活物。但是由于血小板解体,同时释放出PF6和另一些抑制蛋白酶的物质,所以在形成血栓时,不致受到纤溶活动的干扰。
第四节 血型与输血原则
一、血型与红细胞凝集
若将血型不兼容的两个人的血滴放在玻片上混合,其中的红细胞即聚集成簇,这种相容称为凝集(agglutination)。红细胞的凝集有时还伴有溶血。当血型(bolld group)不相容的血液输入循环血液中时,在血管内可发生同样的情况,此凝集成簇的红细胞可以堵塞毛细血管,溶血将损害肾小管,同时常伴发过敏反应,其结果可危及生命。
造成红细胞凝集的机制是抗原-抗体反应。凝集原的特异性完全取决于镶嵌入红细胞膜上的一些特异糖蛋白,在凝集反应中糖蛋白起着抗原的作用,因而称它们为凝集原(agglutinogen)。能与红细胞膜上的凝集原起反应的特异抗体则称为凝集素(agglutinin)。凝集素是由γ-球蛋白构成的,它们溶解在血浆中。发生抗原-抗体反应时,由于每个抗体上具有10个左右与抗原结合的部位,抗体在若干个带有相应抗原的红细胞之间形成桥梁,因而使它们聚集成簇。
人血液中,在红细胞上都具有一套特异的凝集原。目前在红细胞上已确定有许多种不同的抗原,大约有30种抗原能引发相当剧烈的机体反应。血型是指红细胞上特异抗原的类型。表3-5中列出了ABO、Rh、MNSs、P等9个最重要的血型系统和它们所具有的特异抗体。动用现代免疫学手段可以在红细胞膜上鉴别出约400种不同特征的抗原。如果只将其中已经分类的抗原型作为依据,就有3亿种不同的可能组合,可见血型抗原最为重要的血型是ABO系统和Rh系统。
表3-5 重要血型及其特异抗体
| 血型系统 | 抗体 | 溶血性输血反应 |
| ABO | 抗A | 有 |
| 抗B | 有 | |
| 抗A1 | 很少 | |
| 抗H | 无 | |
| Rh | 抗C | 有 |
| 抗c | 有 | |
| 抗Cw | 有 | |
| 抗D | 有 | |
| 抗E | 有 | |
| 抗e | 有 | |
| MNSs | 抗M,N,S,s | 很少 |
| P | 抗P1 | 无 |
| Lutheran | 抗Lub | 有 |
| Kell | 抗K | 有 |
| Lewis | 抗Lea,b | 有 |
| Duffy | 抗Fya | 有 |
| Kidd | 抗JKa | 有 |
虽然通常所说的血型是指红细胞的血型,但是存在于红细胞上的血型抗原也存在于白细胞、血小板和一般组织细胞上,引外在白细胞和血小板上还存在它们本身特有的抗原。
二、红细胞血型
1901年 Landsteiner发现了第一个血型系统,即ABO血型系统,从此为人类揭开了血型的奥秘,并使输血成为安全度较大的临床治疗手段。
(一)ABO血型系统
1.ABO血型的分型及其物质基础ABO血型是根据红细胞膜上存在的凝集原A与凝集原B的情况而将血液分为4型。凡红细胞只含A凝集原的,即称A型;如存在B凝集原的,称为B型;若A与B两种凝集原都有的称为AB型;这两种凝集原都没有的,则称为O型。不同血型的人的血清中各含有不同的凝集素,即不含有对抗内他自身红细胞凝集原的凝集素。在A型人的血清中,只含有抗B凝集素;B型人的血清中,只含有抗A凝集素;AB型人的血清中没有抗A和抗B凝集素;而O型人的血清中则含有抗A和抗B凝集素(表3-6)。后来进一步发现4种血型的红细胞上都含有H抗原,O型的红细胞上也含有H抗原。H抗原是形成A、B抗原的结构基础,但是H物质的抗原性很弱,因此血清中一般都没有抗H抗体。利用抗血清作细致的检测可以发现,A型还可再区分为A1、和A2亚型。在A1亚型红细胞上含有A和A1抗原,而A2型红细胞上仅含有A抗原。相应的在A1型血清中只有抗B凝集素,而A2型血清中除抗B凝集素之外,还含有抗A1凝集素。因此当将A1型的血液输给A2型的人时,血清中的抗A1凝集素可能与A1型的人红细胞上的A1抗原结合产生凝集反应。据调查,我国汉族人中A2型和A2B型分别不超过A型和AB型人群的1%,即使如此,在测定血型和输血时都应注意到A亚型的存在。
表3-6 ABO血型系统中的凝集原和凝集素
| 血型 | 凝集原 | 凝集素 |
| A型 | A | 抗B |
| B型 | B | 抗A |
| AB型 | A+B | 无 |
| O型 | 无 | 抗A+抗B |
上述ABO系统各种血型抗原的特异必理决定于糖蛋白上所含的糖链。这些糖链都是由少数糖基所组成的寡糖链(oligosaccharide)。这些寡糖链都暴露在红细胞的表面,图3-9示意了ABO系统中H、A和B抗原的寡糖链结构差异。
图3-9 ABH抗原物质化学结构
2.血型的遗传学特征血型是先天遗传的。出现在某一染色体的同一位置上的不同基因,称为等们基因(allele)。ABOA(H)系统中控制A、B、H抗原生成的基因即为等位基因。在染色体二倍体上只可能出现上述三个等们基因中的两个,其中一个来自父体,另一个来自母体,这两个等位基因就决定了子代血型的基因型(genotype)。这两种基因型首先决定了转糖酶的氨基酸组成和顺序,也即决定了生成的转糖酶的种类,后者转而决定表现血型抗原特异性的寡糖链的组成,也即这个人的血型表型(phenotype)。表3-7上显示了ABO系统中决定每种血型表型的可能基因型。从表上可以看出,A基因和B基因是显性基因,O基因则为隐性基因。因此,红细胞上表型O只可能来自两个O基因,而表型A或b 由于可能分别来自AO和BO基因型,因而,A型或B型的父母完全可能生下O型的子女。知道了血型的遗传规律,就可以从子女的血型表型来推断亲子关系。例如,AB型的人决不可能是O型子女的父亲。但必须注意的是,法医学上需要依据血型表型来判断亲子关系时,只能作为否定的参考依据,而不能据此作出肯定的的判断。由于血细胞上有许多种血型,测定血型的种类愈多,那么作出否定性判断的可靠性也愈高。
表3-7 ABO(H) 血型系统中的结构
| 表现型 | 遗传型 | 红细胞抗原 | 血清中天然抗体 |
| A | A1A1 | A+A1 | 抗B |
| A1 | |||
| A2 | A2A2 | A+H | 抗B,10%的人有抗A1 |
| A2O | |||
| B | BB | B | 抗A |
| BO | |||
| AB | AB | A+A1+B | --- |
| A2B | A2B | A+B+H | 25%的人有抗A1 |
| O | OO | H | 抗A及抗B |
新生儿的血液中还不具有ABO系统的抗体;在出生后的第一年中这种抗体才逐渐出现在血浆中,可以对抗自已血细胞上所没有的抗原。这类天然抗体多属IgM,其产生的原因尚未完全阐明,据推测,由肠道细菌所释放的物质或某些食物成分进入体内能够刺激血型抗体的产生。因为某些肠道具有与红细胞同样的抗原决定簇。
血型抗原在人群中的分布,在不同地域不同民族中是有差异的。以研究较多的ABO系统为例,在中欧地区人群中,40%以上为A型,稍低于40%为O型,10%左右为B型,6%左右为Ab 型;而在美洲土著民族中则90%属O型。在我国各族人民中ABO血型的分布也不尽相同。详见表3-8以了解各地域、各民族的血型分布规律,将有助于人类学研究各民族的来源和相互关系。
| 调查对象 | 调查人数(个) | A | B | AB | O | ||||
| 人数(个) | % | 人数(个) | % | 人数(个) | % | 人数(个) | % | ||
| 汉族 | 40,980 | 12,831 | 31.31 | 11,501 | 28.06 | 4002 | 9.77 | 12,646 | 30.86 |
| 维吾尔族 | 1,513 | 441 | 29.22 | 483 | 31.92 | 172 | 11.36 | 416 | 27.50 |
| 壮族 | 1,487 | 316 | 21.25 | 410 | 25.57 | 58 | 3.90 | 703 | 47.28 |
| 回族 | 1,355 | 369 | 27.23 | 384 | 28.34 | 115 | 8.48 | 487 | 35.94 |
| 哈萨克族 | 885 | 202 | 22.82 | 264 | 29.83 | 83 | 9.38 | 336 | 37.97 |
| 锡伯族 | 344 | 86 | 25.00 | 138 | 40.12 | 36 | 10.46 | 84 | 24.42 |
| 乌孜别克族 | 129 | 33 | 25.58 | 50 | 38.76 | 13 | 10.08 | 33 | 25.58 |
| 柯尔克孜族 | 124 | 23 | 18.54 | 49 | 39.52 | 9 | 7.26 | 43 | 34.68 |
| 塔塔尔族 | 37 | 15 | 40.54 | 13 | 35.14 | 1 | 2.70 | 8 | 21.62 |
| 彝族 | 1007 | 288 | 28.60 | 303 | 30.09 | 82 | 8.14 | 334 | 33.17 |
| 白族 | 500 | 170 | 34.00 | 117 | 23.40 | 56 | 11.20 | 157 | 31.40 |
| 傣族 | 507 | 112 | 22.08 | 150 | 29.59 | 40 | 7.89 | 205 | 40.44 |
| 景颇族 | 201 | 70 | 34.83 | 41 | 20.39 | 14 | 6.97 | 76 | 37.81 |
| 佤族 | 520 | 200 | 38.46 | 112 | 21.54 | 73 | 14.04 | 135 | 25.96 |
| 土家族 | 960 | 362 | 37.71 | 219 | 22.81 | 61 | 7.19 | 310 | 32.29 |
上海生物制品研究所血型组:<<血型与血库>>第9页。上海人民出版社第1版1977年 湖南医学院生理教研组调查报告
3.ABO血型的检测正确测定血型是保证输血安全的基础。在一般输血中只有ABO系统的血型相合才能考虑输血。测定ABO系统的方法是:在玻片上分别滴上一滴抗B、一滴抗A和一滴抗A-抗B血清,在每一滴血清上再加一滴红细胞悬浮液,轻轻摇动,使红细胞和血清混匀,观察有无凝集现象(图3-10)。
图3-10 ABO血型的测定
(二)Rh血型系统
1.Rh血型系统的发现和在人群中的分布在寻找新血型物质的探索中,当把恒河猴(Rhesus monkey)的红细胞重复注射入家兔体内,引起家兔产生免疫反应,此时在家兔血清中产生抗恒河猴红细胞的抗体(凝集素)。再用含这种抗体的血清与人的红细胞混合,发现在白种人中,约有85%的人其红细胞可被这种血清凝集,表明这些人的红细胞上具有与恒河猴同样的抗原,故称为Rh阳性血型;另有约15%的人的红细胞不被这种血清凝集,称为R h阴性血型,这一血型系统即称为Rh血型。在我国各族人中,汉族和其它大部分民族的人,属Rh阳性的约占99%,Rh阴性的人只占1%左右。但是在另一些少数民族中,Rh阴性的人较多,如苗族为12.3%,塔塔尔族为15.8%。
2.Rh血型系统的基因型及其表达利用血清试验提示了人类红细胞上的Rh血型系统包括着5种不同的抗原,分别称为C、c、D、E、e。从理论上推断,有3对等位基因Cc、Dd 、Ee控制着6个抗原。但实际上未发现单一的抗d血清,因而认为d是“静止基因”,在红细胞表面不表达d抗原。在5个抗原中,D抗原的抗原性最强。因此通常将红细胞上含有D抗原的,即称为Rh阳性;而红细胞上缺乏D抗原的,称为Rh阴性。
3.Rh血型的特点及其在医学实践中的意义前述ABO血型时曾指出,从出生几个月之后在人血清中一直存在着ABO系统的凝集素,即天然抗体。但在人血清中不存在抗Rh的天然抗,只有当Rh阴性的人,接受Rh阳性的血液后,通过体液性免疫才产生出抗Rh的抗体来。这样,第一次输血后一般不产生明显的反应,但在第二次,或多次再输入Rh阳性血液时即可发生抗原-抗体反应,输入的Rh阳性红细胞即被凝集。
Rh系统与ABO系统比较时的另一个不同点是抗体的特征。ABO系统的抗体一般是完全抗体IgM。而Rh系统的抗体主要是不完全抗体IgG,后者分子较能透过胎盘。因此,当阴性的母亲怀有阳性的胎儿时,阳性胎儿的小时红细胞或D抗原可以进入母体,通过免疫反应,在母体的血液中产生免疫抗体,主要是抗D抗体。这种抗体可以透过胎盘进入胎儿的血液,可使胎儿的红细胞民生凝集和溶解,造成新生儿溶血性贫血,严重时可致胎儿死亡。但一般只有在分娩时才有较大量的胎儿红细胞进入母体,而母体血液中的抗体浓度是缓慢增加的,一般需要数月的时间,因此,第一次妊娠常不产生严重反应。如果Rh阴性母亲再次怀有Rh阳性胎儿时,此时,母体血液中高浓度的Rh抗体将会透过胎盘,破坏大量胎儿红细胞。
三、白细胞与血小板血型
白细胞与血小板上与有A、B、H、MN、P等红细胞抗原,此外还有它们所特有的抗原,这些抗原具有临床意义,特别是组织兼容性抗原对选择器官组织的移植和血液成分的输注的合适供应者(donor)有重要意义。白细胞与血小板的抗原可使受者(recipient)产生免疫反应,这时,输血可引起发热反应,移植的器官组织受破坏的速度加快,在体内的生存期缩短。
人白细胞抗原( human leukocyte antigen,HLA)是人类白细胞上最强的同种抗原。HLA系统由数量众多的抗原所组成,是一个极为复杂的抗原系统,控制这些抗原的基因位于第6号染色体的短臂上,在这一部位的染色体上分布着A、B、C、D4个基因位点,它们控制着7组HLA抗原即:HLA-A、HLA-B、HLA-C、HLA-D、HLA-DR、HLA-DQ和HLA-DP,每一组又包括许多种抗原。例如HLA-A组有20多种抗原,HLA-B组有50种以上不同的抗原,HLA-C组则有11种抗原。根据抗原发现的顺序,又对抗原作编号,如HLA-A1、A2……A19等。它们的数量随着新抗原的认定还在继续扩大。
HLA是镶嵌在细胞膜上的糖蛋白,根据HLA的结构和分布特征,将它们分成两类:即I类和Ⅱ类抗原,HLA-A、B、C属I类抗原,分子量为56000,由一条重肽链和一条轻肽链所组成,它们的抗原特异性是由重肽链上氨基酸列所决定的。这类抗原总称 为I类抗原;它们除了分布在白细胞和血小板上,还广泛分布在各种正常组织器官和肿瘤组织的有核细胞膜上。其余的HLA抗原即D、DR、DP和DQ属于Ⅱ类抗原,分子量为63000,它们只分布在B细胞、巨噬细胞、单核细胞和内皮细胞上。
HLA系统除了在医学上与器官移植、植皮、骨髓移植和输轿有密切的关系外,还可应用于亲子鉴定和人类学研究。在应用于亲子鉴定时,由于HLA的数目极多,HLa –A和HLA-B相结合的形式极多,要有相同表现型的机会极少。例如,15个HLA-A和20个HLA-B能够产生20246种表现型。因此,在作亲子关系的否定判定时,HLA测定可过到90%以上的可靠性。各种HLA出现的频率具有明显的种族差异。例如,高加索人种中,HLA-A-A30和B42抗原的出现频率较其它人种为少,而在北美印地安人种,HLA-B51抗原出现频率较高。因此,HLA系统是人类学研究的一个重要指标。
血小板还有一些特有的抗原,如PI、Zw、Ko等系统,它们与红细胞或白细胞上的同种抗原没有关系。这些抗原可因输血和妊娠而产生免疫抗体。约有50%的病人在长期重复输注血小板后,在血清中可出现抗输入血小板的抗体。它们可引起发热反应,使输入的细胞缩短生存期,还以掩盖抗红细胞的特异抗体的存在,因而影响血型测定的正确性。当妊娠母亲血清中出现抗血小板抗体时,可导致新生儿血小板减少症。
四、输血的原则
输血已经成为治疗某些疾病、抢救伤员生命和保证一些手术得以顺利进行的重要手段。但是,由于输血发生差错,造成病人严惩损害,甚至死亡的事故并不鲜见。美国的统计资料报道,在1976年至1985年的10年间。美国共发生输血死亡事故159例,其中由于ABO系统的错误为137例,占86%。为了保证输血的安全性和提高输血的效果,必须注意遵守输血的原则。
随着医学和科学技术的进步,输血疗法已经从原来的单纯输全血,发展为输全血和成分输血(transfusion or blood components)。成分输血,就是把人血中的各种有效成分,如红细胞、粒细胞、血小板和血浆分别制备成高纯度或高浓度的制品再输入。这样既能提高疗效,减少不良反应,又能节约血源。
在准备输血时,首先必须保证供血者与受血者的ABO血型相合,因为这一系统的不相容输血常引起严重的反应。对于在生育年龄的妇女和需要反复输血的病人,还必须使供血者与受血者的Rh血型相合,以避免受血者在被致敏后产生抗Rh的抗体。
即使在ABO系统血型相同的人之间进行输血,在输血前必须进行交叉配血试验(corss-match test),即不仅把供血者的纸细胞与受血者的血清进行血清配合试验(这称为试验主侧);而且要把受血者的约细胞与供血者的血清作配合试验(这称为试验的次侧)。这样,既可检验血型测定是否有误,又能发现他们的红细胞或血清中,是否还存在一些其它的凝集原或凝集素,足以引起红细胞凝集反应。在进行并叉式配血试验时,应在37℃下进行,以保证可能有的凝集反应得以充分显示。
如果交叉配血试验的两侧都没有凝集反应,即为配血相合,可以进行输血;如果主侧有凝集反应,则为配血不合,不能输血;如果不侧不起凝集反应,而次侧有凝集反应,只能在应急情况下输血,输血时不宜太快太多,并密切观察,如发生输血反应,应立即停止输注。
以往曾经把O型的人称为“万能供血者(universal donor)”,认为他们的血液可以输给其他血液的人。但目前认为这种输血是不足取的,因为,虽然O型的红细胞上没有A和B凝集原,因而不会被受血者的血浆凝集,然而O型人的血浆中的抗A和抗B凝集素能与其它血型受血者的红细胞发生凝集反应。当输入的血量较大时,供血者血浆中的凝集素未被受血者的血浆足够稀释时,受血者的红细胞会被广泛凝集。
总之,输血是一个多环节的过程,每个环节上的失误都可能造成严重事故。因此,大进行输血操作时,必须严格遵守输血原则,密切注意观察;而且只在确实需要时才进行输血。决不可盲目滥用。
参考资料
1.吴祖泽主编,造血干细胞移植基础,第1-4章,北京,人民卫生出版社,1988
2.BrizziMF,Avauzi GC,Pegoraro L.Hematopoitic growth factor receptors.Int J Cell Cloning1991;9:274-300
3.Colman RW,Jack H ed.Hemostasisand Thrombosis 2nd ed,Churchill Livingstone,New York,1987
4.Dainiak N.Surfacemembrane-associated regulation of cell assembly,differentiation and growth.Blood 1991;78: 264-276
5.Dessypris EN,Krantz SB.Erythropoietin:regulation of erythropoiesis and clinical use .Adv Pharmacol1990;21:127-147
6.Harian JM.Leukocyte-endothelialinteractions.Blood 1985;65:513-526
7.Jandl JH.Blood-Testoodk ofHematology.Llttle,Brown Co,Boston,1978
8.Murphy MJ Jr ed.b Blood CellGrowth Factors.Alpha Med Press,Dayton ,1991
9.Pittilo RM,Machin SJed.Platelet-vessel Wall Interactions.Springer-Verlag,London,1988
10.Williams WJ, BeutlerE.Hematology 4thed,McGraw-Hill Publishing Co,New York,1990
第四章 血液循环
心脏和血管组成机体的循环系统,血液在其中按一定方向流动,周而复始,称为血液循环。血液循环的主要功能是完成体内的物质运输,运输代谢原料和代谢产物,使机体新陈代谢能不断进行;体内各内分泌腺分泌的激素,或其它体液因素,通过血液的运输,作用于相应的靶细胞,实现机体的体液调节;机体内环境理化特性相对稳定的维持和血液防卫功能的实现,也都有赖于血液的不断循环流动。
第一节 心脏的泵血功能
心脏是一个由心肌组织构成并具有瓣膜结构的空腔器官,是血液循环的动力装置。生命过程中,心脏不断作收缩和舒张交替的活动,舒张时容纳静脉血返回心脏,收缩时把血液射入动脉,为血液流动提供能量。通过心脏的这种节律性活动以及由此而引起的瓣膜的规律性开启和关闭,推动血液沿单一方向循环流动。心脏的这种活动形式与水泵相似,因此可以把心脏视为实现泵血功能的肌肉器官。
几个世纪以来,生物学家一直认为心脏是一个单纯的循环器官,近年来关于心钠素的研究,认训到心脏除循环功能外,还具有内分泌功能。心钠素是脊椎动物心脏分泌的激素,主要在心房肌细胞内合成,具有利尿、利钠、舒张血管和降血压作用。参与机体水电解质平衡、体液容量和血压的调节(参看本章第四节)。除心钠素外,从哺乳动物的心肌组织中还提取分离出某些生物活性多肽,如抗心律失常肽和内源性洋地黄素等,还发现心肌细胞内有肾素~血管紧张素系统存在。有关心脏内分泌功能的研究进展很快,大大加深丰富了对心脏功能的认识和了解。
心脏活动呈周期性,每个周期中心脏表现出以下三方面活动;①兴奋的产生以及兴奋向整个心脏扩布;②由兴奋触发的心肌收缩和随后的舒张,与瓣膜的启闭相配合,造成心房和心室压力和容积的变化,从而推动血液流动;③伴随瓣膜的启闭,出现心音。心脏泵血作用是由心肌电活动、机械收缩和瓣膜活动三者相联系配合才得以实现。明确每个周期中这三者的变化和相互关系,对于了解心脏如何实现其泵血功能,以及它们将对心脏泵血产生什么影响,都是非常必要的。心脏机械活动的周期称为心动周期;生物是变化周期,称为心肌电周期。
一、心动周期的概念
心脏一次收缩和舒张,构成一个机械活动周期,称为心动周期。心房与心室的心动周期均包括收缩期和舒张期。由于心室在心脏泵血活动中起主要作用。故通常心动周期是指心室的活动周期而言。正常心脏的活动由一连串的的心动周期组合而成,因此,心动周期可以作为分析心脏机械活动的基本单元。
心动周期持续的时间与心跳频率有关。成年人心率平均每分钟75次,每个心动周期持续0.8s。一个心动周期中,两心房首先收缩,持续0.1s,继而心房舒张,持续0.1s,继而心房舒张,持续0.7s。当心房收缩时,心室处于舒张期,心房进入舒张期后不久,心室开始收缩,持续0.3s,随后进入舒张期,占时0.5s。心室舒张的前0.4s期间,心房也处于舒张期,这一时期称为全心舒张期(图4~1)。可见,一次心动周期中,心房和心室各自按一定的时程进行舒张与收缩相交替的活动,而心房和心室两者的活动又依一定的次序先后进行,左右两侧心房或两侧心室的活动则几乎是同步的。另一方面,无论心房或心室,收缩期均短于舒张期。如果心率增快,心动周期持续时间缩短,收缩期和舒张期均相应缩短,但舒张期缩短的比例较大;因此,心率增快时,心肌工作的时间相对延长,休息时间相对缩短,这对心脏的持久活动是不利的。
图4~1 心动周期中心房和心室活动的顺序和时间关系
了解心脏的泵血功能,需要弄清以下三个问题:①血液在心脏内的单方向流动是怎样实现的?②动脉内压力比较高,心脏怎样将血液射入动脉的?③压力很低的静脉血液是怎样返回心脏的?
现以左心室为例,说明心室射血和充盈的过程,以便了解心脏泵血的机制。
(一)左心室的射血和充盈过程
左心室的一个心动周期,包括收缩和舒张两个时期,每个时期又可分为若干时相(图4~2)。通常以心房开始收缩作为描述一个心动周期的起点。
1.心房收缩期心房开始收缩之前,心脏正处于全心舒张期,这时,心房和心室内压力都比较低,接近于大气压,即约oPa(以大气压为零);然而,由于静脉血不断流入心房,心房压相对高于心室压,房室瓣处于开启状态,心房腔与心室腔相通,血液由心房顺房~室压力梯度进入心室,使心室充盈。而此时,心室内压远比主动脉压(约80mmHg即10.6kPa)为低,故半月瓣是关闭着的,心室腔与动脉腔不相连通。
心房开始收缩,心房容积缩小,内压升高,心房内血液被挤入已经充盈了血液但仍然处于舒张期状态的心室,使心室的血液充盈量进一步增加。心房收缩持续约0.1s,随后进入舒张期。
2.心室收缩期包括等容收缩相以及快速和减慢射血相。
(1)等容收缩相:心房进入舒张期后不久,心室开始收缩,心室内压力开始升高;当超过房内压时,心室内血液?
■[此处缺少一些内容]■
左右时间内,心室肌仍在作强烈收缩,由心室射入主动脉的血液量很大(约占总射血量的2/3左右),流速也很快,此时,心室容积明显缩小,室内压继续上升达峰值,这段时期称快速射血相(0.10s);由于大量血液进入主动脉,主动脉压相应增高.随后,由于心室内血液减少以及心室肌收缩强度减弱,心室容积的缩小也相应变得缓慢,射血速度逐渐减弱,这段时期称为减慢射血相(0.15s),晕一时期内,心室内压和主动脉压都相应由峰值逐步下降。
早期的实验表明,整个射血相内,心室压始终高于主动脉压,这种心室~动脉压力梯度是血液由心室进入动脉的推动力;然而,近代应用精确的压力测量方法观察到,在快速射血的中期或稍后,心室内压已经低于主动脉压(图4~2),不过此时,心室内血液因为受以心室肌收缩的作用而具有较高的动能,依其惯性作用可以逆着压力梯度继续射入主动脉。
3.心室舒张期包括等容舒张相和心室充盈相,后者又再细分为快速充盈、减慢充盈和心房收缩充盈三个时相。
(1)等容舒张相:心室肌开始舒张后,室内压下降,主动脉内血液向心室方向返流,推动半朋瓣关闭;这时室内压仍明显高于心房压,房室瓣仍然处于关闭状态,心室又成为封闭腔。此时,心室肌舒张,室心压极快的速度大幅度下降,但容积并不改变,从半月瓣关闭直到室内压下降到低于心房压,房室瓣开启时为止,称为等容舒张相,持续约0.06~0.08s。
(2)心室充盈相:当室内压下降到低于心房压时,血液顺着房~室压力梯度由心房向心室方向流动,冲开房室瓣并快速进入心室,心室容积增大,称快速充盈相,占时0.11s左右;其间进入心室的血液约为总充盈量的2/3。随后,血液以较慢的速度继续流入心室,心室容积进一步增大,称减慢充盈相(0.22s)。此后,进入下一个心动周期,心房开始收缩并向心室射血,心室充盈又快速增加。亦有人将这一时期称为心室的主动快速充盈相(占时0.1s)。
从以上对心室充盈和射血过程的描述中,不难理解左心室泵血的机制。室壁心收缩和舒张,是造成室内压力变化,从而导致心房和心室之间以及心室和主动脉之间产生压力梯度的根本原因;而压力梯度是推动血液在相应腔室内之间流动的主要动力,血液的单方向流动则是在瓣膜活动的配合下实现的。还应注意瓣膜的作用对于室内压力的变化起着重要作用,没有瓣膜的配合,等容收缩相和等容舒张相的室内压大幅度升降,是不能完满实现的。
(二)心动周期中心房压力的变化
每一心动周期中,左心房压力曲线依次出现三个小的正向波:a波、c波和v波,以及两个下降波:x降波和y降波。首先,疏收缩,房内压升高,形成a波,随后心房舒张,压力又回降。以后心室开始收缩,室内压升高,室内血液推顶并关闭了房室瓣,使瓣膜叶片向心房腔一侧凸出,造成房内压轻度上升,形成c波。随着心室射血时体积的缩小,心底部向下移动,房室瓣从而也被向下牵,以致心房的容积趋于扩大,房内压下降,形成x降波。以后,静脉血不断流入心房,而房室瓣尚关闭着,血液不能入心室,心房内血液量不断增加,房内压缓慢而持续升高,直到心室等容舒张相结束,心房血得以进入心室为止,由此形成的上升波称v波。随后房室瓣开放,血液由心房迅速进入心室,房内压下降,形成y降波。
一个心动周期中,心房压力波动的幅度较小。成年人于安静卧位,左房压变化幅度为0.3~1.6kPa(2~12mmHg);右房压为0~0.7kPa(0~5mmHg)。
(三)心房和心室在心脏泵血活动中的作用
心室~动脉压力梯度是引起半月瓣开放、推动血液由心室开始射入动脉的直接动力,这种压力梯度是由心室的强烈收缩千百万室内压由原来近于心房压水平升高到超过动脉压而形成的。同样,房~室压力梯度是由血液由心房流入心室的动力,但它的形成主要并不是来自心房收缩,而是依靠心室的舒张;即在心室等容舒张相,室内压大幅度下降,由开始时近于动脉压一直下降到低于心房压,房室瓣开放,血液由心房迅速进入心室。整个心室舒张充盈期内,房~室压力梯度始终存在;然而,这一时期的前一段时间内(即充盈期的前4/5时间内),心房也处于舒张状态,这时心房只 不过是静脉血液返回心室的一条通道,只有后1/5期间心房才收缩。由此可以看出,心房收缩对于心室充盈不起主要作用。故当民生心房纤维性颤动时,虽然心房已不能正常收缩,心室充盈量因此有所减少,但一般不致于严重影响心室的充盈和射血功能;如果发生心室纤维性颤动,心脏泵血活动立即停止,后果十分严重。
虽然心室的充盈绝大部分是在快速充公盈期内完成的,但心房收缩时又挤出部分血液以增加心室充盈(约占总充盈量的30%左右),使心室舒张末期容积和压力都有一定程度增加,这对于心室射血功能是有利的。另一方面,如果心房收缩缺失,将会导致房内压增加,不利于静脉血液回流,从而间接影响心室射血。可以认为,心房收缩起着初级泵的作用,对于心脏射血和血液的回流都是不利的。房泵作用的缺失,对静息状态下心脏泵血功能影响不大;但机体在运动和应急状态下,就可能出现心输出量不足等泵功能的严重损害。
三、心脏泵功能的评定
心脏泵功能是正常或是不正常,是增强或减弱,这是医疗实践以及实验研究工作中经常遇到的问题。因此,用什么样的方法和指标来测量和评定心脏功能,在理论和实践上都是十分重要的。
(一)心脏的输出量
心脏在循环系统中所起的主要作用就是泵出血液以适应机体新陈代谢的需要,不言而喻,心脏输出的血液量是衡量心脏功能的基本指标。
1.每分输出量和每搏输出量一次心跳一侧心室射出的血液量,称每捕输出量,简称搏出量。每分钟射出的血液量,称每分输出量,简称心输出量,等于心率与搏出量的乘积。左右两心室的输出量基本相等。
心输出量与机体新陈代谢水平相适应,可因性别、年龄及其它生理情况而不同。如健康成年男性静息状态下,心纺平均每分钟75次,搏出量约为70ml(60~80ml),心输出量为5L/min(4.5~6.0L/min)。女性比同体重男性的心输出量约低10%,青年时期心输出量高于老年时期。心输出量在剧烈运动时可高达25~35L/min,麻醉情况下则可降低到2.5L/min。
2.心指数心输出量是以个体为单位计算的。身体矮小的人和高大的人,新陈代谢总量不不相等,因此,用输出量的绝对值作为指标进行不同个体之间心功能的比较,是不全面的。群体调查资料表明,人体静息时的心输出时,也和基础代谢率一样,并不与体重成正比,而是与体表面积成正比的。以单位体表面积(m2)计算的心输出量,称为心指数;中等身体的成年人体表面积约为1.6~1.7m2,安静和空腹情况下心输出量约5~6L/min,故心指数约为3.0~3.5L/min·m2。安静和空腹情况下的心指数,称之为静息心指数,是分析比较不同个体心功能时常用的评定指标。
心指数随不同重量条件而不同。年龄在10岁左右时,静息心指数最大,可达4L/min·m2以上,以后随年龄增长而逐渐下降,到80岁时,静息心指数接近于2L/min·m2.肌肉运动时,心指数随运动强度的增加大致成比例地增高。妊娠、情绪激动和进食时,心指数均增高。
(二)射血分数
心室舒张末期充盈量最大,此时心室的容积称为舒张末期容积。心室射血期末,容积最小,这时的心室容积称为收缩末期容积。舒张末期容积与收缩末期容积之差,即为搏出量。正常成年人,左心室舒张末期容积估计约为145ml,收缩末期容积约75ml,搏出量为70ml。可见,每一次心跳,心室内血液并没有全部射出。搏出量占心室舒张末期容积的百分比,称为射血分数。健康成年人搏出量较大时,射血分数为55%~65%。
在评定心泵血功能时,单纯用搏出量作指标,不考虑心室舒张末期容积,是不全面的。正常情况下,搏出量始终与心室舒张末期容积相适应,即当心室舒张末期容积增大时,搏出量也相应增加,射血分数基本不变。但是,在心室异常扩大、心室功能减退的情况下,搏出量可能与正常人没有明显判别,但它并不与已经增大的舒张末期容积相适应,射血分数明显下降。若单纯依据搏出量来评定心泵血功能,则可能作出错误判断。
(三)心脏作功量
血液在心血管内流动过程中所消耗的能量,是由心脏作功所供给的;换句话说,心脏作功所释放的能量转化为压强能和血流的动能,血液才能循环流动。
心室一次收缩所作的功,称为每搏功,可以用搏出的血液所增加的动能和压强能来表示。心脏射出的血液所具有的动能在整个搏功中所占比例很小,可以略而不计。搏出血液的压强能可用平均动脉压表示,约相当于;舒张压+(收缩压~舒张压)×1/3。由于心室充盈是由静脉和心房输送回心的血液充盈心室造成的,计算心室收缩释放的能量时不应将充盈压(可用左室舒张末期压或平均左房压表示,约为0.8kPa[6mmHg])计算在内.搏功单位为g~m。搏功乘以心率即为每分功,单位为kg~m/min。计算左室搏功和每分功的简式如下:
搏功(g~m)=搏出量(cm3)×(1/1000)×(平均动脉压~平均左房压mmHg)×(13.6g/cm3)
每分功(kg~m/min)=搏功(g~m)×心率×(1/1000)
设搏出量为70ml,收缩压120mmHg,舒张压80mmHg,平均左房压6mmHg,心纺75次,代入上式,求得左心室搏功为83.1g~m;每分功为6.23kg~m/min。
右心室搏出量与左心室相等,但肺动脉平均压仅为主动脉平均压的1/6左右,故右心室作功量也只有左心室的1/6。
用作功量来评定心泵血功能,其意义是显而易见的,因为心脏收缩不仅仅是排出一定量的血液,而且这部分血液具有适用高的压强能(以及很快的流速)。在动脉压增高的情况下,心脏要射出与原先同等量的血液就必须加强收缩;如果此时心肌收缩的强度不变,那么,搏出量将会减少。实验资料表明,心肌的耗氧量与心肌的作轼量是相平行的,其中,心输出量的变动不如心室射血期压力和动脉压的变动对心肌耗氧量的影响大。这就是说,心肌收缩释放的能量主要用于维持血压。由此可见,作为评定心泵血功能的指标,心脏作功量要比单纯的心输出量,更为全面。在需要对动脉压不相等的各个人,以及同一个人动脉压发生变动前后的心脏泵血功能进行分析比较时,情况更是如此。
四、心脏泵功能的调节
机体在长期进化的过程中, 发生和发展了一套逐步完善的循环调节机构,使循环功能适应于不同生理情况下新陈代谢的需要。这种调节是在复杂的神经和体液机制参与下,通过对心脏和血管活动的综合调节而实现的(整体调节机制将后述)。本节主要从心脏本身来阐述控制心输出量的因素的作用机制。
心输出量取决于心率和搏出量,机体通过对心率和搏出量两方面的调节来调节心输出量。
(一)搏出量的调节
心室肌的收缩是面临着动脉压(后负荷)的阻挡进行的。等容收缩相内,心室肌收缩首先引起室内压升高,只有当室内压升高到超过动脉压时,心肌纤维才能缩短,心室容积才能缩小,血液才能射出。由此可见,在心率恒定情况下,心室每次收缩的射血量取决于心肌纤维缩短的程度和速度,这决定了心肌收缩产生张力(表现为心室内血液的压力)的程度和速度。凡是能影响心肌收缩强度和速度的因素都能影响搏出量,而搏出量的调节正是通过改变心肌收缩的强度和速度来实现的。
1.心肌收缩的“全或无”现象心肌细胞和骨骼肌细胞一样,收缩反应是由肌膜的兴奋通过兴奋~收缩耦联触发引起的;但是,从整块肌肉来看,在骨骼肌,一个细胞产生的兴奋不能扩布到另一个细胞,只有在支配一个肌细胞有神经纤维发放神经波动时,才会出现收缩效应;若发放冲动的神经纤维数目增加,可以引起更多的肌细胞兴奋和收缩,整块肌肉的收缩增强。因此,如果骨骼肌作单收缩,其收缩强度的改变可以来自两个方面,一是由于单个肌细胞收缩强度的改变,另一原因是参与收缩活动的肌细胞数目发生变化。心肌则不同,相邻的心肌细胞是由闰盘的特殊结构和特性,兴奋可以通过它由一个心肌细胞传播到另一个心肌细胞。因此,整个心室(或整个心房)可以看成是一个功能上互相联系的合胞体,产生于心室某一处的兴奋可以在心肌细胞之间迅速传递,引起组成心室的所有心肌细胞几乎同步收缩。从参与活动的肌细胞数目上看,心肌的收缩是“全或无”的。这就是说,心肌据要么不产生,一旦产生则全部心肌细胞都参与收缩。由此可见,心肌收缩的强度,不象骨骼肌那样可以因参加活动的细胞数目的不同而改变;各个心肌细胞收缩强度的变化是整块心肌收缩强度发生的唯一原因。所以搏出量的调节可以从单个心肌细胞收缩功能调控的角度来探讨。
2.搏出量的调节搏出量和搏功的调节,是心脏生理中最基本最重要地课题之一,从世纪末、本世纪初以来,众多学者对此进行了大量实验研究。50年代中期,Sarnoff对这些研究资料进行详细分析之后,将搏出量和搏功的调节归纳为:①由初长度改变引起的异长调节,以及②由心肌收缩能力改变引起的等长调节等方式。
(1)异长调节:前已述,控制骨骼股收缩强度和速度的一个重要因素就是肌肉本身的初长度,而初长度是被动地由该肌肉收缩前所随的负荷(前负荷)所决定的。前负荷和初长度对肌肉收缩强度和作功能力的影响是双相的,构成的长度~张力曲线类似抛物线,在前负荷和初长度达最适水平之前,肌肉收缩强度和作功能力随前负荷~初长度的增加而增加;超过最适水平,收缩效果将随前负荷~初长度的继续增加而降低(图4~3)。
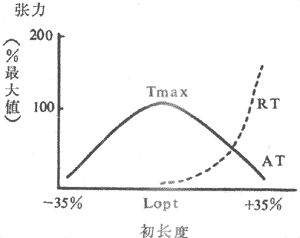
图4~3 骨骼肌的长度~张力曲线
AT:主动张力 RT:静息张力(被动张力)
Tmax:最大张力 Lopt:最适初长
在心肌,初长度同样是控制收缩功能最重要的因素,不过,初长度和收缩功能的关系具有某些特殊性。为了分析前负荷和初长度对心脏泵血功能的影响,可以在实验中逐步改变心室舒张末期压力(亦称充盈压)和容积(相当于前负荷或初长),并测量射血心室的搏功或等容心室的室内峰压,将一系列搏功或室内峰压数据对应心室舒张末期压力和容积,绘制成坐标图,即为心室功能曲线(图4~4)。心室功能曲线大致可分为三段:①充盈压1.6~2kPa(12~15mmHg)是人体心室最适前负荷,位于其左侧的一段为功能曲线升支,它与骨骼肌长度~长力曲张升支段相似;均表明当前负荷~初长度未达最适水平之前,搏功或等容峰压随初长度的增加而增加,通常情况下,左室充压约0.7~0.8kPa(5~6mmHg),可见正常心室是在功能曲线的升支段工作,前负荷~初长度尚远离其适水平.这一特征表明心室具有较大程度的初长度贮备,心室通过前负荷~初长度的增加即异长调节机制使泵血功能增强的容许范围是很宽的.而体内骨骼肌的自然长度已经接近最适长度,前负荷~初长度贮备很小,通过初长度调节其收缩功能的范围也很小;②充盈压2~2.7kPa(15~20mmHg)范围内,曲线逐渐平坦,说明前负荷在上限范围内变动时对泵血功能的影响不大;③随后的曲线呈平坦状,或轻度下倾,并不出现明显的降支,这一点明显不同于骨骼,说明正常心室充压即使超过2.7kPa(20mmHg),搏功不变或仅轻度减少,只有在民生严重病理变化的心室,功能曲线才出现降支。
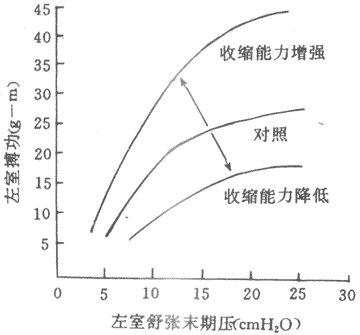
图4~4 心室功能曲线(lcmH2O=0.098kPa)
在一组心肌肌小节标本上的研究表明,在心室最适前负荷和最适初长度时,肌小节初长度为2.0~2.2μm,这正是肌小节的最适初长度,粗细肌丝处于最佳重叠状态。这种情况下,肌小节等长收缩产生的张力最大。达最适水平之前,随着前负荷和肌小的初长度的增加,粗细肌丝有效地重叠的程度增加,激活时可能形成的横桥联接的数目相应增加,肌小节以至整个心室的收缩强度增加,搏出量和搏功增加。由此可见,心室功能曲线的升支正是肌小节初长度~主动张力关系的升支在整个心室功能上的一种反映。在搏出量的这种调节机制中,引起调节的因素是心肌细胞本身初长度的改变,其效应是心肌细胞收缩强度的改变,因些将这种形式的调节称为异长调节。
为什么骨骼肌的长度~张力曲线出现降支,而正常心室的功能曲缍不再现降支?已知,一个具有弹性的物质受到外力作用时,一方面可被拉长,随之产生弹性张力,与此同时,这一个弹性张力又具有阻止它本身被进一步拉长的作用。当骨骼肌处于最适长度即肌小节初长度为2.0~2.2μm的状态下,所产生的静息张力较小,继续增加前负荷,肌细胞可被进一步延伸,肌小节初长度可大大超过2.2μm,肌丝重叠程度明显减小,收缩能力因此下降。但是,由于心肌细胞外间内含有大量劲度较大的胶原纤维,因而心肌的伸展性较小,处于最适初长度时,产生的静息张力已经很大,从而阻止心肌细胞继续被拉长。实验证明,即使在前负荷很大的情况下,心肌肌小节初长度一般也不会超过2.25~2.30μm。心肌细胞这种抵抗过度延伸的特性,对心脏泵血功能具有重要生理意义,它使心脏不致于被在前负荷明显增加时,引起搏出师长作功能力的下降。心室功能曲线不再现降支,并不是心肌初长度超过最适水平之后心肌的收缩功能依然不受影响,而只是在这种情况下初长度不再与前负荷呈平行关系。而在慢性过度扩张的病理心脏上观察到的功能曲线降支,是由于室壁心肌发生了组织学改变,心室容积扩大,心肌细胞收缩功能又严重损伤的缘故。
既然前负荷~初长度是调节搏出量的主要因素,那么在体情况下有哪些因素可以影响心室的前负荷和初长度?在心室其它条件(主要是心室肌的顺应性)不变的情况下,心室前负荷~舒张末期压是由心室舒张末期充盈的血液量来决定的,充盈量大,舒张末期容积也较大。可以设想,凡是影响心室充盈量的因素,都能够通过异长调节机制来使搏出量发生改变。
心室充盈的血量,是静脉回心血量和心室射血剩余血量两者的总和。静脉回心血量又受下述因素的影响:①心室舒张充盈期持续时间。例如,心率增快时,充盈期缩短,心室充盈不完全,充盈压降低,搏出量减少;②静脉回流速度。在充盈期持续时间不变的情况下,静脉内血液通过惊讶进入心室的速度愈快,充盈量愈大,搏出量相应增加。静脉加流速度取决于外周静脉压与心房压和心室压之差。外周静脉增高(如循环血量增加、外周静脉管壁张力增高等情况下)和(或)心房心室压力降低时,可促进静脉回流。心脏每次射血之后的剩余血液量,也影响心室的充盈量,但是,这种影响是多方面的;如果静脉回心血量不变,心室剩余血量的增加将导致部充盈量增加,充盈压增高,搏出量随之增加;但另一方面,当心室剩余血量增加时,心室舒张期内室压增高,静脉回心血量将因此减少,总充盈量不一定增加。总之,在心室射血功能不为的情况下,心室剩余血量的增减对搏出量是否有影响以及发生何种影响,取决于心室总充盈量是否改变以及发生何种改变。
早在1914~1918年,生理学家Starling在哺乳动物身上就观察到肌纤维初长度对心脏的功能的影响,因此异长调节也称为Starling机制,心室功能曲线也可称为Starling曲线。通常,心室射血量与静脉回心血量相平衡,从而维持心室舒张末期压力和容积于正常范围,如果因某种原因造成静脉回心血量超过射血量,则充盈压将增高,通过Starling机制增加搏出量使之与回流量重新达到平衡,否则舒张末期压力和容积将持续增高而得不到纠正。但Starling机制的主要作用是对搏出量进行精细的调节。例如,当体位改变以及当左右心室搏出量不平衡等情况下所出现的充盈量的微小变化,是通过异长调节机制来改变搏出量使之与充盈量达到平衡的;而对于持续的、剧烈的循环功能变化,例如体力劳动时搏出量持久且大幅度的增高,主要靠心肌收缩能力的变化来调节,这时异长调节机制的作用不大。
(2)等长调节:人们进行强体力劳动时,搏出量和搏功可成倍增加,而此时心脏舒张末期容积不一定增大,甚至有所减小;相反,心力衰竭病人,心脏容积扩大而其作功能力反而降低。由此推测,对于心脏的泵血功能,除异长调节处,还有另一种与心肌初长度无关的调节机制存在。精确的实验研究表明,将离体心有保持在最适长度情况下,支甲肾上腺素促使心肌等长收缩产生的最大张力和张力上升速率都增加,等张收缩的幅度增大,缩短速度加快,将一系列等长收缩或等张收缩的实验结果绘制成长度~张力曲线和张力~速度曲张,可以看到,在去甲肾上腺素作用下,长度~张力曲线升支向左上方移位,张力~速度曲张向右上方移位。这些结果表明,处于任何一种初长度下(不只是最适初长度)的心肌,在有去甲肾上腺素的情况下,等长收缩产生的张力都比没有去甲肾上腺素时为高;后一结果表明,任何一种后负荷情况下,受去甲肾腺素作用的心肌等张收缩的缩短速度增快。完整心室的结果类似:给予去甲肾上腺素后,心肺功能曲线左上移位,表明有同一前负荷~舒末容积条件下,等容心室的峰压增高,射血心室容积缩小程度增加;室内压上升速率和射血期容积缩小的速度也能增加。其结果,搏出量和搏功增加,心脏泵血功能明显加强。另一些因素,如乙酰胆碱,则引起相反效应。很明显,上述干预因素引起心肌收缩功能和心脏泵血功能改变的原因并不是初长度的变化,而是通过心肌另一种功能变数起作用的。这种功能变数称之为心肌收缩能力(myocardial contractility)。心肌收缩能力一般地定义为:心肌不依赖于负荷而改变其力学活动(包括收缩活动的强度和速度)的一种内在特性,因此,心肌收缩能力又称为心肌变力状态(inotropic state)。在某些因素作用下,心肌收缩能力发生改变,从而影响心肌细胞学活动的强度和速度,使心脏搏出量和搏功相应发生改变(图4~4)。心脏泵血功能的这种调节是通过收缩能力这个与初长度无关的、心肌内在功能变数的改变而实现的,故称等长调节。
心肌收缩能力受多种因素的影响,兴奋~收缩耦联过程中各个环节(参看第二章)都能影响收缩能力,其中活化横桥数和肌凝蛋白的ATP酶活性是控制收缩能力的主要因素,已知,粗肌丝上的横桥,只有与细肌丝的肌纤蛋白结合形成横桥联接并活化,才能导致肌丝滑行并产生力。在一定初长度的条件下,粗细肌丝的重叠区提供了可以形成横桥联接的最大横桥数,然而不是所有横桥都会形成活化横桥。海参化横桥数与最大横桥数的比例,取决于兴奋后胞浆Ca2+浓度的升高程度和肌钙蛋白对Ca2+的亲和力;凡能增加兴奋后胞浆Ca2+浓度(或)肌钙蛋白Ca2+亲和力的因素,均可增加活化横桥的比例,导致收缩能力的增强。例如儿茶酚胺增加收缩能力的原因之一,就是它通过激活β~肾上腺素能受体,增加胞浆cANP浓度,使肌膜Ca2+通道和肌浆网Ca2+的通道的开放程度增加,导致心肌兴奋后胞浆Ca2+浓度升高程度增加。一些钙增敏剂,如茶碱,可以增加肌钙蛋白对Ca2+的亲和力使肌钙蛋白结合形成活化横桥,肌凝蛋白的ATP酶就被激活,它分解ATP以提供肌丝滑行的能量。甲状腺激素和体育锻炼能提高肌凝蛋白ATP酶活性,促进心肌收缩能力增强;相反,老年人的心脏和甲状腺功能减退患者的心脏,心肌肌凝蛋白分子结构发生改变,其ATP酶的活性较低,收缩能力减弱。
心肌收缩能力的评定:
泵血功能反映的是心室前负荷的后负荷(见后文)、心肌收缩能力及及心率等变数的综合效果,而心肌收缩能力直接反映心肌本身的功能状态,收缩能力的改变具有极其重要的生理和病理意义。因此,理论研究和临床实践中都需要对收缩能力进行定量测量和评定,以了解收缩能力的水平和变化。由于收缩能力并不是某种可测量的单一变数,因此,对收缩能力的具体度量是比较困难的。衡量泵血功能的指标(如搏出量、搏功等)受前、后负荷影响,并不能直接反映收缩能力的水平,只有根据心肌长度~张力曲线或心室功能曲线的移位来判断收缩能力的变化(左上移位提示收缩能力增强,右下移位表明收缩能力低下,图4~4)。但这种评定方法具有操作繁杂、敏感性较低的缺点。目前,常用的方法是采用一系列速度指标来定量评定收缩能力。在离体心肌,最常采用的是张力变化速率(Dt/dt)和长度变化速率(Dl/dt);对完整心室,常采用的指标有:等容相室内压变化速率(Dp/dt)、射血相心室容积变化速率(dV/dt)或心室直径变化速率(dD/dt)等,以及心肌纤维收缩成分的缩短程度(VCE)等。这些速度指标受负荷改变的影响较小,对收缩能力的变化比较敏感,为国内外心脏学研究工作者广泛采用。
(3)后负荷对搏出量的影响:已知,离体肌肉的后负荷对收缩过程产生一定的影响,对心室而言,动脉压起着后负荷的作用,因此,动脉压的变化将影响心室肌的收缩过程,影响搏出量。在心率、心肌初长度和收缩能力不变的情况下,如果动脉压增高,等容收缩相室内压峰值必然也增高,从而使等容收缩相延长而射血相缩短,同时,射血相心室肌纤维缩短的程度和速度均减小,射血速度减慢,搏出量因此减少。应该看到,后负荷对心肌收缩活动的上述影响,是一种单纯机械效应,并不是某种功能调节机制进行调节的结果。从心肌收缩释放的能量的分配比例上,可以很容易理解这个问题。当心肌收缩能力不变时,收缩释放的能量是恒定的。如果室内压力升高,表示心肌产生张力消耗的能量相对增加,用于心肌纤维缩短将相对减少;如果肌肉作等长收缩,释放的能量几乎全部用于产生张力。另一方面,也应该看到,由于动脉压影响了搏出量,又常常继发地引起一些调节机制的活动。如上述,动脉压的突然升高首先导致搏出量减少,结果造成心室内剩余血量增加,充盈量增加,后者又再引起异长调节机制的调节,搏出量可以恢复正常水平,即通过异长调节可以使动脉压增高所招致的搏出量减少的现象得到纠正。但是,既然通过异长调节,搏出量已经恢复原有水平,那么,心室的充盈一不是将同时恢复原水平,异长调节的作用被取消,搏出量不是又因高动脉压状态的存在而减少了吗?这就是说,搏出量是否会出现时而降低、时而恢复的现象?许多实验都证明,随着搏出题的恢复,心室舒张末期容积确实也恢复到原水平,但此时,搏出量仍然维持高水平,并不出现再次减少的现象。进一步分析发现,后一情况的出现乃是心肌收缩能力增强所致。尽管此时产生的张力随负荷的增加而有所增加,但缩短的程度不但可以不降低,反而可能增加。至于这种情况下心肌收缩能力为什么增强,原因尚不十分清楚,可能是神经机液机制调节的结果。
综上所述,心室后负荷的本身直接着搏出量,随后通过异长和等长调节机制,使前负荷和心肌收缩能力与后负荷相匹配,从而使机体得在在动脉压增高的情况下,能够维持适当的心输出量。这种情况对于机体是有重要生理意义的。但也应看到,此时搏出量的维持,是心肌加强收缩的结果,如果动脉压持续增高,心室肌将因处于收缩强状太而逐渐肥厚,即发生了病理性改变,随后将导致泵血功能减退。
(二)心率及其对心输出量的影响
健康成年人的安静状态下,心率平均为每分钟75次(正常范围为每分钟60~100次)。不同生理条件下,心率有很大变动,可低到每分钟40~50次,高达每分钟200次。
心输出量是搏出量与心率的乘积,心率增快,心输出量增加;但这有一定的限度,如果心率增加过快,超过每分钟170~180次,心室充盆时间明显缩短,率盈量减少,搏出题可减少到仅有正常时的一半左右,心输出量亦开始下降。当心率增快但尚未超过此限度时,尽管此时心室充盈时间有所缩短,但由于回心血量中的绝大部分是在快速充盈期内进入心室的,因此,心室充盈量以及搏出量不致于减少或过分减少,而由于心庇护增加,每分钟的输出量增加。反之,如心率太慢,低于每分钟40次,心输出量亦减少。这是因为心室舒张期过长,心室充盈早已接近限度,再延长心舒时间也不能相应增加充盈量和搏出量。可见,心跳频率最适宜时,心输出量最大,心率过快或过慢,心输出量都会减少。
心率受自主神经的控制,交感神经活动增强时,心率增快;迷走神经活动增强时,心率减慢。影响心率的体液因素主要有循环血液中的肾上腺和去甲肾上腺素,以及甲状腺素。此外,心率受体温的影响,体温升高1℃,心率将增加12~18次。
(三)心脏泵功能的贮备
心脏的泵血功能能够广泛适应机体不同生理条件下的代谢需要,表现为心输出量可随机体代谢增长而增加。健康成年人静息状态下心率每分钟75次,搏出量约70ml,心输出量为5L左右。强体力劳动时,心率可达每分钟180~200次,搏出量可增加到150ml左右,心输出量可达25~30L,为静息时的5~6倍。心脏每分钟能射出的最大血量,称最大输出量。它反映心脏的健康程度。由上可以看出,在平时,心输出量产是最大的,但能够在需要时成倍地增长,表明健康人心脏泵血功能有一定的贮备力量。心输出量随机体代谢需要而增加的能力,称为泵功能贮备,或心力贮备。健康人有相当大的心力贮备,而某些心脏疾患的病人,静息时心输出量与健康人没明显差别,尚能够满足静息状态下代谢的需要,但在代谢活动增强时,输出量却不以相应增加,最大输出量较正常人为低;而训练有素的运动员,心脏的最大输出量远比一般人为主,可达35L以上,为静息时的8倍左右。
五、心音心音图
心动周期中,心肌收缩、瓣膜启闭、血液加速度和减速度对心血管壁的加压和减压作用以及形成的涡流等因素引起的机械振动,可通过周围组织传递到胸壁;如将听诊器放在胸壁某些部位,就可以听到声音,称为心音。若用换能器将这些机械振动转换成电信号记录下来,便得到了心音图。
心音发生在心动周期的某些特定时期,其音调和持续时间也有一定的规律;正常心脏可听到4个心音:即第一、第二、第三和第四心音。多数情况下只能听到第一和第二心音,在某些健康儿童和青年人也可听到第三心音,40岁以上的健康人也有可能出现第四心音。心脏某些异常活动可以产生杂音或其它异常心音。因些,听取心音或记录心音图对于心脏疾病的诊断有一定的意义。
第一心音发生在心缩期,音调低,持续时间相对较长,在心尖搏动处(左第五肋间隙锁骨中线)听得最清楚。在心缩期心室射血引起大血管扩张及产生的涡流发出的低频振动,有及由于房室瓣突然关闭所引起的振动,是听诊的第一心音的主要组成成分,因经,通常可用第一心音人选为心室收缩期开始的标志。第二心音发生在心脏舒张期,频率较主,持续时间较短。听诊的第二心音主要与主动脉瓣的关闭有关,故可用用标志心室舒张期开始。第三心音发生在快速充盈期末,是一种低频、低振幅的心音。它可能是由于心室快速充盈期末,血流充盈减,流速突然改变,形成一种力使心室壁和瓣膜发生振动而产生的。第四心音是与心房收缩有关的一组心室收缩期前的振动,故也称心房音。正常心房收缩,听不到声音,但在异常有力的心房收缩和左室壁变硬的情况下,心房收缩使心室充盈的血量增加,心室进一步扩张,引起左室肌及二尖瓣和血液的振动,则可产生第四心音。
第二节 心肌的生物电现象和生理特征
心房和心室不停歇地进行有顺序的、协调的收缩和舒张交替的活动,是心脏实现泵血功能、推动血液循环的必要条件,而细胞膜的兴奋过程则是触发收缩反应的始动因素。本节需要阐述的问题是:引起心脏收缩活动的兴奋来自何处?为什么心脏四个腔室能够作协调的收缩活动?为什么心脏的收缩活动始终是收缩和舒张交替而不出现强直收缩?要回答这些问题,必须了解心肌的生理特性,主要是心肌兴奋和兴奋传导的特征。兴奋和传导是以细胞膜的生物电活动为基础的。因此,首先叙述心肌细胞的生物电现象,然后,根据生物电现象分析叙述心肌兴奋和兴奋传播的规律和生理意义。
心肌细胞的类型组成心脏的心肌细胞并不是同一类型的,根据它们的组织学特点、电生理特性以及功能上的区别,粗略地分为两大类型:两类心肌细胞分别实现一定的职能,互相配合,完成心脏的整体活动。一类是普通的心肌细胞,包括心房肌和心室肌,含有丰富的肌原纤维,执行收缩功能,故又称为工作细胞。工作细胞不能自动地产生节律性兴奋,即不具有自动节律性;但它具有兴奋性,可以在外来刺激作用下产生兴奋;也具有传导兴奋的能力,但是,与相应的特殊传导组织作比较,传导性较低。另一类是一些特殊分化了的心肌细胞,组成心脏的特殊传导系统;其中主要包括P细胞和哺肯野细胞,它们除了具有兴奋性和传导性之外,还具有自动产生节律性兴奋的能力,故称为自律细胞,它们含肌原纤维甚小或完全缺乏,故收缩功能已基本丧失。还有一种细胞位于特殊传导系统的结区,既不具有收缩功能,也没有自律性,只保留了很低的传导性,是传导系统中的非自律细胞,特殊传导系统是心脏内发生兴奋和传播兴奋的组织,起着控制心脏节律性活动的作用。
心脏特殊传导系统的组成和分布心脏的特殊传导系统由不同类型的特殊分化的心肌细胞所组成。包括窦房结、房室交界、房室束和末梢浦肯野纤维网(图4-5)。
窦房结:位于右心房和上腔静脉连接处,主要含有P细胞和过渡细胞。P细胞是自律细胞,位于窦房结中心部分;过渡细胞位于周边部分,不具有自律性,其作用是将P细胞自动产生的兴奋向外传播到心房肌。
房室交界:又称为房室结区,是心房与心室之间的特殊传导组织,是心房兴奋传入心室的通道。房室交界主要包括以下三个功能区域:
房结区:位于心房和结区之间,具有传导性和自律性。
结区:相当于光学显微镜所见的房室结,具有传导性,无自律性。
结希区:位于结区和希氏束之间,具有传导性和自律性。
房室束(又称希氏束)及其分支:房室束走行于室间隔内,在室间隔膜部开始分为左右两支,右束支较细,沿途分支少,分布于右心室,左束支呈带状,分支多,分布于左心室,房室束主要含浦肯野细胞。
图4-5 心脏各部分心肌细胞的跨膜电位和兴奋传导速度
SAM:窦房结 AM:心房肌 AVN;结区 BH:希氏束 PE;哺肯野纤维
TPF:末梢浦肯野纤维 VM:心室肌传导速度单位m/s
浦肯野纤维网:是左右束支的最后分支,由于分支很多,形成网状,密布于左右心室的心内膜下,并垂直向心外膜侧伸延,再与普通心室肌细胞相连接。房室束及末梢浦肯野纤维网的作用,是将心房传来的兴奋迅速传播到整个心室。
关于是否存在心房传导束的问题,争论很多。60年代,Janes提出在窦房结和房室结区之间有三条由浦肯野细胞构成的心房传导束,分别称前、中、后结间束,其兴奋传导速度比一般心房肌为快。但是,近20年来的研究未能证实心房内有形态结构上不同于心房(工作)细胞的特殊传导组织组成的心房传导束存在;另一方面,研究结果表明,在右心房的某些部位(如卵圆窝前方和界嵴处)心房肌纤维排列方向一致,结构整齐,因此其传导速度较其它部位心房肌(这些心房肌被右心房壁上腔静脉开口卵圆窝所形成的孔穴所分割,形成断续状)为快,从而在功能上构成了将窦房结兴奋快速传播到房室交界处的所谓优势传导通路(preferential pathway) 。
一、心肌细胞的生物电现象
与骨骼肌相比,心肌细胞的跨膜电位在波形上和形成机制上要复杂得多;不但如此,上述不同类型的心肌细胞的跨膜电位(图4-5),不仅幅度和持续时间各不相同,而且波形和形成的离子基础也有一定的差别;各类心肌细胞电活动的不一致性,是心脏兴奋的产生以及兴奋向整个心脏传播过程中表现出特殊规律的原因。
(一)工作细胞的跨膜电位及其形成机制
1.静息电位和动作电位人和哺乳动物的心室肌细胞和骨骼肌细胞一样,在静息状态下膜两侧呈极化状态,膜内电位比膜外电位约低90mV,但两者的动作电位有明显不同。骨骼肌细胞动作电位的时程很短,仅持续几个毫秒,复极速度与去极速度几乎相等,记录曲线呈升支和降支基本对称的尖锋状。心室肌细胞动作电位的主要特征在于复极过程比较复杂,持续时间很长,动作电位降支与升支很不对称。通常用0、1、2、3、4等数字分别代表心室肌细胞动作电位和静息电位的各个时期。
(1)除极(去极)过程:除极过程又称0期。在适宜的外来刺激作用下,心室肌细胞发生兴奋,膜内电位由静息状态下的-90mV迅速上升到+30mV左右,即肌膜两侧原有的极化状态被消除并呈极化倒转,构成动作电位的升支。除极相很短暂,仅占1-2ms,而且除极幅度很大,为120mV;可见,心室肌细胞的除极速度很快,膜电位的最大变化速率可达800-1000V/s。
(2)复极过程:当心室细胞除极达到顶峰之后,立即开始复极,但整个复极过程比较缓慢,包括电位变化曲线的形态和形成机制均不相同的三个阶段:
1期复极:在复极初期,仅出现部分复极,膜内电位由+30mV迅速下降到0mV左右,故1期又称为快速复极初期,占时约10ms。0期除极和1期复极这两个时期的膜电位的变化速度都很快,记录图形上表现为尖锋状,故在心肌细胞习惯上常把这两部分合称为锋电位。
2期复极:当1期复极膜内电位达到0mV左右之后,复极过程就变得非常缓慢,膜内电位基本上停滞于0mV左右,细胞膜两侧呈等电位状态,记录图形比较平坦,故复极2期又称为坪或平台期,持续约100-150ms,是整个动作电位持续时间长的主要原因,是心室肌细胞以及其它心肌细胞的动作电位区别于骨骼肌和神经纤维的主要特征。
3期复极;2期复极过程中,随着时间的进展,膜内电位以较慢的速度由0mV逐渐下降,延续为3期复极,2期和3期之间没有明显的界限。在3期,细胞膜复极速度加快,膜内电位由0mV左右较快地下降到-90mV,完成复极化过程,故3期又称为快速复极末期,占时约100-150ms 。
4期:4期是膜复极完毕、膜电位恢复后的时期。在心室肌细胞或其它非自律细胞,4期内膜电位稳定于静息电位水平,因此,4期又可称为静息期。
2.形成机制与骨骼肌一样,离子在细胞膜两侧不均匀分布所形成的浓度梯度(浓度差)(表4-1)、驱动相应离子经过当时开放的细胞膜上特殊离子通道的跨膜扩散,是心肌细胞跨膜电位形成的主要基础,只是由于心肌细胞膜上具有数目较多的离子通道,跨膜电位形成机制中涉及的离子流远比骨骼肌要复杂得多。在电生理学中,电流的方向以正离子在膜两侧的流动方向来命名,正离子外流或负离子内流称外向电流,正离子内流或负离子外流称内向电流。外向电流导致膜内电位向负电性转化,促使膜复极,内向电流导致膜内电位向正电性转化,促使膜除极。
表4-1 心肌细胞中各种主要离子的浓度及平衡电位值
| 离 子 | 浓度(mmo1/L) 细胞内液 细胞外液 |
内/外比值 | 平衡电位(mV) (由Nernst公式计算) |
|
| Na+ | 30 | 140 | 1:4,6 | +41 |
| K+ | 140 | 4 | 35:1 | -94 |
| Ca2+ | 10-4 | 2 | 1:20,000 | +132 |
| CI- | 30 | 104 | 1:3.5 | -33 |
除离子跨膜扩散之外,由细胞上离子泵所实现的离子主动转运和离子交换,在心肌细胞电活动中也占有重要地位。
心室肌细胞静息电位的形成机制与骨骼肌相同,也就是说,尽管肌膜两侧上述几种离子都存在有浓度梯度,但静息状态下肌膜对K+的通透性较高,而对其它离子的通透性很低,因此,K+顺其浓度梯度由膜内向膜外扩散所达到的平衡电位,是静息电位的主要来源。
肌膜钠通道的大量开放和膜两侧浓度梯度及电位梯度的驱动从而出现Na+快速内流,是心室肌细胞0期去极形成的原因。进一步对整个去极过程进行分析就可以看到,与骨骼肌一样,在外来刺激作用下,首先引起部分电压门控式Na+通道开放和少量Na+内流,造成肌膜部分去极化,膜电位绝对值下降;而当膜电位由静息水平(膜内-90mV)去极化到阈电位水平(膜内-70mV)时,膜上Na+通道开放概率明显增加,出现再生性Na+内流(参看第二章),于是Na+顺其浓度梯度和电位梯度由膜外快速进入膜内,进一步使膜去极化,膜内电位向正电性转化。决定0期去极的Na+通道是一种快通道,它不但激活、开放的速度很快,而且激活后很快就失活,当膜除极到一定程度(omV左右)时,Na+通道就开始失活而关闭,最后终止Na+的继续内流。快Na+通道可被河豚毒(TTX)所阻断。由于Na+通道激活速度非常之快,又有再生性循环出现,这就是心室肌细胞0期去极速度很快,动作电位升支非常陡峭的原因。正因为如此,从电生理特性上,尤其是根据0期除极的速率,将心室肌细胞(以及具有同样特征的心肌细胞)称为快反应细胞,其动作电位称为快反应电位,以区别于以后将要介绍的慢反应细胞和慢反应电位。
复极1期是在0期除极之后出现的快速而短暂的复极期,此时快钠通道已经失活,同时激活一种一过性外向电流(Ito),从而使膜迅速复极到平台期电位水平(0~-20mV)。至于Ito的离子成分,70年代曾认为是Cl-(即Cl-内流)。近年来,根据Ito可被四乙基铵和4-氨基吡啶等K+通道阻滞剂所阻断的研究资料,认为K+才是Ito的主要离子成分。也就是说,由K+负载的一过性外向电流是动作电位初期快速复极的主要原因。目前对Ito的通道特征尚不十分清楚,但有资料提示,膜除极和细胞内Ca2+都可以使Ito的通道激活。
平台期初期,膜电位稳定于0mV左右,随后才非常缓慢地复极。膜电位的这种特征是由于平台期同时有内向电流和外向电流存在,初期,两种电流处于相对平衡状态,随后,内向电流逐渐减弱,外向电流逐渐增强,总和的结果是出现一种随时间推移而逐渐增强的、微弱的外向电流,导致膜电位缓慢地向膜内负电性转化。电压钳研究结果表明,在心室肌等快反应细胞,平台期外向离子流是由K+携带的(称Ik1)。静息状态下,K+通道的通透性很高,在0期除极过程中,K+的通透性显著下降,K+外流大大减少,除极相结束时,K+的通透性并不是立即恢复到静息状态下的那种高水平,而是极其缓慢地、部分地恢复,K+外流也就由初期的低水平而慢慢增加(图4-6)。平台期内向离子流主要是由Ca2+(以及Na+)负载的。已经证明,心肌细胞膜上有一种电压门控式的慢Ca2+通道,当膜除极到-40mV时被激活,Ca2+顺其浓度梯度向膜内缓慢扩散从而倾向于使膜除极,与此同时,上述微弱的K+外流倾向于使膜复极化,在平台期早期,Ca2+的内流和K+的外流所负载的跨膜正电荷时相等,膜电位稳定于1期复极所达到的电位水平。随着时间推移,Ca2+通道逐渐失活,K+外流逐渐增加,其结果,出膜的净正电荷量逐渐增加,膜内电位于是逐渐下降,形成平台期晚期。此后,Ca2+通道完全失活,内向离子流终止,外向K+流进一步增强,平台期延续为复极3期,膜电位较快地回到静息水平,完成复极化过程。
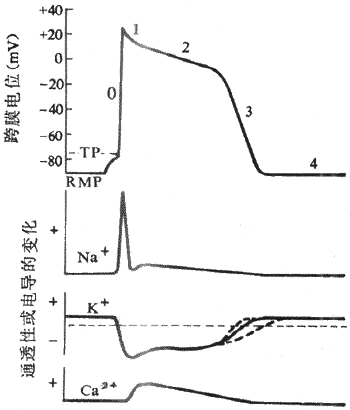
图4-6 心室肌细胞跨膜电位及其形成的离子机制
RMP:静息膜电位 TP :阈电位
肌膜上有Ca2+通道,是心室肌细胞和其它心肌细胞的重要特征。大量研究表明:①从一个心肌细胞的总体而言(不是从单个通道而言),Ca2+通道的激活、失活,以及再复活所需时间均比Na通道要长,经 Ca2+通道跨膜的Ca2+内流,起始慢,平均持续时间也较长。因此相应称为慢通道和慢内向离子流;②慢通道也是电压门控式的,激活慢通道的阈电位水平(-50~-35mV)高于快Na通道(-70~-55mV);③它对某些理化因素的敏感性和反应性不同于快通道,可被Mn2+和多种Ca2+阻断剂(如异博定,D-600等)所阻断,而对于可以阻断快通道的河豚毒和细胞膜的持续低极化状态(膜内电位-50Mv左右)却并不敏感。各种心肌细胞的肌膜上都具有这种慢通道,由此形成的跨膜离子流,是决定心肌细胞电活动以及心室肌等快反应细胞动作电位平台期的最重要的内向离子流之一。
平台期之后,膜的复极逐渐加速,因此时Ca2+通道已经失活,在平台期已经激活的外向K+流出现随时间而递增的趋势。其原因是,3期的复极K+流是再生性的,K+的外流促使膜内电位向负电性转化,而膜内电位越负,K+外流就越增高。这种正反馈过程,导致膜的复极越来越快,直至复极化完成。
在4期内,心室肌细胞膜电位基本上稳定于静息电位水平,但是,离子的跨膜转运仍然在活跃进行。因为,动作电位期间有Na+和Ca2+进入细胞内,而K+外流出细胞,因此,只有从细胞内排出多余的Na+和Ca2+,并摄入K+才能恢复细胞内外离子的正常浓度梯度,保持心肌细胞的正常兴奋性。这种离子转运是逆着浓度梯度进行的主动转运过程。像骨骼肌一样,通过肌膜上Na+-K+泵的作用,将Na+的外运和K+的内运互相耦联形成Na+-K+转运,同时实现Na+和K+的主动转运。关于主动转运Ca2+的转运机制,还没有完全弄清楚。目前大多数作者认为,Ca2+的逆浓度梯度的外运是与Na+的顺浓度的内流相耦合进行的。形成Na+-Ca2+交换。Ca2+的这种主动转运是由Na+的内向性浓度梯度提供能量的,由于Na+内向性浓度梯度的维持是依靠Na+-K+泵而实现的,因此,Ca2+主动转运也是由Na+-K+泵提供能量的。在4期开始后,膜的上述主动转运功能加强,细胞内外离子浓度梯度得以恢复。总的来看,这时转运过程引起的跨膜交换的电荷量基本相等,因此,膜电位不受影响而能维持稳定。
(二)自律细胞的跨膜电位及其形成机制
在没有外来刺激时,工作细胞不能产生动作电位,在外来刺激作用下,产生一次动作电位,但两次动作电位之间膜电位是稳定不变的。而在自律细胞,当动作电位3期复极未期达到最大值(称最大复极电位)之后,4期的膜电位并不稳定于这一水平,而是立即开始自动除极,除极达阈电位后引起兴奋,出现另一个动作电位。这种现象,周而复始,动作电位就不断地产生。出现于4期的这种自动除极过程,具有随时间而递增的特点,其除极速度远较0期除极缓慢;不同类型的自律细胞4期除极速度参差不一,但同类自律细胞4期除极速度比较恒定。这种4期自动除极(亦称4期缓慢除极或缓慢舒张期除极),是自律细胞产生自动节律性兴奋的基础。
根据细胞膜除极的跨膜电流的基本规律可分析自律细胞4期自动除极形成的机制。不难推测,自律细胞由于净外向电流使膜复极(3期)达最大复极电位后,在4期中又出现一种逐渐增强的净内向电流,从而使膜内正电位逐渐增加,膜便逐渐除极。这种进行性净内向电流的产生,有以下三种可能的原因:①内向电流的逐渐增强;②外向电流的逐渐衰退;③两者兼有。不同类型的自律细胞,4期自动除极都是由这种进行性净内向电流所引起,但构成净内向电流的离子流的方向和离子本质并不完全相同。
1.浦肯野细胞浦肯野细胞是一种快反应自律细胞。作为一种快反应型细胞,它的动作电位的形态与心室肌细胞相似,产生的离子基础也基本相同。
关于浦肯野细胞4期自动除极形成的机制,80年代研究资料表明,在浦肯野细胞,随着复极的进行,导致膜复极的外向K+电流逐渐衰减,而同时在膜电位4期可记录到一种随时间推移而逐渐增强的内向电流(If)(图4-7)。If通道在动作电位3期复极电位达-60mV左右开始被激活开放,其激活程度随着复极的进行、膜内负电性的增加而增加,至-100mV左右就充分激活。因此,内向电流表现出时间依从性增强,膜的除极程度因而也随时间而增加,一旦达到阈电位水平,便又产生另一次动作电位,与此同时,这种内向电流在膜除极达-50mV左右因通道失活而中止。可见,动作电位的复极期膜电位本身是引起这种内向电流启动和发展的因素,内向电流的产生和增强导致膜的进行性除极,而膜的除极一方面引起另一次动作电位,一方面又反过来中止这种内向电流。这一连串的过程是自律细胞“自我”启动、“自我”发展,又“自我”限制的,由此可以理解为什么自律细胞能够自动地、不断地产生节律性兴奋。
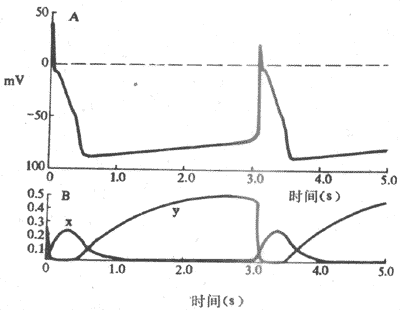
图4-7 浦肯野细胞起搏机制
A;跨膜电位 B:由х闸门控制的Ik衰减以及由у闸门控制的If,两者在形成起搏电位中的相对关系
这种4期内向电流,通常称为起搏电流,其主要离子成分为Na+,但也有K+参与。由于使它充分激活的膜电位为-100mV,因而认为,构成起搏内向电流的是一种被膜的超极化激活的非特异性内向(主要是是Na+)离子流,标志符号为If。If的通道允许Na+通过,但不同于快Na+通道,两者激活的电压水平不同;If可被铯(Cs)所阻断,而河豚毒却不能阻断它。目前,关于If及其通道的研究资料尚有若干不能充分予以解释的疑点,对If的进一步研究正受到心肌电生理学者们的高度关注。
2.窦房结细胞的跨膜电位及其形成机制 窦房结含有丰富的自律细胞,动作电位复极后出现明显的4期自动除极,但它是一种慢反应自律细胞,其跨膜电位具有许多不同于心室肌快反应细胞和浦肯野快反应自律细胞的特征:①窦房结细胞的最大复极电位(-70mV)和阈电位(-40mV)均高于(电位较正)浦肯野细胞;②0期除极结束时,膜内电位为0mV左右,不出现明显的极化倒转;③其除极幅度(70mV)小于浦肯野细胞(为120mV),而0期除极时程(7ms左右)却又比后者(1-2ms)长得多。原因是窦房结细胞0期除极速度(约10V/s)明显慢于浦肯野细胞(200-1000V/s),因此,动作电位升支远不如后者那么陡峭;④没有明显的复极1期和平台期;⑤4期自动除极速度(约0.1V/s)却比浦肯野细胞(约0.02V/s)要快,记录曲线上窦房结细胞4期膜电位变化的斜率大于浦肯野细胞.图4-8显示心室肌快反应细胞与窦房结细胞跨膜电位变化的差别。
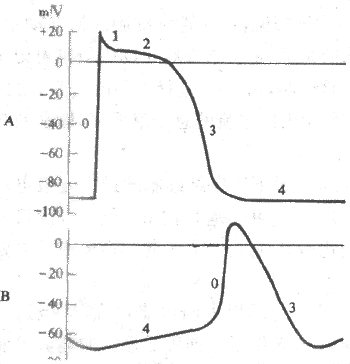
图4-8 心室肌(A)与窦房结(B)细胞跨膜电位的比较
窦房结细胞的直径很小,进行电生理研究有一定困难。直到70年代中期,才开始在窦房结小标本上采用电压钳技术对其跨膜离子流进行了定量研究,但目前尚未能充分阐明它的跨膜电位,尤其是4期起搏电流的离子基础。学者们观察到,窦房结细胞0期除极不受细胞外Na+浓度的影响,对河豚毒很不敏感;相反,它受细胞外Ca2+浓度的明显影响,并可被抑制钙通道的药物和离子(如异搏定、D-600和Mn2+等)所阻断。据此可以认为,引起窦房结细胞动作电位0期除极的内向电流是由Ca2+负载的。这种内向电流被称为第二内向电流;而引起快反应细胞(心室肌、心房肌和浦肯野细胞)0期除极的快Na+内流称为第一内向电流。根据已有的研究资料,可将窦房结细胞动作电位的形成过程描述如下:当膜电位由最大复极电位自动除极达阈电位水平时,激活膜上钙通道,引起Ca2+内向流(Ica),导致0期除极;随后,钙通道逐渐失活,Ca2+内流相应减少;另一方面,在复极初期,有一种K+通道被激活,出现K+外向流(Ik)。Ca2+内流的逐渐减少和K+外流的逐渐增加,膜便逐渐复极。由“慢”通道所控制、由Ca2+内流所引起的缓慢0期除极,是窦房结细胞动作电位的主要特征,因此,相应称为慢反应细胞和慢反应电位,以区别于前述心室肌等快反应细胞和快反应电位。
窦房结细胞的4期自动除极也由随时间而增长的净内向电流所引起,但其构成成分比较复杂,是几种跨膜离子流的混合。目前已知,在窦房结细胞4期可以记录到三种膜电流,包括一种外电流和两种内向电流,不过它们在窦房结细胞起搏活动中所起作用的大小以及起作用的时间有所不同。
(1)Ik通道的激活和逐渐增强所造成的K+外向流,是导致窦房结细胞复极的原因。Ik通道在膜复极达-40mV时便开始逐渐失活,K+外流因此渐渐减少,导致膜内正电荷逐渐增加而形成4期除极。目前认为,由于Ik通道的时间依从性逐渐失活所造成的K+外流进行性衰减,是窦房结细胞4期自动除极的最重要的离子基础(图4-9);
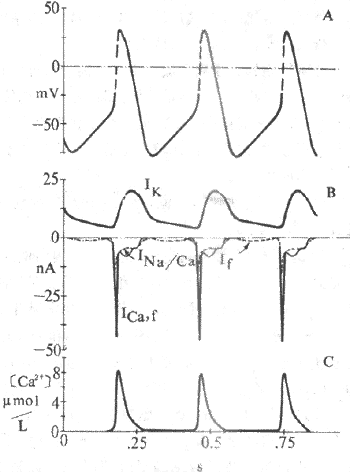
图4-9 窦房结动作电位和起搏电位的离子机制
A.跨膜电位 B.越膜电位C. 胞浆Ca2+浓度表示动作电位升支由Ica,
f构成,起搏电位 由Ik和If及Ina/Ca构成
(2)If:If是一种进行性增强的内向离子(主要为Na+)流,在浦肯野细胞起搏活动中,If起着极重要的作用,而Ik衰减的作用很小。与此恰相反,窦房结细胞4期虽也可记录到If ,但它对起搏活动所起的作用不如Ik衰减。实验证明,用Cs2+选择性阻断If后,窦房结自发放频率仅轻度减少;对家兔窦房结细胞4期净内向电流的总幅值而言,Ik衰减与If两者所起作用的比例为6:1。
If通道的最大激活电位为-100mV左右,而正常情况下窦房结细胞的最大复极电位为-70mV,在这种电位水平下,If通道的激活十分缓慢,这可能是If在窦房结4期除极过程中所起作用不大的原因。若窦房结细胞发生超级化时,If则可能成为起搏电流中的主要成分。
(3)此外,窦房结细胞4期中还存在一种非特异性的缓慢内向电流,在膜除极达-60mV时被激活,可见,它在自动除极过程的后1/3期间才起作用。这种缓慢内向电流可能是生电性Na+-Ca2+交换的结果(Na+-Ca2+交换时,心肌细胞排出一个Ca2+,摄入3个Na+,出/入细胞正电荷之比为2:3,形成内向电流)。
心肌细胞的类型 除了按照功能和电生理特性将心肌细胞分为工作细胞和自律细胞之外,还可以根据其生物活动的特征,特别是动作电位0期除极的速度,将心肌细胞分为快反应细胞和慢反应细胞,其动作电位相应称为快反应电位和慢反应电位;然后再结合其自律性,可将心肌细胞分为以下四种类型:
快反应非自律细胞:包括心房肌细胞和心室肌细胞;
快反应自律细胞:浦肯野自律细胞;
慢反应自律细胞:窦房结自律细胞,以及房结区和结希区的自律细胞;
慢反应非自律细胞:结区细胞。
心肌细胞的跨膜离子流 由多种离子通过细胞膜上的特异性或非特异性离子通道以及经离子交换转运形成的跨膜离子运动,是心肌细胞跨膜电位形成的基础。心肌细胞跨膜离子流的种类众多,性质很复杂,近年又取得许多进展。由于在实验研究中对这些离子通道的命名和认识比较混乱,甚至前后矛盾,故将目前比较肯定的主要几种离子流和通道的名称、存在的部位和作用归纳介绍如下:
1.内向离子流
(1)INa:称为快速Na+流,存在于快反应细胞,是引起快反应细胞0期除极的内向电流(称第一内向电流)的离子基础。
(2)Isi:称为缓慢内向电流,也称第二内向电流,过去认为它是一种由慢通道控制的缓慢Ca2+流,故标志为ICa,其功能是构成快反应细胞的平台期和慢反应细胞的除极期。新近的研究结果在很大程度上修正了对Ica的认识。目前认为,第二内向电流并不是单一的Ca2+流,而是由特性各异的三个组分所构成;第一组分称Ica.f,二、三组分别称Is1.2和Ica.s。其中①Ica.f是一种快速Ca2+流,其通道激活和失活的速度远比已往所认为的要快得多。它融合于快反应细胞Na+内流的最后部分,共同形成动作电位升支的上段,而对平台期的作用很小。它是慢反应细胞除极的离子基础。对工作细胞而言,肌浆网Ca2+再生性释放帅细胞外Ca2+进入胞浆触发的,既然Ica.f的激活十分迅速,那么Ca2+释放的触发有及兴奋-收缩耦联的启动速度都比以往认为的要快得多;②第三组分Ica.s,是一种较Ica.f微弱而缓慢的Ca2+流,主要作用是维持快反应细胞平台期;③Is1.2称第二内向电流第二组分,其离子本质不甚清楚,可能是Na+-Ca2+交换的生电电流,故也称为INa.Ca,在平台期起作用;慢反应自律细胞4期自动除极晚期也有它的作用。
(3)If:超极化激活的非特异性内向离子流,主要由Na+携带,存在于自律细胞4期。If是决定浦肯野快反应自律起搏活动的主要负离子流,而在窦房结慢反应自律细胞起搏活动中,其作用不如Ik衰减。
2.外向离子流
(1)Ikl:存在于快反应细胞,是决定快反应工作细胞静息电位的离子流,并在复极2期和3期起复极作用;
(2)Ik:这种外向电流主要由K+携带,但也有Na2+参加,不是单纯的K+流,故又称Ix,在快反应细胞复极3期起重要作用,故又称为复极电流,因而也决定着浦肯野细胞的最大复极电位;Ix也存在于慢反应自律细胞,促使膜复极,4期内呈进行性衰减,是形成4期除极的主要离子基础;
(3)Ito:是快反应细胞1期复极的离子流,离子成分主要为K+,也有Na+参与。
3.化学门控离子通道 近年相继证实,在心肌细胞膜上,除了电压门控通道外,还存在化学门控离子通道;它们的作用并不在于维持和产生正常跨膜电位(静息电位和动作电位),但对于心肌细胞活动的调节以及异常电活动的电生产有着特别重要的意义。重要的化学门控离子通道有以下三种;
(1)乙酰胆碱控制的K+通道(Ik-ACh):早年曾认为ACh激活的是Ikl,近年发现是通过G蛋白激活开放了另一种与在生物物理学特性和生理学特性上均不相同的K+通道。
(2)ATP依从性K+通道(Ik-ATP):ATP的作用并不是分解供能激活此通道,而是维持此通道在正常情况下处于关闭状态。当心肌细胞内ATP降到临界水平以下时(如心肌缺血时),此种特殊的K+通道开放。大量K+外漏以致缺血心肌细胞局部高钾而引起除极,诱发心律失常。硫脲类药物可阻断此通道。
(3)INa–k—Ca2+通道:是细胞内Ca2+增高时激活的一种非特异性正离子通道,载流离子是Na+和K+,形成一过性内向离子流(Iti)。实验表明,在某些情况下,浦肯野细胞在动作电位复极后可产生一种除极电位(延迟后除极电位),当它达阈电位时就可以诱发另一个新的动作电位,形成异位搏动。Iti就是延迟后除极电位的离子基础。洋地黄中毒,细胞外低K+或低Na+,以及咖啡因、儿茶酚胺等可引起细胞内Ca2+超负荷的因素,均可诱发或加强Iti和延迟后除极电位。
二、心肌的电生理特性
心肌组织具有兴奋性、自律性、传导性和收缩性四种生理特性。心肌的收缩性是指心肌能够在肌膜动作电位的触发下产生收缩反应的特性,它是以收缩蛋白质之间的生物化学和生物物理反应为基础的,是心肌的一种机械特性。兴奋性、自律性和传导性,则是以肌膜的生物电活动为基础的,故又称为电生理特性。心肌组织的这些生理特性共同决定着心脏的活动。
(一)心肌的兴奋性
所有心肌细胞都具有兴奋性,即具有在受到刺激时产生兴奋的能力。衡量心肌的兴奋性,同样可以采用刺激的阈值作指标,阈值大表示兴奋性低,阈值小表示兴奋性高。
1.决定和影响兴奋性的因素 从关于兴奋产生过程的叙述中可知,兴奋的产生包括静息电位去极化到阈电位水平以及Na+通道(以快反应型细胞为例)的激活这样两个环节;当这两方面的因素发生变化时,兴奋性将随之发生改变。
(1)静息电位水平:静息电位(在自律细胞,则为最大复极电位)绝对值增大时,距离阈电位的差距就加大,引起兴奋所需的刺激阈值增大,表现为兴奋性降低。反之,静息电位绝对值减少时,距阈电位的差距缩小,所需的刺激阈值减少,兴奋性增高。
(2)阈电位水平:阈电位水平上移,则和静息电位之间的差距增大,引起兴奋所需的刺激阈值增大,兴奋性降低。反之亦然。
静息电位水平和(或)阈电位水平的改变,都能够影响兴奋性,但在心脏,以静息电位水平改变为多见的原因。
(3)Na+通道的性状:上述兴奋的产生时,都是以Na+通道能够被激活作为前提。事实上,Na+通道并不是始终处于这种可被激活的状态,它可表现为激活、失活和备用三种功能状态:而Na+通道处于其中哪一种状态,则取决于当时的膜电位以及有关的时间进程。这就是说,Na+通道的活动是电压依从性和时间依从性的。当膜电位处于正常静息电位水平-90mV时,Na+通道处于备用状态。这种状态下,Na+通道具有双重特性,一方面,Na+通道是关闭的;另一方面,当膜电位由静息水平去极化到阈电位水平(膜内-70mV)时,就可以被激活,Na+通道迅速开放,Na+因而得以快速跨膜内流。Na+通道激活后就立即迅速失活,此时通道关闭,Na+内流迅速终止。Na+通道的激活和失活,都是比较快速的过程;前者在1ms 内,后者约在几毫秒到10ms内即可完成。处于失活状态的Na+通道不仅限制了Na+的跨膜扩散,并且不能被再次激活;只有在膜电位恢复到静息电位水平时,Na+通道才重新恢复到备用状态,即恢复再兴奋的能力,这个过程称为复活。由上可见,Na+通道是否处备用状态,是该心肌细胞当时是否具有兴奋性的前提;而正常静息膜电位水平又是决定Na+通道能否处于或能否复活到备用状态的关键。Na+通道的上述特殊性状,可以解释有关心肌细胞兴奋性的一些现象。例如,当膜电位由正常静息水平(-90mV)去极化到阈电位水平(-70mV)时,Na+通道被激活,出现动作电位;而如果静息状况下膜电位为-50mV左右,即肌膜处于持续低极化状态时,就不能引起Na+通道激活,表现为兴奋性的丧失。至于Na+通道上述三种状态的实质以及膜电位是如何影响Na+通道性状的问题,目前尚未彻底阐明。
2.一次兴奋过程中兴奋性的周期性变化心肌细胞每产生一次兴奋,其膜电位将发生一系列有规律的变化,膜通道由备用状态经历激活、失活和复活等过程,兴奋性也随之发生相应的周期性改变。兴奋性的这种周期性变化,影响着心肌细胞对重复刺激的反应能力,对心肌的收缩反应和兴奋的产生及传导过程具有重要作用。心室肌细胞一次兴奋过程中,其兴奋性的变化可分以下几个时期(图4-10):
(1)有效不应期:心肌细胞发生一次兴奋后,由动作电位的去极相开始到复极3期膜内电位达到约-55mV这一段时期内,如果再受到第二个刺激,则不论刺激有多强,肌膜都不会进一步发生任何程度的去极化;膜内电位由-55mV继续恢复到约-60mV这一段时间内,如果给予的刺激有足够的强度,肌膜可发生局部的部分去极化,但并不能引起扩播性兴奋(动作电位)。心肌细胞兴奋后不能立即再产生第二次兴奋的特性,称为不应性,不应性表现为可逆的、短暂的兴奋性缺失或极度下降。心肌细胞一次兴奋过程中,由0期开始到3期膜内电位恢复到-60mV这一段不能再产生动作电位的时期,称为有效不应期。其原因是这段时间内膜电位绝对值太低,Na+通道完全失活(前一阶段),或刚刚开始复活(后一阶段),但还远远没有恢复到可以被激活的备用状态的缘故。
(2)相对不应期:从有效不应期完毕(膜内电位约-60mV)到复极化基本上完成(约-80mV)的这段期间,为相对不应期。这一时期内,施加给心肌细胞以高于正常阈值的强刺激,可以引起扩播性兴奋。出现相对不应期的原因是:此期膜电位绝对值高于有效不应期末时的膜电位,但仍低于静息电位,这时Na+通道已逐渐复活,但其开放能力尚未恢复正常;故心肌细胞的兴奋性虽比有效不应期时有所恢复,但仍然低于正常,引起兴奋所需的刺激阈值高于正常,而所产生的动作电位(称期前兴奋)0期的幅度和速度都比正常为小,兴奋的传导也比较慢。此外,此期处于前一个动作电位的3期,尚有K+迅速外流的趋势,因此,在此期内新产生的动作电位,其时程较短(K+外流可使平台期缩短),不应期也较短。
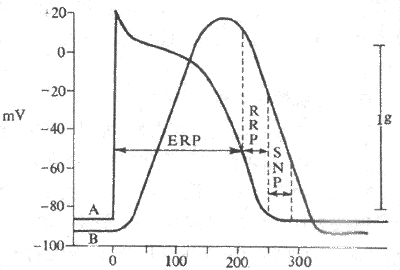
图4-10 心室肌动作电位期间兴奋性的变化及其与机械收缩的关系
A:动作电位 B:机械收缩ERP:有效不应期
RRP:相对不应期 SNP:超常期
(3)超常期:心肌细胞继续复极,膜内电位由-80mV恢复到-90mV这一段时期内,由于膜电位已经基本恢复,但其绝对值尚低于静息电位,与阈电位水平的差距较小,用以引起该细胞发生兴奋所需的刺激阈值比正常要低,表明兴奋性高于正常,故称为超常期。另一方面,此时Na+通道基本上恢复到可被激活的正常备用状态,但开放能力仍然没有恢复正常,产生的动力电位的0期去极的幅度和速度,兴奋传导的速度都仍然低于正常。
最后,复极完毕,膜电位恢复正常静息水平,兴奋性也恢复正常。
3.兴奋过程中,兴奋性周期性变化与收缩活动的关系细胞在发生一次兴奋过程中,兴奋性发生周期性变化,是所有神经和肌组织共同的特性;但心肌细胞的有效不应期特别长,一直延续到机械反应的舒张期开始之后。因此,只有到舒张早期之后,兴奋性变化进入相对不应期,才有可能在受到强刺激作用时产生兴奋和收缩。从收缩开始到舒张早期之间,心肌细胞不会产生第二个兴奋和收缩。这个特点使得心肌不会像骨骼肌那样产生完全强直收缩而始终作收缩和舒张相交替的活动,从而使心脏有血液回心充盈的时期,这样才可能实现其泵血功能。
以下实验可以说明心肌组织的这一特点。正常情况下,窦房结产生的每一次兴奋传播到心房肌或心室肌的时间,都是在它们前一次兴奋的不应期终结之后,因此,整个心脏能够按照窦房结的节律而兴奋。但在某些情况下,如果心室在有效不应期之后受到人工的或窦房结之外的病理性异常异常刺激,则可产生一次期前兴奋,引起期前收缩或额外收缩。期前兴奋也有它自己的有效不应期,这样,当紧接在期前兴奋之后的一次窦房结兴奋传到心室肌时,常常正好落在期前兴奋的有效不应期内,因而不能引起心室兴奋和收缩,形成一次“脱失”,必须等到再下一次窦房结的兴奋传到心室时才能引起心室收缩。这样,在一次期前收缩之后往往出现一段较长的心室舒张期,称为代偿性间歇(图4-11)。随之,才恢复窦性节律。
(二)心肌的自动节律性
组织、细胞能够在没有外来刺激的条件下,自动地发生节律性兴奋的特性,称为自动节律性,简称自律性。具有自动节律性的组织或细胞,称自律组织或自律细胞。组织、细胞单位时间(每分钟)内能够自动发生兴奋的次数,即自动兴奋的频率,是衡量自动节律性高低的指标。
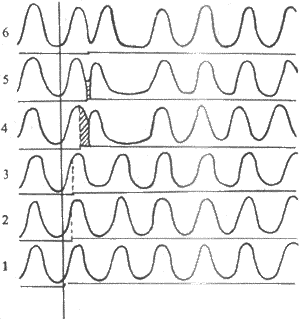
图4-11 期前收缩和代偿性间歇
每条曲线下的电磁标记号批示给予电刺激的时间,曲线1-3,
刺激落在有效不应期内,不引起反应;曲线4-6,
刺激落在相对不应期内,引起期前收缩和代偿性间歇
1.心肌的自动节律性和各自律级组织的相互关系很早以前就有人观察到,在适宜条件下,两栖类和哺乳类动物的离体心脏,在未受到任何刺激的情况下,可以长时间地、自动地、有节奏地进行兴奋和收缩。但是,只有到了近代,根据细胞内微电极技术记录的跨膜电位是否具有4期自动去极化这一特征,才确切地证明,并不是所有心肌细胞,而只是心脏特殊传导组织内某些自律细胞才具有自动节律性。特殊传导系统各个部位(结区除外)的自律性有等级差别;其中窦房结细胞自律性最高,自动兴奋频率约为每分钟100次,末梢浦肯野纤维网自律性最低(约每分钟25次),而房室交界(约每分钟50次)和房室束支的自律性依次介于两者之间。
由一个起搏点主宰整个心脏的整体活动具有极其重要的生理意义。那么,各部分自律组织的活动怎么能统一起来而不致于“各自为政”呢?实验中很容易观察到,心脏始终是依照当时情况下自律性最高的部位所发出的兴奋来进行活动的。这就是说,各部分的活动统一在自律性最高部位的主导作用之下。正常情况下,窦房结的自律性最高,它自动产生的兴奋向外扩布,依次激动心房肌、房室交界、房室束、心室内传导组织和心室肌,引起整个心脏兴奋和收缩。可见,窦房结是主导整个心脏兴奋和跳动的正常部位,故称为正常起搏点。其它部位自律组织并不表现出它们自身的自动节律性,只是起着兴奋传导作用,故称为潜在起搏点。在某种异常情况下,窦房结以外的自律组织(例如,它们的自律性增高,或者窦房结的兴奋因传导阻滞而不能控制某些自律组织)也可能自动发生兴奋,而心房或心室则依从当时情况下节律性最高部位的兴奋而跳动,这些异常的起搏部位则称为异位起搏点。
窦房结对于潜在起搏点的控制,通过两种方式实现:①抢先占领。窦房结的自律性高于其它潜在起搏点,所以,在潜在起搏点4期自动去极尚未达到阈电位水平之前,它们已经受到窦房结发出并依次传布而来的兴奋的激动作用而产生了动作电位,其自身的自动兴奋就不可能出现;②超速压抑或超速驱动压抑(overdrive suppression)。窦房结对于潜在起搏点,还可产生一种直接的抑制作用。例如,当窦房结对心室潜在起搏点的控制突然中断后,首先会出现一段时间的心室停搏,然后心室才能按其自身潜在起搏点的节律发生兴奋和搏动。出现这个现象的原因是:在自律性很高的窦房结的兴奋驱动下,潜在起搏点“被动”兴奋的频率远远超过它们本身的自动兴奋频率。潜在起搏长时间的“超速”兴奋的结果,出现了抑制效应;一旦窦房结的驱动中断,心室潜在起搏点需要一定的时间才能从被压抑状态中恢复过来,出现它本身的自动兴奋。另外还可以看到,超速压抑的程度与两个起搏点自动兴奋频率的差别呈平行关系,频率差别愈大,抑制效应愈强,驱动中断后,停搏的时间也愈长。因此,当窦房结兴奋停止或传导受阻后,首先由房室交界代替窦房结作为起搏点,而不是由心室传导组织首先代替;因为窦房结和房室交界的自动兴奋频率差距较小,超速压抑的程度较小。超速压抑产生的机制比较复杂,目前尚未完全弄清;但这一事实提示我们,在人工起搏的情况下,如因故需要暂时中断起搏器时,在中断之前其驱动频率应该逐步减慢,以避免发生心搏暂停。
2.决定和影响自律性的因素自律细胞的自动兴奋,是4期膜自动去极化使膜电位从最大复极电位达到阈电位水平而引起的。因此,自律性的高低,既受最大复极电位与阈电位的差距的影响,也取决于4期膜自动去极的速度(图4-12)。
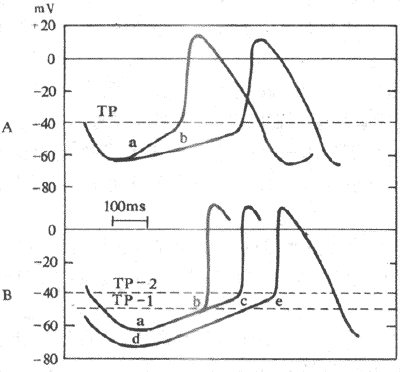
图4-12 影响自律性的因素
A:起搏电位斜率由a减少到b时,自律性降低
B:最大复极电位水平由a达到d,或阈电位由TP-1升到TP-2时,自律性均降低 Tp :阈电位
(1)最大复极电位与阈电位之间的差距:最大复极电位绝对值减少和(或)阈电位下移,均使两者之间的差距减少,自动去极化达到阈电位水平所需时间缩短,自律性增高;反之亦然。例如,迷走神经系统兴奋时可使窦房结自律细胞K+通道开放率增高,故其复极3期内K+外流增加,最大复极电位绝对值增大,自律性降低,心率减慢。
(2)4期自动除极速度:4期自动除极速度与膜电位从最大复极电位水平达到阈电梯水平所需时间密切相关;若除极速度增快,达阈电位水平所需时间缩短,单位时间内发生兴奋的次数增多,自律性增高。从前一节已知,4期自动除极速度取决于净内向电流增长的速度,即取决于膜内净正电荷增长速度。例如,儿茶酚胺可以增强If,因而加速浦肯野细胞4期除极速度,提高其自律性。
(三)心肌的传导性和心脏内兴奋的传导
心肌在功能上是一种合胞体,心肌细胞膜的任何部位产生的兴奋不但可以沿整个细胞膜传播,并且可以通过闰盘传递到另一个心肌细胞,从而引起整块心肌的兴奋和收缩。动作电位沿细胞膜传播的速度可作为衡量传导性的指标。
1.心脏内兴奋传播的途径和特点 正常情况下窦房结发出的兴奋通过心房肌传播到整个右心房和左心房,尤其是沿着心房肌组成的“优势传导通路”迅速传到房室交界区,经房室束和左、右束支传到浦肯野纤维网,引起心室肌兴奋,再直接通过心室肌将兴奋由内膜侧向外膜侧心室肌扩布,引起整个心室兴奋。由于各种心肌细胞的传导性高低不等,兴奋在心脏各个部分传播的速度是不相同的。在心房,一般心房肌的传导速度较慢(约为0.4m/s),而“优势传导通路”的传导速度较快,窦房结的兴奋可以沿着这些通路很快传播到房室交界区。在心室,心室肌的传导速度约为1m/s,而心室内传导组织的传导性却高得多,末梢浦肯野纤维传导速度可达4m/s,而且它呈网状分布于心室壁,这样,由房室交界传入心室的兴奋就沿着高速传导的浦肯野纤维网迅速而广泛地向左右两侧心室壁传导。很明显,这种多方位的快速传导对于保持心室的同步收缩是十分重要的。房室交界区细胞的传导性很低,其中又以结区最低,传导速度仅0.02m/s。房室交界是正常时兴奋由心房进入心室的唯一通道,交界区这种缓慢传导使兴奋在这里延搁一段时间(称房-室延搁)才向心室传播,从而可以使心室在心房收缩完毕之后才开始收缩,不致于产生房室收缩重叠的现象。可以看出,心脏内兴奋传播途径的特点和传导速度的不一致性,对于心脏各部分有次序地、协调地进行收缩活动,具有十分重要的意义。
2.决定和影响传导性的因素 心肌的传导性取决于心肌细胞某些结构特点和电生理特性。
(1)结构因素:细胞直径与细胞内电阻呈反变关系,直径小的细胞内电阻大,产生的局部电流小于粗大的细胞,兴奋传导速度也较后者缓慢。心房肌、心室肌和浦肯野细胞的直径大于窦房结和房室交界细胞,其中,末梢浦肯野细胞的直径最大(在某些动物,直径可达70μm),兴奋传导速度最快;窦房结细胞直径很小(约5-10μm),传导速度很慢;而结区细胞直径更小,传导速度也最慢。
在机体生命过程中,心肌细胞直径不会突然发生明显的变化,因此,它只是决定传导性的一个比较固定的因素,对于各种生理或某些病理情况下心肌传导性的变化,不起重要作用。
(2)生理因素:心肌细胞的电生理特性是决定和影响心肌传导性的主要因素。与其它可兴奋细胞相同,心肌细胞兴奋的传播也是通过形成局部电流而实现的(参看第二章)。因此,可以从局部电流的形成和邻近未兴奋部位膜的兴奋性这两方面来分析影响传导性的因素。这两方面因素是密切相关联的,为了方便才分别叙述。
动作电位0期除极的速度和幅度局部电流是兴奋部位膜0期去极所引起的,0期去极的速度愈快,局部电流的形成也就愈快,很快就促使邻近未兴奋部位膜去极达到阈电位水平,故兴奋传导愈快。另一方面,0期去极幅度愈大,兴奋和未兴奋部位之间的电位差愈大,形成的局部电流愈强,兴奋传导也愈快。问题是,为什么局部电流的强度能影响传导速度?可能是强的局部电流扩布的距离大,可以使距兴奋部位更远的下游部位受到局部电流的刺激而兴奋,故兴奋的传导较快。除了细胞直径这个因素之外,浦肯野纤维等快反应细胞0期去极速度和幅度明显高于窦房结等慢反应细胞,是前者传导性比后者为高的主要原因。
已知,各种心肌细胞0期去极速度和幅度的差别,主要由膜上(0期)离子通道的固有性质决定。那么,同一心肌细胞(以快反应细胞为例)0期去极速度和幅度又受什么因素的影响?在叙述兴奋性时已经指出,快Na+通道的性状,即激活、失活和复活状态是决定兴奋性正常、缺失和低下的主要因素;也就是说,对兴奋性而言,Na+通道的性状决定着通道能否被激活开放(兴奋性的有无)以及激活的难易程度(兴奋性的高低)。这里,将进一步讨论,Na+通道的性状还决定着膜去极达阈电位水平后通道开放的速度和数量,从而决定膜0期去极的速度和幅度。Na+通道开放速度和数量这种性状,称为Na+通道的效率或可利用率(通道开放数量称开放概率)。实验证明,Na+通道的效率也是电压依从性的,它依从于临受刺激前的膜静息电位值。定量地分析Na+通道的效率(用0期去极的最大速率反映Na+通道开放的速度)与静息膜电位值的函数关系的曲线为膜反应曲线(图4-13)。膜反应曲线呈S形。正常静息电位值(-90mV)情况下,膜受刺激去极达阈电位水平后,Na+通道快速开放,0期去极最大速度可达500V/s。如膜静息电位值(绝对值)降低,去极最大速度下降;若膜静息电位值(绝对值)进一步降低到膜内为-60~-55mV时,去极速度几乎为0,即Na+通道已失活而不能开放。上述这种现象称为Na+通道效率的电压依从性下降。需要引起注意的是,在静息膜电位值(绝对值)很低(膜内-60~-55mV)状况下,如果膜受到刺激,并不是根本不产生电位变化,而是产生一种0期去极速度和幅度都很小的动作电位。这是因为,在这种情况下快Na+通道已经失活,而慢Ca2+通道未受影响,因此,原来的快反应细胞此时出现了由Ca2+内流所致的慢反应电位的缘故;兴奋传导速度也就明显减慢。不过,这已经是膜0期去极的离子通道发生了更换,不再属于Na+通道效率的量变范畴。
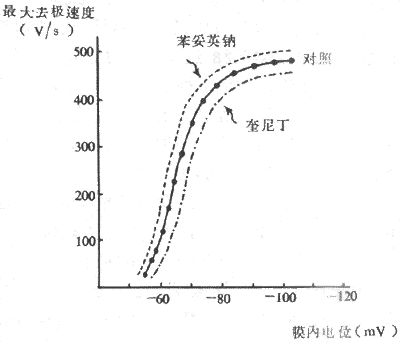
图4-13 膜反应曲线
除了静息膜电位之外,Na+通道开放的速度还受心肌细胞本身生理性质的影响。例如,苯妥英钠可使膜反应曲线左上移位,奎尼丁使之右下移位。这表明,在这些药物作用下,Na+通道开放效率仍然是电压依从性的,但是,同一静息膜电位水平的0期去极最大速度的数值并不相同,前者高于正常,后者低于正常。
膜反应曲线只描述了静息膜电位值对Na+通道开放速度即0期去极速度的影响,实际上,由Na+通道开放数量所决定的0期去极幅度也同样依从于静息膜电位值。正常静息膜电位情况下,Na+通道不但开放速度快,而且开放数量也多,动作电位0期去极的速度快,幅度也高(图4-14,左);若静息膜电位值(绝对值)低下,则产生升支缓慢、幅度低的动作电位(图4-14右)。
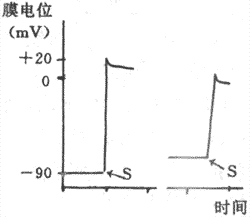
图4-14 静息膜电位对动作电位升支速度和幅度的影响
S:给予刺激
邻近未兴奋部位膜的兴奋性 兴奋的传导是细胞膜依次兴奋的过程,因此,膜的兴奋性必然影响兴奋的传导。前已述:①静息膜电位(或最大复极电位)与阈电位的差距及②邻近未兴奋部位膜上决定0期去极的离子通道的性状,是决定兴奋性从而也是影响传导性的主要因素。当差距扩大时,兴奋性降低(所需刺激阈值增高),同时,膜去极达阈电位水平所需时间延长,传导速度因此减慢。如在邻近部位形成额外刺激产生期前兴奋的情况,由兴奋部位形成的局部电流刺激就将在期前兴奋复极完成之前到达邻近部位,如落在期前兴奋的有效不应期内,则不能引起兴奋,导致传导阻滞;如落在期前兴奋的相对不应期或超常期内,可引起升支缓慢、幅度小的动作电位,兴奋传导因之减慢。可见,不应期的存在,是可能导致兴奋传导障碍的重要因素。
三、自主神经对心肌生物电活动和收缩功能的影响
支配心脏的自主神经及其递质对心肌生物电活动和收缩功能均产生明显影响,它们对心肌生物电活动和电生理特性的影响,主要是通过调节离子通道的开放而实现的,而对心肌收缩功能的调节机制则比较复杂;收缩功能的改变,除了是它们引起生物电改变的继发效应之外,还可能通过其它机制对收缩功能产生直接影响。
(一)迷走神经和乙酰胆碱的作用
迷走神经兴奋时,节后纤维释放递质乙酰胆碱,激动心肌细胞膜上M型胆碱能受体,产生负性压力、负性变时和负性传导性等效应。研究证明,乙酰胆碱能普遍提高K+通道的开放概率,促进外向K+流,是迷走神经心肌效应的主要机制。
K+外流的普遍增加将影响心肌细胞生物电活动的多个环节:①静息状态下K+外流的增加将导致静息电位绝对值增大;因此,静息电位与阈电位的差距扩大,心肌兴奋性有所下降;②在窦房结细胞,复极过程中K+外流增加的结果是最大复极电位绝对值增大;另一方面,其4期K+外流的增加将使Ik衰减过程减弱,自动除极速度减慢。这两方面因素均导致窦房结自律性降低,心率因而减慢;③复极过程中K+外流增加导致复极加速,动作电位时程缩短,有效不应期相应缩短,由于动作电位时程缩短,每一动作电位期间进入细胞内Ca2+量相应减少;除此之外,近年研究还发现,乙酰胆碱有直接抑制Ca2+通道、减少内向Ca2+流的作用。由于进入细胞内Ca2+量减少,心肌收缩能力相应降低,表现出负性变力效应。此外,当左侧迷走神经兴奋时,房室交界慢反应细胞动作电位幅度减小,兴奋传导速度减慢,这也是乙酰胆碱抑制Ca2+通道、减少Ca2+内流的结果。
(二)心交感神经和儿茶酚胺的作用
心交感神经末梢释放的递质是去甲肾上腺素,它与心肌细胞膜β型肾上腺素能受体相结合,通过某种机制(使通道蛋白质磷酸化)改变膜上离子通道的开放概率和其它亚细胞结构的功能,产生正性变力、正性变时和正性变传导性效应。肾上腺髓质分泌的去甲肾上腺素和肾上腺素,以及外源性β受体激动剂也有类似作用。乙酰胆碱心肌效应的机制相对比较单一,它主要是通过普遍提高膜的K+通道的开放概率而发挥作用的;儿茶酚胺的作用比较复杂,它除了能明显增加Ca2+通道的开放概率这一主要作用之外,还通过其它多种细胞和亚细胞机制调节心肌的电生理特性和收缩功能。其具体作用和作用机制归纳如下:①儿茶酚胺(去甲肾上腺素、肾上腺素)能加强自律细胞4期的跨膜内向电流If使4期自动除极速度加快,自律性增高;②在慢反应细胞,由于0期Ca2+内流加强加速,其动作电位上升速度和幅度均增加,房室交界区兴奋传导速度加快;③儿茶酚胺能使复极相K+外流增快,从而使复极过程加快,复极相因此缩短,不应期相应缩短。不应期缩短意味着0期离子通道复活过程加快,这与儿茶酚胺使窦房结兴奋发放频率增加的作用互相协调,使心率得以增加;④儿茶酚胺通过加强心肌收缩能力增强加速心肌的收缩,也加速心肌的舒张,其作用机制比较复杂。很明显,儿茶酚胺提高肌膜和肌浆网Ca2+通道开放概率的特性将导致细胞内Ca2+浓度增高,提供了促使心肌收缩能力增强的条件;另一方面,儿茶酚胺又促使肌钙蛋白对Ca2+亲和力下降,从而减弱心肌收缩能力。由于儿茶酚胺的前一种作用强于后者,因此,尽管它对心肌收缩能力的两种作用的方向相反,表现出的最终效果仍是强有力的正性变力作用。在儿茶酚胺作用下,心肌舒张速度增快,整个舒张过程明显加强,这是儿茶酚胺很具有特征性的一种效应。这种效应是三方面作用的结果:首先,儿茶酚胺促使肌钙蛋白对Ca2+的释放速率增加,这是因为儿茶酚胺降低了肌钙蛋白的Ca2+亲和力,Ca2+可以很快从与肌钙蛋白结合的状态下解离出来;其次,儿茶酚胺可以提高肌浆网重摄取Ca2+的速度,并可刺激Na+-Ca2+交换,使复极期向细胞排出Ca2+加速。综合上述,在儿茶酚胺作用下,舒张期胞浆Ca2+浓度下降速度增快,肌钙蛋白又很快与Ca2+解离,这些作用都有利于肌原纤维中粗、细肌丝的离解,心肌因而以较快的速度完成舒张过程。
心交感神经和儿茶酚胺对心肌的作用是多方面的,很复杂的,但这些效应是互相统一协调的。在心交感神经兴奋引起心率加快,从而使心缩期和心舒期都缩短的情况下,心肌舒张加速可以弥补因心室舒张期缩短带来的心室充盈不足的后果。另一方面,由于收缩能力增强,心肌收缩增强、增快,因此,心脏射血量也不致于因心缩期的缩短而减少;再加上兴奋传导加速,心房肌以及心室肌收缩的同步性有所加强,也促使心房或心室的收缩强度增加,有利于维持心输出量。这样,当心交感神经兴奋时,在心率加快的同时,搏出量增加或不变,心输出量因而得以大大增加。
四、体表心电图
在正常人体,由窦房结发出的一次兴奋,按一定的途径和进程,依次传向心房和心室,引起整个心脏的兴奋;因此,每一个心动周期中,心脏各部分兴奋过程中出现的电变化传播方向、途径、次序和时间等都有一定的规律。这种生物电变化通过心脏周围的导电组织和体液,反映到身体表面,使身体各部位在每一心动周期中也都发生有规律的电变化。将测量电极放置在人体表面的一定部位记录出来的心脏电变化曲线,就是临床上记录的心电图(electrocardiogram,ECG)。心电图反映心脏兴奋的产生、传导和恢复过程中的生物电变化,而与心脏的机械收缩活动无直接关系。
心肌细胞的生物电变化是心电图的来源,但是,心电图曲线与单个心肌细胞的生物电变化曲线有明显的区别(图4-15)。造成这种区别的主要原因有以下几点:①单个心肌细胞电变化是用细胞内电极记录法得到的,即一个测量电极放在细胞外表面而另一个电极插入到细胞膜内,所测到的电变化是同一细胞的膜内外的电位差,它不仅可测出膜的动作电位,也可测出膜的静息电位。心电图的记录方法原则上属于细胞外记录法,它只能测出已兴奋部位和尚处于兴奋状态的部位之间的电位差。在静息状态下,或是肌膜各部位都处于兴奋状态下时,膜外各部位之间没有电位差,细胞外记录曲线都将呈等电位线,不能加以区别;②心肌细胞电变化曲线是单个心肌细胞在静息时或兴奋时膜内外电位变化曲线;而心电图反映的是一次心动周期中整个心脏的生物电变化,因此,心电图上每一瞬间的电位数值,都是很多心肌细胞电活动的综合效应在体表的反映;③与细胞内记录法不同,心电图是在身体表面间接记录的心脏电变化,因此,电极放置的位置不同,记录的心电图曲线也不相同。
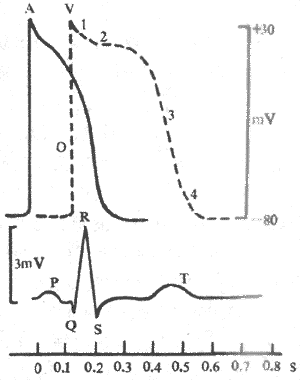
图4-15心肌细胞电变化曲线与常规心电图的比较
A:心房肌细胞电变化 V:心室肌细胞电变化
正常典型心电图的波形及其生理意义 心电图记录纸上有横线和纵线划出长和宽均为1mm的小方格。记录心电图时,首先调节仪器放大倍数,使输入1mV电压信号时,描笔在纵向上产生10mm偏移,这样,纵线上每一小格相当于0.1mV的电位差。横向小格表示时间,每一小格相当于0.04s(即走纸速度为每秒25mm)。因此,可以在记录纸上测量出心电图各波的电位数值和经历的时间。
测量电极安放位置和连线方式(称导联方式)不同所记录到的心电图,在波形上有所不同,但基本上都包括一个P波,一个QRS波群和一个T波,有时在T波后,还出现一个小的U波(图4-16)。
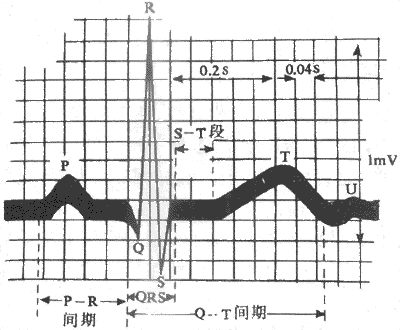
图4-16 正常人心电模式图
1.P波 反映在左右两心房的去极化过程。P波波形小而圆钝,历时0.08-0.11s,波幅不超过0.25mV。
2.Ta波(心房T波)代表心房复极过程所产生的电变化。它开始于P波之后,与P波的方向相反。P-Ta间期(从P波开始到Ta波终了的时程)为0.15-0.45s;故Ta波与P-R段、QRS波和ST段的初期重叠在一起,而且Ta波波幅很低,故通常心电图上看不出Ta波。
3.QRS波群代表左右两心室去极化过程的电位变化。典型的QRS波群,包括三个紧密相连的电位波动:第一个向下波为Q波,以后是高而尖峭的向上的R波,最后是一个向下的S波。但在不同导联中,这三个波不一定都出现。正常QRS波群历时约0.06-0.10s,代表心室肌兴奋扩布所需的时间;各波波幅在不同导联中变化较大。
4.T波 反映心室复极(心室肌细胞3期复极)过程中的电位变化,波幅一般为0.1-0.8mV。在R波较高的导联中T波不应低于R波的1/10。T波历时0.05-0.25s。T波的方向与QRS波群的主波方向相同。
5.U波 是T波后0.02-0.04s可能出现的一个低而宽的波;方向一般与T波一致,波宽约0.1-0.3s,波幅大多在0.05mV以下。U波的意义和成因均不十分清楚。
在心电图中,除了上述各波的形状有特定的意义之外,各波以及它们之间的时程关系也具有理论和实践意义。其中比较重要的有以下几项:
1.PR间期(或PQ间期) 是指从P波起点到QRS波起点之间的时程,为0.12-0.20s。PR间期代表由窦房结产生的兴奋经由心房、房室交界和房室束到达心室,并引起心室开始兴奋所需要的时间,故也称为房室传导时间;在房室传导阻滞时,PR间期延长。
2.PR段 从P波终点到QRS波起点之间的曲线,通常与基线同一水平。PR段形成的原因是由于兴奋冲动通过心房之后在向心室传导过程中,要通过房室交界区;兴奋通过此区传导非常缓慢,形成的电位变化也很微弱,一般记录不出来,故在P波之后,曲线又回到基线水平,成为PR段。
3.QT间期 从QRS波起点到T波终点的时程;代表心室开始兴奋去极到完全复极到静息状态的时间。
4.ST段 指从QRS波群终了到T波起点之间的与基线平齐的线段,它代表心室各部分心肌细胞均处于动作电位的平台期(2期),各部分之间没有电位差存在,曲线又恢复到基线水平。
第三节 血管生理
一、各类血管的功能特点
不论体循环或肺循环,由心室射出的血液都流经由动脉、毛细血管和静脉相互串联构成的血管系统,再返回心房。在体循环,供应各器官的血管相互间又呈并联关系(图4-17)。从生理功能上可将血管分为以下几类:
图4-17 体循环各器官血管并联关系示意图
1.左心室射血时,主动脉压升高,一方面推动动脉内的血液向前流动,另一方面使主动脉扩张,容积增大。因此,左心室射出的血液在射血期内只有一部分进入外周,另一部分则被贮存在大动脉内。主动脉瓣关闭后,被扩张的大动脉管壁发生弹性回缩,将在射血期多容纳的那部分血液继续向外周方向推动。大动脉的这种功能称为弹性贮器作用。
2.分配血管 从弹性贮器血管以后到分支为小动脉前的动脉管道,其功能是将血液输送至各器官组织,故称为分配血管。
3.毛细血管前阻力血管 小动脉和微动脉的管径小,对血流的阻力大,称为毛细血管前阻力血管。微动脉的管壁富含平滑肌,后者的舒缩活动可使血管口径发生明显变化,从而改变对血流的阻力和所在器官、组织的血流量。
4.毛细血管前括约肌 在真毛细血管的起始部常有平滑肌环绕,称为毛细血管前括肌(precapillary sphincter)。它的收缩或舒张可控制毛细血管的关闭或开放,因此可决定某一时间内毛细血管开放的数量。
5.交换血管 指真毛细血管。其管壁仅由单层内皮细胞构成,外面有一薄层基膜,故通透性很高,成为血管内血液和血管外组织液进行物质交换的场所。
6.毛细血管后阻力血管 指微静脉。微静脉因管径小,对血流也产生一定的阻力。它们的舒缩可影响毛细血管前阻力和毛细血管后阻力的比值,从而改变毛细血管压和体液在血管内和组织间隙内的分配情况。
7.容量血管 静脉和相应的动脉比较,数量较多,口径较粗,管壁较薄,故其容量较大,而且可扩张性较大,即较小的压力变化就可使容积发生较大的变化。在安静状态下,循环血量的60%-70%容纳在静脉中。静脉的口径发生较小变化时,静脉内容纳的血量就可发生很大的变化,而压力的变化较小。因此,静脉在血管系统中起着血液贮存库的作用,在生理学中将静脉称为容量血管。
8.短路血管 指一些血管床中小动脉和静脉之间的直接联系。它们可使小动脉内的血液不经过毛细血管而直接流入小静脉。在手指、足趾、耳廓等处的皮肤中有许多短路血管存在,它们在功能上与体温调节有关。
二、血流量、血流阻力和血压
血液在心血管系统中流动的一系列物理学问题属于血流动力学的范畴。血流动力学和一般的流体力学一样,其基本的研究对象是流量、阻力和压力之间的关系。由于血管是有弹性和可扩张的而不是硬质的管道系统,血液是含有血细胞和胶体物质等多种成分的液体,而不是理想液体,因此血流动力学除与一般流体力学有共同点之外,又有它自身的特点。
(一)血流量和血流速度
单位时间内流过血管某一截面的血量称为血流量,也称容积速度,其单位通常以ml/min或L/min来表示。血液中的一个质点在血管内移动的线速度,称为血流速度。血液在血管流动时,其血流速度与血流量成正比,与血管的截面成反比。
1.泊肃叶(Poiseuilli)定律泊肃叶研究了液体在管道系统内流动的规律,指出单位时间内液体的流量(Q)与管道两端的压力差P1-P2以及管道半径r的4次方成正比,与管道的长度L成反比。这些关系可用下式表示:
Q=K(r4/L)(P1-P2)
这一等式中的K为常数。后来的研究证明它与液体的粘滞度η有关。因此泊肃叶定律又可写为
Q=π(P1-P2)r4/8ηL
2.层流和湍流血液在血管内流动的方式可分为层流和湍流两类。在层流的情况下,液体每个质点的流动方向都一致,与血管的长轴平行;但各质点的流速不相同,在血管轴心处流速最快,越靠近管壁,流速越慢。因此可以设想血管内的血液由无数层同轴的圆柱面构成,在同一层的液体质点流速相同,由轴心向管壁,各层液体的流速依次递减,如图4-18所示。图中的箭头指示血流的方向,箭的长度表示流速,在血管的纵剖面上各箭头的连线形成一抛物线。泊肃叶定律适用于层流的情况。当血液的流速加快到一定程度后,会发生湍流。此时血液中各个质点的流动方向不再一致,出现旋涡。在湍流的情况下,泊肃叶定律不再适用,血流量不是与血管两端的压力差成正比,而是与压力差的平方根成正比。关于湍流的形成条件,Reynolds提出一个经验公式:
Re=VDσ/η
式中的V为血液在血管内的平均流速(单位为cm/s),D为管腔直径(单位为cm),σ为血液密度(单位为g/cm3),η为血液沾滞度(单位为泊),Re为Reynolds数,没有单位。一般当Re数超过2000时,就可发生湍流。由上式可知,在血流速度快,血管口径大,血液粘滞度低的情况下,容易产生湍流。
图4-18 层流情况下各层血液的流速
(二)血流阻力
血液在血管内流动时所遇到的阻力,称为血流阻力。血流阻力的产生,是由于血液流动时因磨擦而消耗能量,一般是表现为热能。这部分热能不可能再转换成血液的势能或动能,故血液在血管内流动时压力逐渐降低。在湍流的情况下,血液中各个质点不断变换流动的方向,故消耗的能量较层流时更多,血流阻力就较大。
血流阻力一般不能直接测量,而需通过计算得出。血液在血管中的流动与电荷在导体中流动有相似之处。根据欧姆定律,电流强度与导体两端的电位差成正比,与导体的电阻成反比。这一关系也适用于血流,即血流量与血管两端的压力差成正比,与血流阻力R成反比,可用下式表示:
Q=(P1-P2)/R
在一个血管系统中,若测得血管两端的压力差和血流量,就可根据上式计算出血流阻力。如果比较上式和泊肃叶定律的方程式,则可写出计算血流阻力的方程式,即
R=8ηL/πr4
这一算式表示,血流阻力与血管的长度和血液的粘滞度成正比,与血管半径的4次方成反比。由于血管的长度变化很小,因此血流阻力主要由血管口径和血液粘滞度决定。对于一个器官来说,如果血液粘滞度不变,则器官的血流量主要取决于该器官的阻力血管的口径。阻力血管口径增大时,血流阻力降低,血流量就增多;反之,当阻力血管口径缩小时,器官血流量就减少。机体对循环功能的调节中,就是通过控制各器官阻力血管和口径来调节各器官之间的血流分配的。
血液粘滞度是决定血流阻力的另一因素。全血的粘滞度为水的粘滞度的4-5倍。血液粘滞度的高低取决于以下几个因素:
1.红细胞比容一般说来,红细胞比容是决定血液粘滞度的最重要的因素。红细胞比容愈大,血液粘滞度就愈高。
2.血流的切率 在层流的情况下,相邻两层血液流速的差和液层厚度的比值,称为血流切率(shear rate)。从图4-18可见,切率也就是图中抛物线的斜率。匀质液体的粘滞度不随切率的变化而改变,称为牛顿液。血浆属于牛顿液。非匀质液体的粘滞度随着切率的减小而增大,称为非牛顿液。全血属非牛顿液。当血液在血管内以层流的方式流动时,红细胞有向中轴部分移动的趋势。这种现象称为轴流(axial flow)。当切率较高时,轴流现象更为明显,红细胞集中在中轴,其长轴与血管纵轴平行,红细胞移动时发生的旋转以及红细胞相互间的撞击都很小,故血液的粘滞度较低。在切率低时,红细胞可发生聚集,使血液粘滞度增高。
3.血管口径 血液在较粗的血管内流动时,血管口径对血液粘滞度不发生影响。但当血液在直径小于0.2-0.3mm的微动脉内流动时,只要切率足够高,则随着血管口径的进一步变小,血液粘滞度也变低。这一现象产生原因尚不完全清楚,但对机体有明显的益处。如果没有此种反应,血液在小血管中流动的阻力将会大大增高。
4.温度 血液的粘滞度随温度的降低而升高。人体的体表温度比深部温度低,故血液流经体表部分时粘滞度会升高。如果将手指浸在冰水中,局部血液的沾滞度可增加2倍。
(三)血压
血压是指血管内的血液对于单位面积血管壁的侧压力,也即压强。按照国际标准计量单位规定,压强的单位为帕(Pa),即牛顿/米2(N/m2)。帕的单位较小,血压数值通常用千帕(kPa)来表示(1mmHg等于0.133kPa)。
血压的形成,首先是由于心血管系统内有血液充盈。循环系统中血液充盈的程度可用循环系统平均充盈压来表示。在动物实验中,用电刺激造成心室颤动使心脏暂时停止射血,血流也就暂停,因此循环系统中各处的压力很快就取得平衡。此时在循环系统中各处所测得的压力都是相同的,这一压力数值即循环系统平均充盈压。这一数值的高低取决于血量和循环系统容量之间的相对关系。如果血量增多,或血管容量缩小,则循环系统平均充盈压就增高;反之,如果血量减少或血管容量增大,则循环系统平均充盈压就降低。用巴比妥麻醉的狗,循环系统平均充盈压约为0.93kPa(7mmHg)。人的循环系统平均充盈压估计接近这一数值。
形成血压的另一个基本因素是心脏射血。心室肌收缩时所释放的能量可分为两部分,一部分用于推动血液流动,是血液的功能;另一部分形成对血管壁的侧压,并使血管壁扩张,这部分是势能,即压强能。在心舒期,大动脉发生弹性回缩,又将一部分势能转变为推动血液的动能,使血液在血管中继续向前流动。由于心脏射血是间断性的,因此在心动周期中动脉血压发生周期性的变化。另外,由于血液从大动脉流向心房的过程中不断消耗能量,故血压逐渐降低。在机体处于安静状态时,体循环中毛细血管前阻力血管部分血压降落的幅度最大。
生物学实验中测量血压的经典方法,是将导管的一端插入动脉、静脉或心腔,将导管的另一端连至一装有水银的U形管,从U形管两边水银面高度的差即读得测定部位的血压值。水银检压计测得的压力读数为平均压。现在已有多种类型的压力换能器,可将压强能的变化转变为电能的变化,并精确地测出心动周期中各瞬间的血压数值。在临床上,常用听诊器间接测定肱动脉的收缩压和舒张压。在有些情况下,也可用导管插入血管直接测量血压。在用导管直接测量血压时,如果导管的开口正对血流,则血流的动能也转变成压强能,因此测得的血压值大于血液对血管壁的侧压。称为端压。当人体处于安静状态时,体循环中血流的动能部分在总的能量中只占很小比例,在心缩期主动脉压达最大值时,血流的动能也仅占总能量的3%。在肌肉运动时,血流速度大大加快,动能部分所占的比例增高。在肺循环中,由于肺动脉压较低,而血流速度和体循环中相近,因此血流的动能部分所占的比例较大。
三、动脉血压和动脉脉搏
(一)动脉血压
1.动脉血压的形成 前已述,循环系统同足够的血液充盈和心脏射血是形成血压的基本因素。在动脉系统,影响动脉血压的另一个因素是外周阻力。外周阻力(peripheral resistance)主要是指小动脉和微动脉对血流的阻力。假如不存在外周阻力,心室射出的血液将全部流至外周,即心室收缩释放的能量可全部表现为血流的动能,因而对血管壁的侧压不会增加。
左心室的射血是间断性的。在每个心动周期中,左心室内压随着心室的收缩和舒张发生较大幅度的变化。一般情况下,左心室每次收缩时向主动脉内射出60-80ml血液。由于小动脉和微动脉对血流有较高的阻力,以及主动脉和大动脉管壁具有较大的可扩张性,因此左心室一次收缩所射出的血液,在心缩期内大约只有三分之一流至外周,其余约三分之二被暂时贮存在主动脉和大动脉内,使主动脉和大动脉进一步扩张。主动脉压也就随之升高。这样,心室收缩时释放的能量中有一部分以势能的形式贮存在弹性贮器血管的管壁中。心室舒张时,半月瓣关闭,射血停止,被扩张的弹性贮器血管管壁发生弹性回缩,将在心缩期贮存的那部分血液继续推向外周,并使主动脉压在心舒期仍能维持在较高的水平,例如10.64kPa(80mmHg)左右,而不像心舒期的左心室内压接近0kPa。可见,由于弹性贮器血管的作用,使左心室的间断射血变为动脉内的连续血流;另一方面,还使每个心动周期中动脉血压的变动幅度远小于左心室内压的变动幅度。老年人的大动脉管壁硬化,主动脉的直径和容积增大,而可扩张性减小,弹性贮器的功能受损,因此每个心动周期中动脉血压的波动幅度明显增大。
2.动脉血压的正常值 心室收缩时,主动脉压急剧升高,在收缩期的中期达到最高值。这时的动脉血压值称为收缩压。心室舒张时,主动脉压下降,在心舒末期动脉血压的最低值称为舒张压。收缩压和舒张压的差值称为脉搏压。简称脉压。一个心动周期中每一个瞬间动脉血压的平均值,称为平均动脉压。简略计算,平均动脉压大约等于舒张压加1/3脉压。
一般所说的动脉血压是指主动脉压。因为在大动脉中血压降落很小,故通常将在上臂测得的肱动脉压代表主动脉压。我国健康青年人在安静状态时的收缩压为13.3-16.0kPa(100-120mmHg),舒张压为8.0-10.6kPa(60-80mmHg),脉搏压为4.0-5.3kPa(30-40mmHg),平均动脉压在13.3kPa(100mmHg)左右。
动脉血压 除存在个体差异外,还有性别和年龄的差异。一般说来,女性在更年期前动脉血压比同龄男性的低,更年期后动脉血压升高。男性和女性的动脉血压都随年龄的增长而逐渐升高,收缩压的升高比舒张压的升高更为显著。新生儿的收缩压仅为5.3kPa(40mmHg)左右。出生后第一个月内,收缩压很快升高,到第一月末约可达到10.6kPa(80mmHg)。以后,收缩压继续升高,到12岁时约为14.0kPa(105mmHg)。在青春期,收缩压又较快地上升,17岁的男性青年,收缩压可达16.0kPa(120mmHg)。青春期以后,收缩压随年龄增长而缓慢升高。至60岁时,收缩压约18.6kPa(140mmHg)。
当血液从主动脉流向外周时,因不断克服血管对血流的阻力而消耗能量,血压也就逐渐降低。在各段血管中,血压降落的幅度与该段血管对血流的阻力的大小成正比。在主动脉和大动脉段,血压降落较小。如果主动脉的平均压为13.3kPa(100mmHg)。则到直径为3mm的动脉处,平均压仍在12.6kPa(95mmHg)左右。到小动脉时,血流阻力大,血压降落的幅度也变大。在体循环中,微动脉段的血流阻力最大,血压降落也最为显著。如果微动脉起始端的血压为11.3kPa(85mmHg),则血液流经微动脉后压力降落 7.3kPa(55mmHg),故在毛细血管起始端,血压仅4.0kPa(30mmHg)。在不同的动脉段记录血压时,可以看到从主动脉到外周动脉,血压的波动幅度变大。和主动脉内的血压波动相比,外周动脉的收缩压较高,舒张压较低,故脉搏压较大,而平均压低于动脉压(图4-19)。产生这种现象的原因,主要是由于血压压力波的折返。当动脉的压力波动在传播至较小的动脉分支处,特别是到微动脉时,因受到阻碍而发生折返。折返的压力波逆流而上,如果遇到下行的波动,两者可发生叠加,形成一个较大的波。在股动脉记录血压时,常可看到在一个大的波后面有一个较小的返折波(图4-19),故股动脉的血压波动幅度大于主动脉的血压波动幅度。
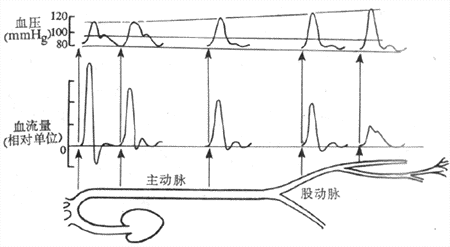
图4-19 主动脉和外周动脉的脉搏压、平均压和血流变化(1mmHg=0.133kPa)
3.影响动脉血压的因素,凡是能影响心输出量和外周阻力的各种因素,都能影响动脉血压。循环血量和血管系统容量之间的相互关系,即循环系统内血液充盈的程度,也能影响动脉血压。现将影响动脉血压因素分述如下:
(1)心脏每搏输出量:如果每搏输出量增大,心缩期射入主动脉的血量增多,心缩期中主动脉和大动脉内增加的血量变多,管壁所受的张力也更大,故收缩期动脉血压的升高更加明显。由于动脉血压升高,血流速度外周阻力和心率的变化不大,则大动脉内增多的血量仍可在心舒期流至外周,到舒张期末,大动脉内存留的血量和每搏输出量增加之前相比,增加并不多。因此,当每搏输出量增加而外周阻力和心率变化不大时,动脉血压的升高主要表现为收缩压的升高,舒张压可能升高不多,故脉压增大。反之,当每搏输出量减少时,则主要使收缩压降低,脉压减小。可见,在一般情况下,收缩压的高低主要反映心脏每搏输出量的多少。
(2)心率:如果心率加快,而每搏输出量和外周阻力都不变,由由于心舒期缩短,在心舒期内流至外周的血液就减少,故心舒期末主动脉内存留的血量增多,舒张期血压就升高。由于动脉血压升高可使血流速度加快,因此在心缩期内可有较多的血液流至外周,收缩压的升高不如舒张压的升高显著,脉压比心率增加前减小。相反,心率减慢时,舒张压降低的幅度比收缩压降低的幅度大,故脉压增大。
(3)外周阻力:如果心输出量不变而外周阻力加大,则心舒期中血液向外周流动的速度减慢,心舒期末存留在主动脉中的血量增多,故舒张压升高。在心缩期,由于动脉血压升高使血流速度加快,因此收缩压的升高不如舒张压的升高明显,故脉压加大。可见,在一般情况下,舒张压的高低主要反映外周阻力的大小。
外周阻力的改变,主要是由于骨骼肌和腹腔器官阻力血管口径的改变。原发性高血压的发病,主要是由于阻力血管口径变小而造成外周阻力过高。另外,血液粘滞度也影响外周阻力。如果血液粘滞度增高,外周阻力就增大,舒张压就升高。
(4)主动脉和大动脉的弹性贮器作用:如前所述,由于主动脉和大动脉的弹性贮器作用,动脉血压的波动幅度明显小于心室内压的波动幅度。老年人的动脉管壁硬化,大动脉的弹性贮器作用减弱,故脉压增大。
(5)循环血量和血管系统容量的比例:循环血量和血管系统容量相适应,才能使血管系统足够地充盈,产生一定的体循环平均充盈压。在正常情况下,循环血量和血管容量是相适应的,血管系统充盈程度的变化不大。失血后,循环血量减少。此时如果血管系统的容量改变不大,则体循环平均充盈压必然降低,使动脉血压降低。在另一些情况下,如果循环血量不变而血管系统容量增大时,也会造成动脉血压下降。
上述对影响动脉血压的各种因素,都是在假设其它因素不变的前提下,分析某一因素发生变化时对动脉血压可能发生的影响。实际上,在各种不同的生理情况下,上述各种影响动脉血压的因素可同时发生改变。因此,在某种生理情况下动脉血压的变化,往往是各种因素相互作用的综合结果。
(二)动脉脉搏
在每个心动周期中,动脉内的压力发生周期性的波动。这种周期性的压力变化可引起动脉血管发生搏动,称为动脉脉搏。在手术时暴露动脉,可以直接看到动脉随每次心搏而发生的搏动。用手指也可摸到身体浅表部位的动脉搏动。
1.动脉脉搏的波形 用脉搏描记仪可以记录浅表动脉搏的波形。这种记录图形称为脉搏图(图4-20)。动脉脉搏的波形可因描记方法和部位的不同而有差别,但一般都包括以下几个组成部分:
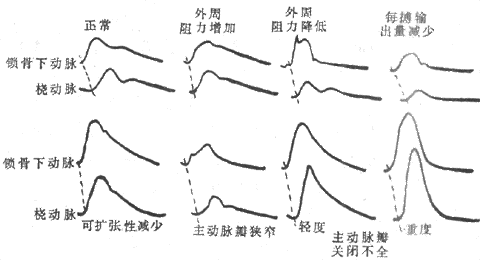
图4-20不同情况下锁骨下动脉与桡动脉的脉搏图
(1)上升支:在心室快速射血期,动脉血压迅速上升,管壁被扩张,形成脉搏波形中的上升支。上升支的斜率和幅度受射血速度、心输出量以及射血所遇的阻力的影响,射血遇到的阻力大,心输出量小,射血速度慢,则脉搏波形中上升支的斜率小,幅度也低;反之,射血所遇的阻力小,心输出量大,射血速度快,则上升支较陡,幅度也较大。大动脉的可扩张性减小时,弹性贮器作用减弱,动脉血压的波动幅度增大,脉搏波上升支的斜率和幅度也加大。主动脉瓣狭窄时,射血阻力高,脉搏波上升支的斜率和幅度都较小。
(2)下降支:心室射血的后期,射血速度减慢,进入主动脉的血量少于由主动脉流向外周的血量,故被扩张的大动脉开始回缩,动脉血压逐渐降低,形成脉搏波形中下降支的前段。随后,心室舒张,动脉血压继续下降,形成下降支的其余部分。在主动脉记录脉搏图时,其下降支上有一个切迹,称为降中峡。降中峡发生在主动脉瓣关闭的瞬间。因为心室舒张时室内压下降,主动脉内的血液向心室方向返流。这一返流使主动脉瓣很快关闭。返流的血液使主动脉根部的容积增大,并且受到闭合的主动脉瓣阻挡,发生一个返折波,因此在降中峡的后面形成一个短暂的向上的小波,称为降中波。动脉脉搏波形中下降支的形状可大致反映外周阻力的高低。外周阻力高时,脉搏波降支的下降速率较慢,切迹的位置较高。如果外周阻力较低,则下降支的下降速率较快,切迹位置较低,切迹以后下降支的坡度小,较为平坦。主动脉瓣关闭不全时,心舒期有部分血液倒流入心室。故下降支很陡,降中波不明显或者消失。
2.动脉脉搏波的传播速度 动脉脉搏可以沿着动脉管壁向外周血管传播,其传播的速度远较血流的速度为快。一般说来,动脉管壁的可扩张性愈大,脉搏波的传播速度就愈慢。由于主动脉的可扩张性最大,故脉搏波在主动脉的传播速度最慢,约3-5m/s,在大动脉的传播速度约为7-10m/s,到小动脉段可加快到15-35m/s。老年人主动脉管壁的可扩张性减小,脉搏波的传播速度可增加到大约10m/s。
由于小动脉和微动脉对血流的阻力很大,故在微动脉段以后脉搏波动即大大减弱。到毛细血管,脉搏已基本消失。
很早以来,医生在进行诊断时要按摸病人的脉搏,最通常的是按病人桡动脉的脉搏。按脉可以了解病人的脉搏频率和节律是否规则等情况,同时也在心理上构成了医生和病人之间的接触和联系。中医把切脉作为诊断疾病的重要手段之一。由于动脉脉搏与心输出量、动脉的可扩张性以及外周阻力等因素有密切的关系。因此,在某些情况下脉搏可以反映心血管系统的异常状况。中医学中的脉象,就是研究各种生理和病理情况下桡动脉脉搏的特征。在中医诊断学中,对脉象有很详细的描述。
四、静脉血压和静脉回心血量
静脉在功能上不仅仅是作为血液回流入心脏的通道,由于整个静脉系统的容量很大,而且静脉容易被扩张,又能够收缩,因此静脉起着血液贮存库的作用。静脉的收缩或舒张可有效地调节回心血量和心输出量,使循环机能能够适应机体在各种生理状态时的需要。
(一)静脉血压
当体循环血液经过动脉和毛细血管到达微静脉时,血压下降至约2.0-2.7kPa(15-20mmHg)。右心房作为体循环的终点,血压最低,接近于零。通常将右心房和胸腔内大静脉的血压称为中心静脉压,而各器官静脉的血压称为外周静脉压。中心静脉压的高低取决于心脏射血能力和静脉回心血量之间的相互关系。如果心脏射血能力较强,能及时地将回流入心脏的血液射入动脉,中心静脉压就较低。反之,心脏射血能力减弱时,中心静脉压就升高。另一方面,如果静脉回流速度加快,中心静脉压也会升高。因此,在血量增加,全身静脉收缩,或因微动脉舒张而使外周静脉压升高等情况下,中心静脉压都可能升高。可见,中心静脉压是反映心血管功能的又一指标。临床上在用输液治疗休克时,除须观察动脉血压变化外,也要观察中心静脉压的变化。中心静脉压的正常变动范围为0.4-1.2kPa(4-12mmH2O)。如果中心静脉压偏低或有下降趋势,常提示输液量不足;如果中心静脉压高于正常并有进行性升高的趋势,则提示输液过快或心脏射血功能不全。当心脏射血功能减弱而使中心静脉压升高时,静脉回流将会减慢,较多的血液滞留在外周静脉内,故外周静脉压升高。
静脉脉搏动脉脉搏波在到达毛细血管时已经消失,因此外周静脉没有脉搏波动。但是右心房在心动周期中的血压波动可以逆向传递到与心房相连续的大静脉,引起这些大静脉的周期性压力和容积变化,形成静脉脉搏。由于引起搏动的原因不同,故大静脉的脉搏波形和动脉脉搏的波形完全不同。正常情况下,静脉脉搏不很明显。但在心力衰竭时,静脉压升高,右心房内的压力波动也较容易传递至大静脉,故在心力衰竭病人的颈部常可见到较明显的静脉搏动。
(二)重力对静脉压的影响
血管系统内的血液因受地球重力场的影响,产生一定的静水压。因此,各部分血管的血压除由于心脏作功形成以外,还要加上该部分血管处的静水压。各部分血管的静水压的高低取决于人体所取的体位。在平卧时,身体各部分血管的位置大致都处在和心脏相同的水平,故静水压也大致相同。但当人体从平卧转为直立时,足部血管内的血压比卧位时高。其增高的部分相当于从足至心脏这样的一段血柱高度形成的静水压,约12kPa(90mmHg),见图4-21。而在心脏水平以上的部分,血管内的压力较平卧时为低,例如颅顶脑膜矢状窦内压可降至-1.33kPa(-10mmHg)。重力形成的静水压的高低,对于处在同一水平上的动脉和静脉是相同的,但是它对静脉功能的影响远比对动脉功能的影响大。因为静脉较动脉有一明显的特点,即其充盈程度受跨壁压的影响较大。跨壁压是指血管内血液对管壁的压力和血管外组织对管壁的压力之差。一定的跨壁压是保持血管充盈膨胀的必要条件。跨壁压减小到一定程度,血管就不能保持膨胀状态,即发生塌陷。静脉管壁较薄,管壁中弹性纤维和平滑肌都较少,因此当跨壁压降低时就容易发生塌陷。此时静脉的容积也减小。当跨壁压增大时,静脉就充盈,容积增大。当人在直立时,足部的静脉充盈饱满,而颈部的静脉则塌陷。静脉的这一特性在人类特别值得注意。因为当人在直立时,身体中大多数容量血管都处于心脏水平以下,如果站立不动,由于身体低垂部分的静脉充盈扩张,可比在卧位时多容纳400-600ml血液,这部分血液主要来自胸腔内的血管。这样就造成体内各部分器官之间血量的重新分配,并导致暂时的回心血量减少,中心静脉压降低,每搏输出量减少和收缩压降低。后文将述及,这些变化会发动神经和体液的调节机制,使骨骼肌、皮肤和肾、腹腔内脏的阻力血管收缩以及心率加快,故动脉血压可以恢复。许多动物由于四足站地,多数容量血管都处于心脏水平以上,故体位改变时血量分配的变化不像在人类中那样明显。
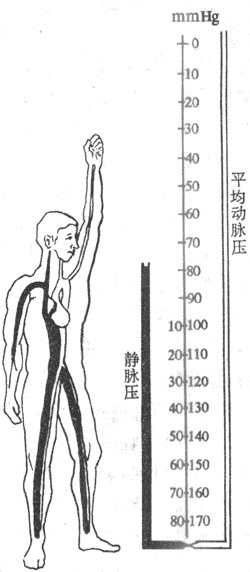
图4-21 直立体位对肢体动脉和静脉血压的影响(1mmHg=0.13kPa)
(三)静脉血流
1.静脉对血流的阻力 单位时间内由静脉回流入心脏的血量等于心输出量。在静脉系统中,由微静脉至右心房的压力降落仅约2kPa(15mmHg)。可见静脉对血流的阻力很小,约占整个体循环总阻力的15%。静脉在血液循环中是将血液从组织引流回心脏的通道,并且起血液贮存库的作用。小的血流阻力与静脉的功能是相适应的。
微静脉在功能上是毛细血管后阻力血管。毛细血管后阻力的改变可影响毛细血管血压。因为后者的高低取决于毛细血管前阻力和毛细血管后阻力的比值。微静脉收缩,使毛细血管后阻力升高,如果毛细血管前阻力不变,则毛细血管前阻力和毛细血管后阻力的比值变小,于是毛细血管血压升高,组织液的生成增多(见后)。因此,机体可通过对微静脉收缩状态的调节来控制血液和组织液之间的液体交换,并间接地调节循环血量。
前面提到,静脉的跨壁压改变时可改变静脉的扩张状态,从而也改变静脉对血流的阻力。大静脉在处于扩张状态时,对血流的阻力很小;但当管壁塌陷时,因其管腔截面由圆形变成椭圆形,截面积减小,因此对血流的阻力增大。另外,血管周围组织对静脉的压迫也可增加静脉对血流的阻力。例如锁骨下静脉在跨越第一肋骨处受肋骨的压迫;颈部皮下的颈外静脉直接受外界大气的压迫;腹腔内的大静脉受腹腔器官的压迫,等等。位于胸腔内的大静脉则因受胸膜腔内负压的作用,跨壁压较大,一般不会塌陷。颅腔、脊柱、骨和肝、脾等器官内的静脉,因受到血管周围结缔组织的支持,也不会塌陷。
2.静脉回心血量及其影响因素单位时间内的静脉回心血量取决于外周静脉压和中心静脉压的差,以及静脉对血流的阻力。故凡能影响外周静脉压、中心静脉压以及静脉阻力的因素,都能影响静脉回心血量。
(1)体循环平均充盈压:体循环平均充盈压是反映血管系统充盈程度的指标。实验证明,血管系统内血液充盈程度愈高,静脉回心血量也就愈多。当血量增加或容量血管收缩时,体循环平均充盈压升高,静脉回心血量也就增多。反之,血量减少或容量血管舒张时,体循环平均充盈压降低,静脉回心血量减少。
(2)心脏收缩力量:心脏收缩时将血液射入动脉,舒张时则可以从静脉抽吸血液。如果心脏收缩力量强,射血时心室排空较完全,在心舒期心室内压就较低,对心房和大静脉内血液的抽吸力量也就较大。右心衰竭时,射血力量显著减弱,心舒期右心室内压较高,血液淤积在右心房和大静脉内,回心血量大大减少。患者可出现颈外静脉怒张,肝充血肿大,下肢浮肿等特征。左心衰竭时,左心房压和肺静脉压升高,造成肺淤血和肺水肿。
(3)体位改变:前已述,当人体从卧位转变为立位时,身体低垂部分静脉扩张,容量增大,故回心血量减少。站立时下肢静脉容纳血量增加的程度可受到若干因素的限制,例如下肢静脉内的静脉瓣,以及下面将叙述的下肢肌肉收缩运动和呼吸运动等。下肢静脉瓣膜受损的人,常不能长久站立。即使在正常人,如长久站立不动,也会导致回心血量减少,动脉血压降低。体位改变对静脉回心血量的影响,在高温环境中更加明显。在高温环境中,皮肤血管舒张,皮肤血管中容纳的血量增多。因此,如果人在高温环境中长时间站立不动,回心血量就会明显减少,导致心输出量减少和脑供血不足,可引起头晕甚至昏厥。长期卧床的病人,静脉管壁的紧张性较低,可扩张性较高,加之腹腔和下肢肌肉的收缩力量减弱,对静脉的挤压作用减小,故由平卧位突然站起来时,可因大量血液积滞在下肢,回心血量过小而发生昏厥。
(4)骨骼肌的挤压作用:人体在站立位的情况下,如果下肢进行肌肉运动,回心血量和在没有肌肉运动时就不一样。一方面,肌肉收缩时可对肌肉内和肌肉间的静脉发生挤压,使静脉血流加快;另一方面,因静脉内有瓣膜存在,使静脉内的血液只能向心脏方向流动而不能倒流。这样,骨骼肌和静脉瓣膜一起,对静脉回流起着“泵”的作用,作为“静脉泵”或“肌肉泵”。下肢肌肉进行节律性舒缩活动时,例如步行,肌肉泵的作用就能很好地发挥。因为当肌肉收缩时,可将静脉内的血液挤向心脏,当肌肉舒张时,静脉内压力降低,有利于微静脉和毛细血管内的血液流入静脉,使静脉充盈。肌肉泵的这种作用,对于在立位情况下降低下肢静脉压和减少血液在下肢静脉内潴留有十分重要的生理意义。例如,在站立不动时,足部的静脉压为12kPa(90mmHg),而在步行时则降低至3.3kPa(25mmHg)以下。在跑步时,两下肢肌肉泵每分钟挤出的血液可达数升。在这种情况下,下肢肌肉泵的作功在相当程度上加速了全身的血液循环,对心脏的泵血起辅助的作用。但是,如果肌肉不是作节律性的舒缩,而是维持在紧张性收缩状态,则静脉持续受压,静脉回流反而减少。
(5)呼吸运动:呼吸运动也能影响静脉回流。在第五章会详述,胸膜腔内压是低于大气压的,称为胸膜腔负压。由于胸膜腔内压为负压,胸腔内大静脉的跨壁压较大,故经常处于充盈扩张状态。在吸气时,胸腔容积加大,胸膜腔负压值进一步增大,使胸腔内的大静脉和右心房更加扩张,压力也进一步降低,因此有利于外周静脉内的血液回流入右心房。由于回心血量增加,心输出量也相应增加。呼气时,胸膜腔负压值减小,由静脉回流入右心房的血量也相应减少。可见,呼吸运动对静脉回流也起着“泵”的作用。有些人在站立时呼吸加深,显然可以促进身体低垂部分的静脉血液回流。需要指出,呼吸运动对肺循环静脉回流的影响和对体循环的影响不同。吸气时,随着肺的扩张,肺部的血管容积显著增大,能贮留较多的血液,故由肺静脉回流至左心房的血量减少,左心室的输出量也相应减少。呼气时的情况则相反。
五、微循环
微循环是指微动脉和微静脉之间的血液循环。血液循环最根本的功能是进行血液和组织之间的物质交换,这一功能就是在微循环部分实现的。
(一)微循环的组成
各器官、组织的结构和功能不同,微循环的结构也不同。人手指甲皱皮肤的微循环形态比较简单,微动脉和微静脉之间仅由呈袢状的毛细血管相连。骨骼肌和肠系膜的微循环形态则比较复杂。典型的微循环由微动脉、后微动脉、毛细血管前括约肌、真毛细血管、通血毛细血管(或称直捷通路)、动-静脉吻合支和微静脉等部分组成。图4-22是一个典型的微循环单元。
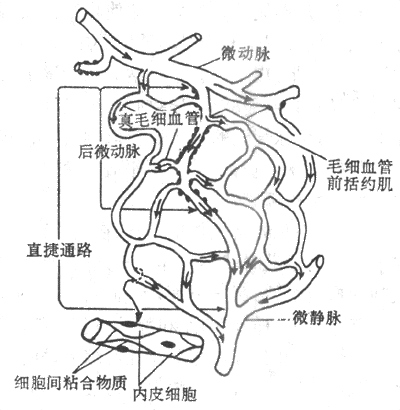
图4-22 肠系膜微循环模式图
微动脉管壁有环行的平滑肌,其收缩和舒张可控制微血管的血流量。微动脉分支成为管径更细的后微动脉。每根后微动脉向一根至数根真毛细血管供血。真毛细血管通常从后微动脉以直角方向分出。在真毛细血管起始后端通常有1-2个平滑肌细胞,形成一个环,即毛细血管前括约肌。该括约肌的收缩状态决定进入真毛细血管的血流量。
毛细血管的血液经微静脉进入静脉。最细的微静脉管径不超过20-30μm,管壁没有平滑肌,在功能上有交换血管的作用。较大的微静脉管壁有平滑肌,在功能上是毛细血管后阻力血管。微静脉的舒缩状态可影响毛细血管血压,从而影响毛细血管处的液体交换和静脉回心血量。
另外,微动脉和微静脉之间还可通过直捷通路和动-静脉短路发生沟通。直捷通路(thoroughfare channel)是指血液从微动脉经后微动脉和通血毛细血管进入微静脉的通路。通血毛细血管是后微动脉的直接延伸,其管壁平滑肌逐渐稀小以至消失。直捷通路经常处于开放状态,血流速度较快,其主要功能并不是物质交换,而是使一部分血液能迅速通过微循环而进入静脉。直捷通路在骨骼肌组织的微循环中较为多见。动-静脉短路(arteriovenous shunt)是吻合微动脉和微静脉的通道,其管壁结构类似微动脉。在人体某些部分的皮肤和皮下组织,特别是手指、足趾、耳廓等处,这类通路较多。动-静脉吻合支在功能上不是进行物质交换,而是在体温调节中发挥作用的。当环境温度升高时,动-静脉吻合支开放增多,皮肤血流量增加,皮肤温度升高,有利于发散身体热量。环境温度低时,则动-静脉短路关闭,皮肤血流量减少,有利于保存体热。动-静脉短路开放,会相对地减少组织对血液中氧的摄取。在某些病理状态下,例如感染性和中毒性休克时,动-静脉短路大量开放,可加重组织的缺氧状况。
(二)毛细血管的结构和通透性
毛细血管壁由单层内皮细胞构成,外面有基膜包围,总的厚度约0.5μm,在细胞核的部分稍厚。内皮细胞之间相互连接处存在着细微的裂隙,成为沟通毛细血管内外的孔道(图4-23)。
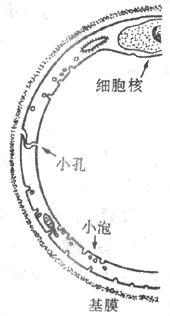
图4-23 毛细血管壁亚显微结构示意图
毛细血管内皮有四种主要类型:
1.连续内皮 分布在皮肤、骨骼肌、平滑肌、心肌、肺等多数器官组织。内皮细胞厚度为0.1-0.2μm,细胞核处稍厚。细胞之间有紧密连接(tight junction),其裂隙大小一般小于血浆蛋白质分子的大小,故水、离子,小于血浆蛋白的溶质分子都可以通过。这种内皮对血浆中各种溶质的通透性很小。脂浴性物质如O2和CO2以及水分子可以直接通过内皮细胞的细胞膜和胞浆。另外,内皮细胞还有吞饮功能,在细胞内可看到吞饮囊泡,囊泡内容是血浆或组织液,包含有蛋白质分子。有时数个囊泡可融合成一个贯通内皮细胞壁的暂时的通道。
2.有孔内皮分布在胃肠粘膜、腺体、肾小球和肾小管周围毛细血管。这类内皮在5%-50%的面积上细胞厚度不到0.05μm,并且有小孔。其余部分的结构与连续内皮相似。在肾小球毛细血管,管壁的小孔是直径为50-60nm的圆孔,小孔是开放的,外面被基膜覆盖。在胃肠粘膜和肾小管周围毛细血管,小孔被一层纤薄的隔膜封闭。有孔内皮对水和小的溶质的通透性高于连续内皮,但对血浆蛋白质的通透性仍很小。
3.非连续内皮 分布在肝、骨髓、脾的血窦。内皮细胞的间隙可宽达1μm,并且基膜也是不连续的。蛋白质和其它大分子可以自由通过这些间隙。
4.紧密连接内皮 分布在中枢神经系统和视网膜。这类内皮细胞较高大,故毛细血管管壁较厚。内皮细胞之间都是紧密连接。内皮细胞内很少见到吞饮囊泡。水和脂溶性分子可直接通过细胞,一些离子和小分子非脂溶性物质(如葡萄糖、氨基酸)则只能由特异的载体转运。
(三)毛细血管的数量和交换面积
有人粗略估计,人体全身约有400亿根毛细血管。不同器官组织中毛细血管的密度有很大差异,例如在心肌、脑、肝、肾,毛细血管的密度为每立方毫米组织2500-3000根;骨骼肌为每立方毫米组织100-400根;骨、脂肪、结缔组织中毛细血管密度较低。假设毛细血管的平均半径为3μm,平均长度为750μm,则每根毛细血管的表面积约为14000μm2。由于微静脉的起始段也有交换功能,故估计每根毛细血管的有效交换面积为22000μm2。由此可以估计全身毛细血管(包括有交换功能的微静脉)总的有效交换面积将近1000m2。
(四)策循环的血流动力学
微循环中的血流一般为层流。血液在流经微循环血管网时血压逐渐降低。在直径为8-40μm的微动脉处,对血流的阻力最大,血压降落也最大。到毛细血管的靠动脉端,血压约4.0-5.3kPa(30-40mmHg),毛细血管中段血压约3.3kPa(25mmHg),至靠静脉端约1.3-2.0kPa(10-15mmHg)。毛细血管血压的高低取决于毛细血管和前阻力和毛细血管后阻力的比值。一般说来,当这一比例为5:1时,毛细血管的平均血压约为2.7kPa(20mmHg)。这一比值增大时,毛细血管血压就降低;比值变小时毛细血管血压升高。某一组织中微循环的血流量与微动脉和微静脉之间的血压差成正比,与微循环中总的血流阻力成反比。由于在总的血流阻力中微动脉处的阻力占较大比例,故微动脉的阻力对血流量的控制起主要作用。
测量一个器官的血流量时,常可见到在一定时间内其血流量是稳定的。但如果在显微镜下观察微循环中单个血细胞的移动速度,则可看到在同一时间内不同微血管中的流速是有很大差别的,而且同一血管在不同时间内流速也有较大变化。其原因是由于后微动脉和毛细血管前括约肌不为发生每分钟约5-10次的交替性收缩和舒张,称为血管舒活动。后微动脉和毛细血管前括约肌收缩,其后的真毛细血管网关闭,舒张时真毛细血管网开放。在安静状态下,骨骼肌组织中在同一时间内只有20%-35%的真毛细血管处于开放状态。血管舒缩活动主要与局部组织的代谢有关。毛细血管关闭时,该毛细血管周围组织中代谢产物积聚,氧分压降低。代谢产物和低氧都能导致局部的后微动脉和毛细血管前括约肌舒张及毛细血管开放,于是局部组织内积聚的代谢产物被血流清除,后微动脉和毛细血管前括约肌又收缩,使毛细血管关闭。如此周而复始。当组织代谢活动加强时,愈来愈多的微动脉和毛细血管前括约肌发生舒张,使愈来愈多的毛细血管处于开放状态,从而使血液和组织、细胞之间发生交换的面积增大,交换的距离缩短。因此,微循环的血流量和组织的代谢活动水平相适应。
(五)血液和组织液之间的物质交换
组织、细胞之间的空间称为组织间隙,其中为组织液所充满。组织液是组织、细胞直接所处的环境。组织、细胞通过细胞膜和组织液发生物质交换。组织液与血液之间则通过毛细血管壁进行物质交换。因此,组织、细胞和血液之间的物质交换需通过组织液作为中介。
血液和组织液之间的物质交换主要是通过以下几种方式进行的:
1.扩散扩散是指液体中溶质分子的热运动,是血液和组织液之间进行物质交换的最主要的方式。毛细血管内外液体中的分子,只要其直径小于毛细血管壁的孔隙,就能通过管壁进行扩散运动。分子运动是可以向各个不同方向进行的杂乱的运动,故当血液流经毛细血管时,血液内的溶质分子可以扩散入组织液,组织液内的溶质分子也可以扩散入血液。对于某一种物质来说,其通过毛细血管壁进行的扩散的驱动力是该物质在管壁两侧的浓度差,即从浓度高的一侧向浓度低的一侧发生净移动。溶质分子在单位时间内通过毛细血管壁进行扩散的速率与该溶质分子在血浆和组织液中的浓度差、毛细血管壁对该溶质分子的通透性、毛细血管壁的有效交换面积等因素成正比,与毛细血管壁的厚度(即扩散距离)成反比。对于非脂溶性物质,毛细血管壁的通透性(紧密连接内皮除外)与溶质分子的大小有关,分子愈小,通透性愈大。毛细血管壁孔隙的总面积虽仅占毛细血管壁总面积的约千分之一,但由于分子运动的速度高于毛细血管血流速度数十倍,故血液在流经毛细血管时,血浆和组织液的溶质分子仍有足够的时间进行扩散交换。脂溶性物质如O2、CO2等可直接通过内皮细胞进行扩散,因此整个毛细血管壁都成为扩散面,单位时间内扩散的速率更高。
2.滤过和重吸收当毛细血管壁两侧的静水压不等时,水分子就会通过毛细血管壁从压力高的一侧向压力低的一侧移动。水中的溶质分子,如其分子直径小于毛细血管壁的孔隙,也能随同水分子一起滤过。另外,当毛细血管壁两侧的渗透压不等时,可以导致水分子从渗透压低的一侧向渗透压高的一侧移动。由于血浆蛋白质等胶体物质较难通过毛细血管壁的孔隙,因此血浆的胶体渗透压能限制血浆的水分子向毛细血管外移动;同样,组织液的胶体渗透压则限制组织液的水分子向毛细血管内移动。在生理学中,将由于管壁两侧静水压和胶体渗透压的差异引起的液体由毛细血管内向毛细血管外的移动称为滤过,而将液体向相反方向的移动称为重吸收。血液和组织液之间通过滤过和重吸收的方式发生的物质交换,和通过扩散方式发生的物质交换相比,仅占很小的一部分,但在组织液的生成中起重要的作用。
3.吞饮在毛细血管内皮细胞一侧的液体可被内皮细胞膜包围交吞饮入细胞内,形成吞饮囊泡。囊泡被运送至细胞的另一侧,并被排出至细胞外。因此,这也是血液和组织液之间通过毛细血管壁进行物质交换的一种方式。一般认为,较大的分子如血浆蛋白等可以由这种方式通过毛细血管壁进行交换。
六、组织液的生成
正常成人的体重的60%左右是水,其中约5/8存在于细胞内,称为细胞内液;其余3/8存在于细胞外,称为细胞外液。细胞外液中,约有1/5在血管内,即血浆的水分;其余4/5在血管外,即组织液和各种腔室内液体(脑脊液、眼球内液等)的水分。组织液存在于组织、细胞的间隙内,绝大部分呈胶冻状,不能自由流动,因此不会因重力作用而流至身体的低垂部分;将注射针头插入组织间隙内,也不能抽出组织液。组织液凝胶的基质是胶原纤维和透明质酸细丝。组织液中有极小一部分呈液态,可自由流动。组织液中各种离子成分与血浆相同。组织液中也存在各种血浆蛋白质,但其浓度明显低于血浆。
(一)组织液的生成
组织液是血浆滤过毛细血管壁而形成的。如前所述,液体通过毛细血管壁的滤过和重吸收取决于四个因素,即毛细血管血压(Pc)、组织液静水压(Pif),血浆胶体渗透压(πp)和组织液胶体渗透压(πif)。其中,Pc和πif是促使液体由毛细血管内向血管外滤过的力量,而πp和Pif是将液体从血管外重吸收入毛细血管内的力量。滤过的力量(即Pc+πif)和重吸收的力量(即πp+Pif)之差,称为有效滤过压。单位时间内通过毛细血管壁滤过的液体量V等于有效滤过压与滤过系数Kf的乘积,即
V=Kf[(Pc+πif)-(πp+Pif)]
滤过系数的大小取决于毛细血管壁对液体的通透性和滤过面积。以图4-24所设的各种压力数值为例,可见在毛细血管动脉端的有效滤过压为1.3kPa(10mmHg),液体滤出毛细血管;而在毛细血管静脉端的有效滤过压力为负值,故发生重吸收。总的说来,流经毛细血管的血浆,约有0.5%在毛细血管动脉端以滤过的方式进行组织间隙,其中约90%在静脉端被重吸收回血液,其余约10%进行毛细淋巴管,成为淋巴液。
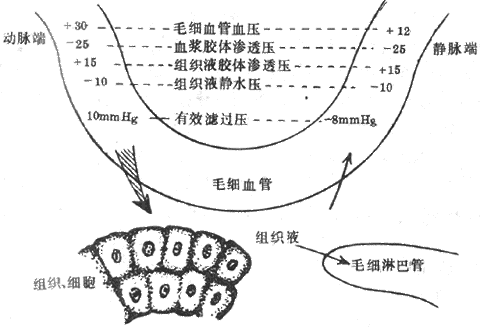
图4-24 组织液生成与回流示意图
+代表使液体滤出毛细血管的力量 -代表使液体吸收回毛细血管的力量
(1mmHg=0.133kPa)
(二)影响组织液生成的因素
在正常情况下,组织液不断生成,又不断被重吸收,保持动态平衡,故血量和组织液量能维持相对稳定。如果这种动态平衡遭到破坏,发生组织液生成过多或重吸收减少,组织间隙中就有过多的潴留,形成组织水肿。上述决定有效滤过压的各种因素,如毛细血管血压升高和血浆胶体渗透压降低时,都会使组织液生成增多,甚至引起水肿。静脉回流受阻时,毛细血管血压升高,组织液生成也会增加。淋巴回流受阻时,组织间隙内组织液积聚,可导致组织水肿。此外,在某些病理情况下,毛细血管壁的通透性增高,一部分血浆蛋白质滤过进行组织液,使组织液生成增多,发生水肿。
七、淋巴液的生成和回流
淋巴管系统是组织液向血液回流的一个重要的辅助系统。毛细淋巴管以稍膨大的盲端起始于组织间隙,彼此吻合成网,并逐渐汇合成大的淋巴管。全身的淋巴液经淋巴管收集,最后由右淋巴导管和胸导管导入静脉。
(一)淋巴液的生成
组织液进入淋巴管,即成为淋巴液。因此,来自某一组织的淋巴液的成分和该组织的组织液非常接近。在毛细淋巴管起始端,内皮细胞的边缘像瓦片般互相覆盖,形成向管腔内开启的单向活瓣。另外,当组织液积聚在组织间隙内时,组织中的胶原纤维和毛细淋巴管之间的胶原细丝可以将互相重叠的内皮细胞边缘拉开,使内皮细胞之间出现较大的缝隙。因此,组织液包括其中的血浆蛋白质分子可以自由地进入毛细淋巴管。
正常成人在安静状态下大约每小时有120ml淋巴液流入血液循环,其中约100ml经由胸导管,20ml经由右淋巴导管进入血液。以此推算,每天生成的淋巴液总量约为2-4L,大致相当于全身血浆总量。组织液和毛细淋巴管内淋巴液的压力差是组织液进入淋巴管的动力。组织液压力升高时,能加快淋巴液的生成速度。
(二)淋巴液的回流及影响淋巴液回流的因素
毛细淋巴管汇合形成集合淋巴管。后者的管壁中有平滑肌,可以收缩。另外,淋巴管中有瓣膜,使淋巴液不能倒流。淋巴管壁平滑肌的收缩活动和瓣膜共同构成“淋巴管泵”,能推动淋巴流动。淋巴管周围组织对淋巴管的压迫也能推动淋巴流动,例如肌肉收缩,相邻动脉的搏动,以及外部物体对身体组织的压迫和按摩等等。凡能增加淋巴生成的因素也都能增加淋巴液的回流量。
淋巴液回流的生理功能,主要是将组织液中的蛋白质分子带回至血液中,并且能清除组织液中不能被毛细血管重吸收的较大的分子以及组织中的红细胞和细菌等。小肠绒毛的毛细淋巴管对营养物质特别是脂肪的吸收起重要的作用。由肠道吸收的脂肪的80%-90%是经过这一途径被输送入血液的。因此小肠的淋巴呈乳糜状。淋巴回流的速度虽较缓慢,但一天中回流的淋巴液相当于全身血浆总量,故淋巴液回流在组织液生成和重吸收的平衡中起着一定的作用。
第四节 心血管活动的调节
人体在不同的生理状况下,各器官组织的代谢水平不同,对血流量的需要也不同。机体的神经和体液机制可对心脏和各部分血管的活动进行调节,从而适应各器官组织在不同情况下对血流量的需要,协调地进行各器官之间的血流分配。
一、神经调节
心肌和血管平滑肌接受自主神经支配。机体对心血管活动的神经调节是通过各种心血管反射实现的。
(一)心脏和血管的神经支配
1.心脏的神经支配支配心脏的传出神经为心交感神经和心迷走神经。
(1)心交感神经及其作用:心交感神经的节前神经元位于脊髓第1-5胸段的中间外侧柱,其轴突末梢释放的递质为乙酰胆碱,后者能激活节后神经元膜上的N型胆碱能受体。心交感节后神经元位于星状神经节或颈交感神经节内。节后神经元的轴突组织心脏神经丛,支配心脏各个部分,包括窦房结、房室交界、房室束、心房肌和心室肌。
在动物实验中看到,两侧心交感神经对心脏的支配有所差别。支配窦房结的交感纤维主要来自右侧心交感神经,支配房室交界的交感主要来自左侧心交感神经。在功能上,右侧心交感神经兴奋时以引起心率加快的效应为主,而左侧心交感神经兴奋则以加强心肌收缩能力的效应为主。
心交感节后神经元末梢释放的递质为去甲肾上腺素,与心肌细胞膜上的β型肾上腺素能受体结合,可导致心率加快,房室交界的传导加快,心房肌和心室肌的收缩能力加强。这些效应分别称为正性变时作用、正性变传导作用和正性变力作用。刺激心交感神经可使心缩期缩短,收缩期室内压上升的速率加大;室内压峰值增高,心舒早期室内压下降的速率加大。这些变化还有利于心室在舒张期的充盈。交感神经末梢释放的去甲肾上腺素和循环血液中的儿茶酚胺都能作用于心肌细胞膜的β肾上腺素能受体,从而激活腺苷酸环化酶,使细胞内cAMP的浓度升高,继而激活蛋白激酶和细胞内蛋白质的磷酸化过程,使心肌膜上的钙通道激活,故在心肌动作电位平台期Ca2+的内流增加,细胞内肌浆网释放的Ca2+也增加,其最终效应是心肌收缩能力增强,每搏作功增加。交感神经兴奋引起的正性变传导作用可使心室各部分肌纤维的收缩更趋同步化,这也有利于心肌收缩力的加强。
心交感神经对心肌的效应,主要是通过β肾上腺素能受体实现的。但心肌也有α肾上腺素能受体。激活心肌的α肾上腺素能受体主要引起正性变力效应,而心率的变化则不显著;另外,室内压上升和下降的速率并无明显加快,故心肌的收缩期延长。心肌α肾上腺素能受体的生理功能还不很清楚,有人认为,当β肾上腺素能受体功能受损时(例如长期使用β肾上腺素能受体拮抗剂),心肌α肾上腺素能受体可继续对交感神经和儿茶酚胺发生反应。在病理情况下,心肌α肾上腺素能受体可能在心肌缺血后再灌注引起的心律失常中起一定的作用。
(2)心迷走神经及其作用:支配心脏的副交感神经节前纤维行走于神经干中。这些节前神经元的细胞体位于延髓的迷走神经背核和疑核,在不同的动物中有种间差异。在胸腔内,心迷走神经纤维和心交感神经一起组成心脏神经丛,并和交感纤维伴行进入心脏,与心内神经节细胞发生突触联系。心迷走神经的节前和节后神经元都是胆碱能神经元。节后神经纤维支配窦房结、心房肌、房室交界、房室速及其分支。心室肌也有迷走神经支配,但纤维末梢的数量远较心房肌中为少。两侧心迷走神经对心脏的支配也有差别,但不如两侧心交感神经支配的差别显著。右侧迷走神经对窦房结的影响占优势;左侧迷走神经对房室交界的作用占优势。
心迷走神经节后纤维末梢释放的乙酰胆碱作用于心肌细胞膜的M型胆碱能受体,可导致心率减慢,心房肌收缩能力减弱,心房肌不应期缩短,房室传导速度减慢,即具有负性变时、变力和变传导作用。刺激迷走神经时也能使心室肌收缩减弱,但其效应不如心房肌明显。迷走神经减弱心肌收缩能力的机制是由于其末梢释放的乙酰胆碱作用于M胆碱能受体后,可使腺苷酸环化酶抑制,因此细胞内cAMP浓度降低,肌浆网释放Ca2+减少(参见本章第二节)。
一般说来,心迷走神经和心交感神经对心脏的作用是相对抗的。但是当两者同时对心脏发生作用时,其总的效应并不等于两者分别作用时发生效应的代数和。在多数情况下,心迷走神经的作用比交感神经的作用占有较大的优势。在动物实验中如同时刺激迷走神经和心交感神经,常出现心率减慢效应。其机制比较复杂。此外,在交感神经末梢上有接头前M型胆碱能受体,在迷走神经末梢上有接头前α肾上腺素能受体。迷走神经末梢释放的乙酰胆碱可作用于交感神经末梢的M型胆碱能受体,使交感神经末梢释放递质减少;交感神经末梢释放的去甲肾上腺素也可作用于迷走神经末梢的α肾上腺素能受体,使迷走神经末梢释放递质减少。这种通过接头前受体影响神经末梢递质释放的过程称为递质释放的接头前(或突触前)调制。
(3)支配心脏的肽能神经元:用免疫细胞化学方法证明,心脏中存在多种神经纤维,如神经肽Y、血管活性肠肽、降钙素基因相关肽、阿片肽等。现已知一些肽类递质可与其它递质,如单胺和乙酰胆碱,共存于同一神经元内,并共同释放。目前对于分布在心脏的肽神经元的生理功能还不完全清楚,但心脏内肽能神经纤维的存在说明这些肽类递质也可能参与对心肌和冠状血管作用,降钙素基因相关肽有加快心率的作用等。
2.血管的神经支配除真毛细血管外,血管壁都有平滑肌分布。不同血管的平滑肌的生理特性有所不同,有些血管平滑肌有自发的肌源性活动,而另一些血管平滑肌很少有肌源性活动。但绝大多数血管平滑肌都受局部组织代谢产物影响。支配血管平滑肌的神经纤维可分为缩血管神经纤维和舒血管神经纤维两大类,两者又统称为血管运动神经纤维。
(1)缩血管神经纤维:缩血管神经纤维都是交感神经纤维,故一般称为并感缩血管纤维,其节前神经元位于脊髓胸、腰段的中间外侧柱内,末梢释放的递质为乙酰胆碱。节后神经元位于椎旁和椎前神经节内,末梢释放的递质为去甲肾上腺素。血管平滑肌细胞有α和β两类肾上腺素能受体。去甲肾上腺素与α肾上腺素能受体结合,可导致血管平滑肌收缩;与β肾上腺素能受体结合,则导致血管平滑肌舒张。去甲肾上腺素与α肾上腺素能受体结合的能力较与β受体结合的能力强,故缩血管纤维兴奋时引起缩血管效应。
体内几乎所有的血管都受交感缩血管纤维支配,但不同部位的血管中缩血管纤维分布的密度不同。皮肤血管中缩血管纤维分布最密,骨骼肌和内脏的血管次之,冠状血管和脑血管中分布较少。在同一器官中,动脉中缩血管纤维的密度高于静脉,微动脉中密度最高,但毛细血管前括约肌中神经纤维分布很少。
人体内多数血管只接受交感缩血管纤维的单一神经支配。在安静状态下,并感缩血管纤维持续发放约1-3次/秒的低频冲动,称为交感缩血管紧张,这种紧张性活动使血管平滑肌保持一定程度的收缩状态。当交感缩血管紧张增强时,血管平滑肌进一步收缩;交感缩血管紧张减弱时,血管平滑肌收缩程度减低,血管舒张。在不同的生理状况下,交感缩血管纤维的放电频率在每秒低于1次至每秒8-10次的范围内变动。这一变动范围足以使血管口径在很大范围内发生变化,从而调节不同器官的血流阻力和血流量。当支配某一器官血管床的并感缩血管纤维兴奋时,可引起该器官血管床的血流阻力增高,血流量减少;同时该器官毛细血管前阻力和毛细血管后阻力的比值增大,使毛细血管血压降低,组织液的生成减少而有利于重吸收;此外,该器官血管床的容量血管收缩,器官内的血容量减少。
近年来,用免疫细胞化学等方法证明,缩血管纤维中有神经肽Y与去甲肾上腺素共存,神经兴奋时两者可共同释放。神经肽Y具有极强烈的缩血管效应。
(2)舒血管神经纤维;体内有一部分血管除接受缩血管纤维支配外,还接受舒血管纤维支配。舒血管神经纤维主要有以下几种:
1)交感舒血管神经纤维:有些动物如狗和猫,支配骨骼肌微动脉的交感神经中除有缩血管纤维外,还有舒血管纤维。交感舒血管纤维末梢释放的递质为乙酰胆碱,阿托品可阻断其效应。交感舒血管纤维在平时没有紧张性活动,只有在动物处于情绪激动状态和发生防御反应时才发放冲动,使骨骼肌血管舒张,血流量增多。在人体内可能也有交感舒血管纤维存在。
2)副交感舒血管神经纤维:少数器官如脑膜、唾液腺、胃肠外分泌腺和外生殖器等,其血管平滑肌除接受交感缩血管纤维支配外,还接受副交感舒血管纤维支配。例如面神经中有支配软脑膜血管的副交感纤维,迷走神经中有支配肝血管的副交感纤维,盆神经中有支配盆腔器官和外生殖器血管的副交感纤维等等。副交感舒血管纤维末梢释放的递质为乙酰胆碱,后者与血管平滑肌的M型胆碱能受体结合,引起血管舒张。副交感舒血管纤维的活动只对器官组织局部血流起调节作用,对循环系统总的外周阻力的影响很小。
3)脊髓背根舒血管纤维:皮肤伤害性感觉传入纤维在外周末梢可发生分支。当皮肤受到伤害性刺激时,感觉冲动一方面沿传入纤维向中枢传导,另一方面可在末梢分叉处沿其它分支到达受刺激部位邻近的微动脉,使微动脉舒张,局部皮肤出现红晕。这种仅通过轴突外周部位完成的反应,称为轴突反射。这种神经纤维也称背根舒血管纤维,其释放的递质还不很清楚,有人认为是P物质,也有人认为可能是组胺或ATP。近年来用免疫细胞化学方法证明,脊神经节感觉神经元中有降钙素基因相关肽与P物质共存;另外,在许多血管周围常可看到有降钙素基因相关肽神经纤维分布。降钙素基因相关肽有强烈的舒血管效应,故有人认为这种多肽可能是引起轴突反射舒血管效应的递质。
4)血管活性肠肽神经元:有些自主神经元内有血管活性肠肽和乙酰胆碱共存,例如支配汗腺的交感神经元和支配颌下腺的副交感神经元等。这些神经元兴奋时,其末稍一方面释放乙酰胆碱,引起腺细胞分泌;另一方面释放血管活性肠肽,引起舒血管效应,使局部组织血流增加。
(二)心血管中枢
神经系统对心血管活动的调节是通过各种神经反射来实现的。在生理学中将与控制心血管活动有关的神经元集中的部位称为心血管中枢。控制心血管活动的神经元并不是只集中在中枢神经系统的一个部位,而是分布在中枢神经系统从脊髓到大脑皮层的各个水平上,它们各具不同的功能,又互相密切联系,使整个心血管系统的活动协调一致,并与整个机体的活动相适应。
1.延髓心血管中枢一般认为,最基本的心血管中枢位于延髓。这一概念最早是在19世纪70年代提出的。它基于以下的动物实验结果:在延髓上缘横断脑干后,动物的血压并无明显的变化,刺激坐骨神经引起的升血压反射也仍存在;但如果将横断水平逐步移向脑干尾端,则动脉血压就逐渐降低,刺激坐骨神经引起的升血压反射效应也逐渐减弱。当横断水平下移至延髓闩部时,血压降低至大约5.3kPa(40mmHg)。这些结果说明,心血管的正常的紧张性活动不是起源于脊髓,而是起源于延髓,因为只要保留延髓及其以下中枢部分的完整,就可以维持心血管正常的紧张性活动,并完成一定的心血管反射活动。
延髓心血管中枢的神经元是指位于延髓内的心迷走神经元和控制心交感神经和交感缩血管神经活动的神经元。这些神经元在平时都有紧张性活动,分别称为心迷走紧张、心交感紧张和交感缩血管紧张。在机体处于安静状态时,这些延髓神经元的紧张性活动表现为心迷走神经纤维和交感神经纤维持续的低频放电活动。
一般认为,延髓心血管中枢至少可包括以下四个部位的神经元:
(1)缩血管区:引起交感缩血管神经正常的紧张性活动的延髓心血管神经元的细胞体位于延髓头端的腹外侧部,称为C1区。这些神经元内含有肾上腺素,它们的轴突下行到脊髓的中间外侧柱。心交感紧张也起源于此区神经元。
(2)舒血管区:位于延髓尾端腹外侧部A1区(即在C1区的尾端)的去甲肾上腺素神经元,在兴奋时可抑制C1区神经元的活动,导致交感缩血管紧张降低,血管舒张。
(3)传入神经接替站:延髓孤束核的神经元接受由颈动脉窦、主动脉弓和心脏感受器经舌咽神经和迷走神经传入的信息,然后发出纤维至延髓和中枢神经系统其它部位的神经元,继而影响心血管活动。
(4)心抑制区:心迷走神经元的细胞体位于延髓的迷走神经背核和疑核。
2.延髓以上的心血管中枢在延髓以上的脑干部分以及大脑和小脑中,也都存在与心血管活动有关的神经元。它们在心血管活动调节中所起的作用较延髓心血管中枢更加高级,特别是表现为对心血管活动和机体其它功能之间的复杂的整合。例如下丘脑是一个非常重要的整合部位,在体温调节、摄食、水平衡以及发怒、恐惧等情绪反应的整合中,都起着重要的作用。这些反应都包含有相应的心血管活动的变化。在动物实验中可以看到,电刺激下丘脑的一些区域,可以引起躯体肌肉以及心血管、呼吸和其它内脏活动的复杂的变化。这些变化往往是通过精细整合的,在生理功能上往往是相互协调的。例如电刺激下丘脑的“防御反应区”,可立即引起动物的警觉状态,骨骼肌肌紧张加强,表现出准备防御的姿势等行为反应,同时出现一系列心血管活动的改变,主要是心率加快,心搏加强,心输出量增加,皮肤和内脏血管收缩,骨骼肌血管舒张,血压稍有升高。这些心血管反应显然是与当时机体所处的状态相协调的,主要是使骨骼肌有充足的血液供应,以适应防御、搏斗或逃跑等行为的需要。
大脑的一些部位,特别是边缘系统的结构,如颞极、额叶的眶面、扣带回的前部、杏仁、隔、海马等,能影响下丘脑和脑干其它部位的心血管神经元的活动,并和机体各种行为的改变相协调。大脑新皮层的运动区兴奋时,除引起相应的骨骼肌收缩外,还能引起该骨骼肌的血管舒张。刺激小脑的一些部位也可引起心血管活动的反应。例如刺激小脑顶核可引起血压升高,心率加快。顶核的这种效应可能与姿势和体位改变时伴随的心血管活动变化有关。
(三)心血管反射
当机体处于不同的生理状态如变换姿势、运动、睡眠时,或当机体内、外环境发生变化时,可引起各种心血管反射,使心输出量和各器官的血管收缩状况发生相应的改变,动脉血压也可发生变动。心血管反射一般都能很快完成,其生理意义在于使循环功能能适应于当时机体所处的状态或环境的变化。
1.颈动脉窦和主动脉弓压力感觉反射当动脉血压升高时,可引起压力感受性反射,其反射效应是使心率减慢,外周血管阻力降低,血压回降。因此这一反射曾被称为降压反射。
(1)动脉压力感觉器:压力感受性反射的感受装置是位于颈动脉窦和主动脉弓血管外膜下的感觉神经末梢,称为动脉压力感受器(图4-25)。动脉压力感觉器并不是直接感觉血压的变化,而是感觉血管壁的机械牵张程度。当动脉血压升高时,动脉管壁被牵张的的程度就升高,压力感觉器发放的神经冲动也就增多。在一定范围内,压力感觉器的传入冲动频率与动脉管壁扩张程度成正比。由图4-26可见,在一个心动周期内,随着动脉血压的波动,窦神经的传入冲动频率也发生相应的变化。
图4-25颈动脉窦区与主动脉弓区的压力感受器与化学感受器
(2)传入神经和中枢联系:颈动脉窦压力感受器的传入神经纤维组成颈动脉窦神经。窦神经加入舌咽神经,进入延髓,和孤束核的神经元发生突触联系。主动脉弓压力感受器的传入神经纤维行走于迷走神经干内,然后进入延髓,到达孤束核。兔的主动脉弓压力感受器传入纤维自成一束,与迷走神经伴行,称为主动脉神经。
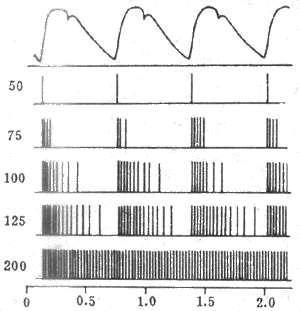
图4-26单根窦神经压力感受器传入纤维在不同动脉压时的放电图
中最上方为主动脉血压波,左侧的数字为主动脉平均压(mmHg,1mmHg=0.133kPa)
压力感受器的传入神经冲动到达孤束核后,可通过延髓内的神经通路使延髓端腹外侧部C1区的血管运动神经元抑制从而使交感神经紧张性活动减弱;孤束核神经元还与延髓内其它神经核团以及脑干其它部位如脑桥、下丘脑等的一些神经核团发生联系,其效应也是使交感神经紧张性活动减弱。另外,压力感受器的传入冲动到达孤束核后还与迷走神经背核和疑核发生联系,使迷走神经的活动加强。
(3)反射效应:动脉血压升高时,压力感受器传入冲动增多,通过中枢机制,使心迷走紧张加强,心交感紧张和交感缩血管紧张减弱,其效应为心率减慢,心输出量减少,外周血管阻力降低,故动脉血压下降。反之,当动脉血压降低时,压力感受器传入冲动减少,使迷走紧张减弱,交感紧张加强,于是心率加快,心输出量增加,外周血管阻力增高,血压回升。
在动物实验中可将颈动脉窦区和循环系统其余部分隔离开来,但仍保留它通过窦神经与中枢的联系。在这样的制备中,人为地改变颈动脉窦区的灌注压,就可以引起体循环动脉压的变化,并画出压力感受性反射功能曲线(见图4-27)。由图可见,压力感受性反射功能曲线的中间部分较陡,向两端渐趋平坦。这说明当窦内压在正常平均动脉压水平(大约13.3kPa或100mmHg)的范围内发生变动时,压力感受性反射最为敏感,纠正偏离正常水平的血压的能力最强,动脉血压偏离正常水平愈远,压力感受性反射纠正异常血压的能力愈低。
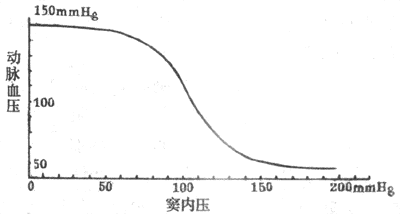
图4-27 在实验中测得的颈动脉窦内压力与动脉血压的关系(1mmHg=0.133kPa)
(4)压力感受性反射的生理意义:压力感受性反射在心输出量、外周血管阻力、血量等发生突然变化的情况下,对动脉血压进行快速调节的过程中起重要的作用,使动脉血压不致发生过分的波动,因此在生理学中将动脉压力感受器的传入神经称为缓冲神经。在动物实验中可看到,正常狗24小时内动脉血压的变化范围一般在平均动脉压(约13.3kPa或100mmHg)约1.3-2.0kPa(10-15mmHg)以内;而在切除两侧缓冲神经的狗,血压经常出现很大的波动,其变动范围可超过平均动脉压上下各6.7kPa(50mmHg)。但是,在切除缓冲神经的动物,一天中血压的平均值并不明显高于正常,因此认为压力感受性反射在动脉血压的长期调节中并不起重要作用。在慢性高血压患者或实验性高血压动物中,压力感受性反射功能曲线向右移位。这种现象称为压力感受性反射的重调定(resetting),表示在高血压的情况下压力感受性反射的工作范围发生改变,即在较正常高的血压水平上进行工作,故动脉血压维持在比较高的水平。压力感受性反射重调定的机制比较复杂。重调定可发生在感受器的水平,也可发生在反射的中枢部分。
2.心肺感受器引起的心血管反射在心房、心室和肺循环大血管壁存在许多感受器,总称为心肺感受器,其传入神经纤维行走于迷走神经干内。引起心肺感受器兴奋的适宜刺激有两大类。一类是血管壁的机械牵张。当心房、心室或肺循环大血管中压力升高或血容量增多而使心脏或血管壁受到牵张时,这些机械或压力感受器就发生兴奋。和颈动脉窦、主动脉弓压力感受器相比较,心肺感受器位于循环系统压力较低的部分,故常称之为低压力感受器,而动脉压力感受器则称为高压力感受器。在生理情况下,心房壁的牵张主要是由血容量增多而引起的,因此心房壁的牵张感受器也称为容量感受器。另一类心肺感受器的适宜刺激是一些化学物质,如前列腺素、缓激肽等。有些约物如藜芦碱等也能刺激心肺感受器。
大多数心肺感受器受刺激时引起的反射效应是交感紧张降低,心迷走紧张加强,导致心率减慢,心输出量减少,外周血管阻力降低,故血压下降。在多种实验动物中,心肺感受器兴奋时肾交感神经活动的抑制特别明显,使肾血流量增加,肾排水和排钠量增多。这表明心肺感受器引起的反射在血量及体液的量和成分的调节中有重要的生理意义。心肺感受器引起的反射的传出途径除神经外还有体液的成分。心肺感受器的传入冲动可抑制血管升压素的释放。血管升压素的减少导致肾排水增多(见后)。
3.颈动脉体和主动脉体化学感受性反射在颈总动脉分叉处和主动脉弓区域,存在一些特殊的感受装置,当血液的某些化学成分发生变化时,如缺氧、CO2分压过高、H+浓度过高等,可以刺激这些感受装置。因此这些感受装置被称为颈动脉体和主动脉体化学感受器。这些化学感受器受到刺激后,其感觉信号分别由颈动脉窦神经和迷走神经传入至延髓孤束核,然后使延髓内呼吸神经元和心血管活动神经元的活动发生改变。
化学感受性反射的效应主要是呼吸加深加快(详见第五章)。在动物实验中人为地维持呼吸频率和深度不变,则化学感受器传入冲动对心血管活动的直接效应是心率减慢,心输出量减少,冠状动脉舒张,骨骼肌和内脏血管收缩。由于外周血管阻力增大的作用超过心输出量减少的作用,故血压升高。在动物保持自然呼吸的情况下,化学感受器受刺激时引起的呼吸加深加快,心输出量增加,外周血管阻力增大,血压升高。
化学感受性反射在平时对心血管活动并不起明显的调节作用。只有在低氧、窒息、失血、动脉血压过低和酸中毒情况下才发生作用。
4.躯体感受器引起的心血管反射刺激躯体传入神经时可以引起各种心血管反射。反射的效应取决于感受器的性质、刺激的强度和频率等因素。用低至中等强度的低频电脉冲刺激骨骼肌传入神经,常可引起降血压效应;而用高强度高频率电刺激皮肤传入神经,则常引起升血压效应。在平时,肌肉活动,皮肤冷、热刺激以及各种伤害性刺激都能引起心血管反射活动。中医针刺治疗某些心血管疾病的生理基础,就在于激活肌肉或皮肤的一些感受器传入活动,通过中枢神经系统内复杂的机制,使异常的心血管活动得到调整。
5.其它内脏感受器引起的心血管反射扩张肺、胃、肠、膀胱等空腔器官,挤压睾丸等,常可引起心率减慢和外周血管舒张等效应。这些内脏感受器的传入神经纤维行走于迷走神经或交感神经内。
6.脑缺血反应当脑血流量减少时,心血管中枢的神经元可对脑缺血发生反应,引起交感缩血管紧张显著加强,外周血管高度收缩,动脉血压升高,称为脑缺血反应。
(四)心血管反射的中枢整合型式
在过去较长的时期中,生理学的一个概念是认为整个交感神经系统或者一起兴奋,或者一起抑制。但后来认识到,不同部分的交感神经、副交感神经的活动都是有分化的。具体地说,对于某种特定的刺激,不同部分的交感神经的反应方式和程度是不同的,即表现为一定整合型式的反应,使各器官之间的血流分配能适应机体当时功能活动的需要。例如当动物的安全受到威胁而处于警觉、戒备状态时,可出现一系列复杂的行为和心血管反应,称为防御反应。猫的防御反应表现为瞳孔扩大、竖毛、耳廓平展、弓背、伸爪、呼吸加深、怒叫,最后发展为搏斗或逃跑;伴随防御反应的心血管整合型式,最特征性的是骨骼肌血管舒张,同时心率加快,心输出量增加,内脏和皮肤血管收缩,血压轻度升高。人在情绪激动时也可发生这一整套心血管反应整合型式。肌肉运动时心血管活动的整合型式与防御反应相似,但血管舒张仅发生在进行运动的肌肉,不进行运动的肌肉的血管发生收缩。睡眠时心脏和血管的活动恰好与防御反应时相反,即心率减慢,心输出量稍减少,内脏血管舒张,骨骼肌血管收缩,血压稍降低。
二、体液调节
心血管活动的体液调节是指血液和组织液中一些化学物质对心肌和血管平滑肌的活动发生影响,从而起调节作用。这些体液因素中,有些是通过血液携带的,可广泛作用于心血管系统;有些则在组织中形成,主要作用于局部的血管,对局部组织的血流起调节作用。
(一)肾素-血管紧张素系统
肾素是由肾近球细胞合成和分泌的一种酸性蛋白酶,经肾静脉进入血循环。血浆中的肾素底物,即血管紧张素原,在肾素的作用下水解,产生一个十肽,为血管紧张素I。在血浆和组织中,特别是在肺循环血管内皮表面,存在有血管紧张素转换酶,在后者的作用下,血管紧张素I水解,产生一个八肽,为血管紧张素Ⅱ。血管紧张素Ⅱ在血浆和组织中的血管紧张素酶A的作用下,再失去一个氨基酸,成为七肽血管紧张素Ⅲ。上述过程可由图4-28表示。血管紧张素Ⅱ和血管紧张素Ⅲ作用于血管平滑肌和肾上腺皮质等细胞的血管紧张素受体,引起相应的生理效应。
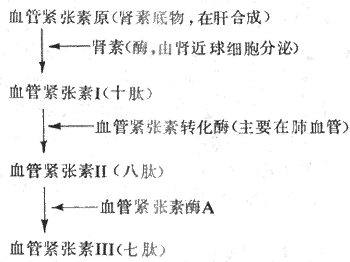
当各种原因引起肾血流灌注减少时,肾素分泌就会增多。血浆中Na+浓度降低时,肾素分泌也增加。肾素分泌受神经和体液机制的调节,详见第八章。
对体内多数组织、细胞来说,血管紧张素I不具有活性。血管紧张素中最重要的是血管紧张素Ⅱ。血管紧张素Ⅱ可直接使全身微动脉收缩,血压升高;也可使静脉收缩,回心血量增多。血管紧张素Ⅱ可作用于交感缩血管纤维末梢上的接头前血管紧张素受体,起接头前调制的作用,使交感神经末梢释放递质增多。血管紧张素Ⅱ还可作用于中枢神经系统内一些神经元的血管紧张素受体,使交感缩血管紧张加强。因此,血管紧张素Ⅱ可以通过中枢和外周机制,使外周血管阻力增大,血压升高。此外,血管紧张素Ⅱ可强烈刺激肾上腺皮质球状带细胞合成和释放醛固酮,后者可促进肾小管对Na+的重吸收,并使细胞外液量增加。血管紧张素Ⅱ还可引起或增强渴觉,并导致饮水行为。血管紧张素Ⅲ的缩血管效应仅为血管紧张素Ⅱ的10%-20%,但刺激肾上腺皮质合成和释放醛固酮的作用较强。
在某些病理情况下,如失血时,肾素-血管紧张素系统的活动加强,并对循环功能的调节起重要作用。
(二)肾上腺素和去甲肾上腺素
肾上腺素和去甲肾上腺素在化学结构上都属于儿茶酚胺。循环血液中的肾上腺素和去甲肾上腺素主要来自肾上腺髓质的分泌。肾上腺素能神经末梢释放的递质去甲肾上腺素也有一小部分进入血液循环。肾上腺髓质释放的儿茶酚胺中,肾上腺素约占80%,去甲肾上腺素约占20%。
血液中的肾上腺素和去甲肾上腺素对心脏和血管的作用有许多共同点,但并不完全相同,因为两者对不同的肾上腺素能受体的结合能力不同。肾上腺素可与α和β两类肾上腺素能受体结合。在心脏,肾上腺素与β肾上腺素能受体结合,产生正性变时和变力作用,使心输出量增加。在血管,肾上腺素的作用取决于血管平滑肌上α和β肾上腺素能受体分布的情况。在皮肤、肾、胃肠、血管平滑肌上α肾上腺素能受体在数量上占优势,肾上腺素的作用是使这些器官的血管收缩;在骨骼肌和肝的血管,β肾上腺素能受体占优势,小剂量的肾上腺素常以兴奋β肾上腺素能受体的效应为主,引起血管舒张,大剂量时也兴奋α肾上腺素能受体,引起血管收缩。去甲肾上腺素主要与α肾上腺素能受体结合,也可与心肌的β1肾上腺素能受体结合,但和血管平滑肌的β2肾上腺素能受体结合的能力较弱。静脉注射去甲肾上腺素,可使全身血管广泛收缩,动脉血压升高;血压升高又使压力感受性反射活动加强,压力感受性反射对心脏的效应超过去甲肾上腺素对心脏的直接效应,故心率减慢。
(三)血管升压素
血管升压素是下丘脑视上核和室旁核一部分神经元内合成的。这些神经元的轴突行走在下丘脑垂体束中并进入垂体后叶,其末梢释放的血管升压素作为垂体后叶激素进入血循环。血管升压素的合成和释放过程也称为神经分泌。
血管升压素在肾集合管可促进水的重吸收,故又称为抗利尿激素(见第八章)。血管升压素作用于血管平滑肌的相应受体,引起血管平滑肌收缩,是已知的最强的缩血管物质之一。在正常情况下,血浆中血管升压素浓度升高时首先出现抗利尿效应;只有当其血浆浓度明显高于正常时,才引起血压升高。这是因为血管升压素能提高压力感受性反射的敏感性,故能缓冲升血压效应。血管升压素对体内细胞外液量的调节起重要作用。在禁水、失水、失血等情况下,血管升压素释放增加,不仅对保留体内液体量,而且对维持动脉血压,都起重要的作用。
(四)血管内皮生成的血管活性物质
多年来一直以为血管内皮只是衬在心脏和血管腔面的一层单层细胞组织;在毛细血管处,通过内皮进行血管内外的物质交换。近年已证实,内皮细胞可以生成并释放若干种血管活性物质,引起血管平滑肌舒张或收缩。
1.血管内皮生成的舒血管物质血管内皮生成和释放的舒血管物质有多种。内皮细胞内的前列环素合成酶可以合成前列环素(也称前列腺素I2,即PGI2)。血管内的搏动性血流对内皮产生的切应力可使内皮释放PGI2,后者使血管舒张。
现在认为,内皮生成的另一类舒血管物质更重要,即内皮舒张因子(endothelium-derivedrelaxing factor, EDRF)。EDRF的化学结构尚未完全弄清,但多数人认为可能是一氧化氮(NO),其前体是L-精氨酸。EDRF可使血管平滑肌内的鸟苷酸环化酶激活,cGMP浓度升高,游离Ca2+浓度降低,故血管舒张。血流对血管内皮产生的切应力可引起EDRF的释放。低氧也可使内皮释放EDRF。此外,内皮细胞表面存在着一些受体,例如P物质受体、5-羟色胺受体、ATP受体、M型胆碱能受体等,这些受体被相应的物质激活后,可释放EDRF。有些缩血管物质,如去甲肾上腺素、血管升压素、血管紧张素Ⅱ等,也可使内皮释放EDRF,后者可减弱缩血管物质对血管平滑肌的直接收缩效应。在离体实验中可看到,将乙酰胆碱作用于内皮完整的血管,引起血管舒张;而将血管内皮去除后,乙酰胆碱则使血管收缩。
2.血管内皮生成的缩血管物质血管内皮细胞也可产生多种缩血管物质,称为内皮缩血因子(endothelum-derived vasoconstrictor factor,EDCF)。近年来研究得较深入的是内皮素。内皮素(endothelin)是内皮细胞合成和释放的由21个氨基酸构成的多肽,是已知的最强烈的缩血管物质之一。给动物注射内皮素可引起持续时间较长的升血压效应。但在升血压之前常先出现一个短暂的降血压过程。有人解释,内皮素也可引起EDRF的释放,故有一短暂的降血压反应。在生理情况下,血管内血流对内皮产生的切应力可使内皮细胞合成和释放内皮素。
(五)激肽释放酶-激肽系统
激肽释放酶是体内的一类蛋白酶,可使某些蛋白质底物激肽原分解为激肽。激肽具有舒血管活性,可参与对血压和局部组织血流的调节。
激肽释放酶可分为两大类,一类存在于血浆,称为血浆激肽释放酶;另一类存在于肾、唾液腺、胰腺等器官组织内,称为腺体激肽释放酶或组织激肽释放酶。激肽原是存在于血浆中的一些蛋白质,分为高分子量激肽原和低分子量激肽原。在血浆中,血浆激肽释放酶作用于高分子量激肽原,使之水解,产生一种九肽,即缓激肽。在肾、唾液腺、胰腺、汗腺以及胃肠粘膜等组织中,腺体激肽释放酶作用于血浆中的低分子量激肽原,产生一种十肽,为赖氨酰缓激肽,也称胰激肽或血管舒张素。后者在氨基肽酶的作用下失去赖氨酸,成为缓激肽。缓激肽在激肽酶的作用下水解失活。
激肽可使血管平滑肌舒张和毛细血管通透性增高;但对其它的平滑肌则引起收缩。在人体和动物实验中证实,缓激肽和血管舒张素是已知的最强烈的舒血管物质。在一些腺体器官中生成的激肽,可以使器官局部的血管舒张,血流量增加。
循环血液中的缓激肽和血管舒张素等激肽也参与对动脉血压的调节,使血管舒张,血压降低。
(六)心钠素
心钠素(cardionatrin)是由心房肌细胞合成和释放的一类多肽。在人的循环血液中,最主要的是一种由28个氨基酸构成的多肽。心钠素可使血管舒张,外周阻力降低;也可使每搏输出量减少,心率减慢,故心输出量减少。心钠素作用于肾的受体,还可以使肾排水和排钠增多,故心钠素也称为心房利尿钠肽(atrial natriuretic peptide)。此外,心钠素还能抑制肾的近球细胞释放肾素,抑制肾上腺球状带细胞释放醛固酮;在脑内,心钠素可以抑制血管升压素的释放。这些作用都可导致体内细胞外液量减少。
当心房壁受到牵拉时,可引起心钠素的释放。在生理情况下,当血容量增多、取头低足高的体位、身体浸入水中(头露出水面)时,血浆心钠素浓度升高,并引起利尿和尿钠排出增多等效应。因此,心钠素是体内调节水盐平衡的一种重要的体液因素。心钠素和另外一些体液因素在血压和水盐平衡的调节中还起相互制约的作用。内皮素和血管升压素也都能刺激心房肌细胞释放心钠素。
(七)前列腺素
前列腺素是一族二十碳不饱和脂肪酸,分子中有个环戊烷,其前体是花生四烯酸或其它二十碳不饱和脂肪酸。全身各部的组织细胞几乎都含有生成前列腺素的前体及酶,因此都能产生前列腺素。前列腺素按其分子结构的差别,可分为多种类型。各种前列腺素对血管平滑肌的作用是不同的,例如前列腺素E2具有强烈的舒血管作用,前列腺素F2α则使静脉收缩。前列环素(即前列腺素I2)是在血管组织中合成的一种前列腺素,有强烈的舒血管作用。
交感缩血管纤维末梢释放递质的过程受前列腺素调制。去甲肾上腺素和血管紧张素Ⅱ等缩血管物质作用于血管平滑肌相应的受体,引起血管平滑肌收缩,同时也使血管平滑肌生成前列腺素E2和前列环素。前列腺素E2和前列环素可使血管平滑肌对去甲肾上腺素和血管紧张素Ⅱ的敏感性降低。另一方面,血管平滑肌生成的前列腺素又可通过神经-平滑肌接头间隙作用于交感神经纤维末梢接头前的前列腺素受体,使交感纤维末梢释放递质减少。可见,前列腺素在交感神经-血管平滑肌接头处起着一种局部负反馈调节作用。
(八)阿片肽
体内的阿片肽有多种。垂体释放的β-内啡肽和促肾上腺皮质激素来自同一个前体。在应激等情况下,β-内啡肽和促肾上腺皮质激素一起被释放入血液。β-内啡肽可使血压降低。β-内啡肽的降血压作用可能主要是中枢性的。血浆中的β-内啡肽可进入脑内并作用于某些与心血管活动有关的神经核团,使交感神经活动抑制,心迷走神经活动加强。内毒素、失血等强烈刺激可引起β-内啡肽释放,并可能成为引起循环休克的原因之一。针刺穴位也可引起脑内阿片肽的释放。这可能是针刺使高血压患者血压下降的机制之一。
除中枢作用外,阿片肽也可作用于外周的阿片受体。血管壁的阿片受体在阿片肽作用下,可导致血管平滑肌舒张。另外,交感缩血管纤维末梢也存在接头前阿片受体,这些受体被阿片肽激活时,可使交感纤维释放递质减少。
(九)组胺
组胺是由组氨酸在脱羧酶的作用下产生的。许多组织,特别是皮肤、肺和肠粘膜的肥大细胞中含有大量的组胺。当组织受到损伤或发生炎症和过敏反应时,都可释放组胺。组胺有强烈的舒血管作用,并能使毛细血管和微静脉的管壁通透性增加,血浆漏入组织,导致局部组织水肿。
三、局部血流调节
体内各器官的血流量一般取决于器官组织的代谢活动,代谢活动愈强,耗氧愈多,血流量也就愈多。器官血流量主要通过对灌注该器官的阻力血管的口径的调节而得到控制。除了前述的神经调节和体液调节机制外,还有局部组织内的调节机制。在不同器官的血管,神经、体液和局部机制三者所起作用的相互关系是不同的,在多数情况下,几种机制起协同作用,但在有些情况下也可起相互对抗的作用。另外,不同器官的血流量变化范围也有较大的差别,功能活动变化较大的器官,如骨骼肌、胃肠、肝、皮肤等,血流量的变化范围较大;脑、肾等器官的血流量则比较稳定,在一定的血压变化范围内,器官血流量可保持稳定。
实验证明,如果将调节血管活动的外部神经、体液因素都去除,则在一定的血压变动范围内,器官、组织的血流量仍能通过局部的机制得到适当的调节。这种调节机制存在于器官组织或血管本身,故也称为自身调节。心脏的泵血功能也有自身的调节机制,已在本章第一节中叙述。关于器官组织血流量的局部调节机制,一般认为主要有以下两类:
(一)代谢性自身调节机制
组织细胞代谢需要氧,并产生各种代谢产物。局部组织中的氧和代谢产物对该组织局部的血流量起代谢性自身调节作用。当组织代谢活动增强时,局部组织中氧分压降低,代谢产物积聚增加。组织中氧分压降低以及多种代谢产物,如CO2、H+离子、腺苷、ATP、K+离子等,都能使局部的微动脉和毛细血管前括约肌舒张。因此,当组织的代谢活动加强(例如肌肉运动)时,局部的血流量增多,故能向组织提供更多的氧,并带走代谢产物。这种代谢性局部舒血管效应有时相当明显,如果同时发生交感缩血管神经活动加强,该局部组织的血管仍舒张。
前面提到,有一些体液因素也可在组织中形成,并对局部的血流量起调节作用,例如激肽、前列腺素、组胺等,由于这些物质都是特殊的体液因素,故在生理学中将它们归在体液调节中。
(二)肌源性自身调节机制
许多血管平滑肌本身经常保持一定的紧张性收缩,称为肌源性活动。血管平滑肌还有一个特性,即当被牵张时其肌源性活动加强。因此,当供应某一器官的血管的灌注压突然升高时,由于血管跨壁压增大,血管平滑肌受到牵张刺激,于是肌源性活动增强。这种现象在毛细血管前阻力血管段特别明显。其结果是器官的血流阻力增大,器官的血流量不致因灌注压升高而增多,即器官血流量能因此保持相对稳定。当器官血管的灌注压突然降低时,则发生相反的变化,即阻力血管舒张,血流量仍保持相对稳定。这种肌源性的自身调节现象,在肾血管表现特别明显,在脑、心、肝、肠系膜和骨骼肌的血管也能看到,但皮肤血管一般没有这种表现。在实验中用罂粟碱、水合氯醛或氰化钠等药物抑制平滑肌的活动后,肌源性自身调节现象也随之消失。
四、动脉血压的长期调节
动脉血压的神经调节主要是在短时间内血压发生变化的情况下起调节作用的。而当血压在较长时间内(数小时,数天,数月或更长)发生变化时,神经反射的效应常不足以将血压调节到正常水平。在动脉血压的长期调节中起重要作用的是肾。具体地说,肾通过对体内细胞外液量的调节而对动脉血压起调节作用。有人将这种机制称为肾-体液控制系统。此系统的活动过程如下:当体内细胞外液量增多时,血量增多,血量和循环系统容量之间的相对关系发生改变,使动脉血压升高;而当动脉血压升高时,能直接导致肾排水和排钠增加,将过多的体液排出体外,从而使血压恢复到正常水平。体内细胞外液量减少时,发生相反的过程,即肾排水和排钠减少,使体液量和动脉血压恢复。
肾-体液控制系统调节血压的效能取决于一定的血压变化能引起多大程度的肾排水排钠变化。实验证明,血压只要发生很小的变化,就可导致肾排尿量的明显变化。血压从正常水平(13.3kPa,100mmHg)升高1.3kPa(10mmHg),肾排尿量可增加数倍,从而使细胞外液量减少,动脉血压下降。反之,动脉血压降低时,肾排尿明显减少,使细胞外液量增多,血压回升。
肾-体液控制系统的活动也可受体内若干因素的影响,其中较重要的是血管升压素和肾素-血管紧张素-醛固酮系统。前已述,血管升压素在调节体内细胞外液量中起重要作用。血管升压素使肾集合管增加对水的重吸收,导致细胞外液量增加。当血量增加时,血管升压素减少,使肾排水增加。血管紧张素Ⅱ除引起血管收缩,血压升高外,还能促使肾上腺皮质分泌醛固酮。醛因酮能使肾小管对Na+的重吸收增加,并分泌K+和H+,在重吸收Na+时也吸收水,故细胞外液量和体内的Na+量增加,血压升高。
总之,血压的调节是复杂的过程,有许多机制参与。每一种机制都在一个方面发挥调节作用,但不能完成全部的、复杂的调节。神经调节一般是快速的、短期的调节,主要是通过对阻力血管口径及心脏活动的调节来实现的;而长期调节则主要是通过肾对细胞外液量的调节实现的。
第五节 器官循环
体内每一器官的血流量取决于主动脉压和中心静脉压之间的压力差,又取决于该器官阻力血管的舒缩状态。由于各器官的结构和功能各不相同,器官内部的血管分布又各有特征,因此其血流量的调节除服从前已述的一般规律外,还有其本身的特点。本节叙过心、肺、脑几个主要器官的血液循环特征。关于肾的血液循环特征,将在第八章叙述。
一、冠脉循环
(一)冠脉循环的解剖特点
心肌的血液供应来自左、右冠状动脉。冠状动脉的主干行走于心脏的表面,其小分支以垂直于心脏表面的方向穿入心肌,并在心内膜下层分支成网。这种分支方式使冠脉血管容易在心肌收缩进受到压迫。左、右冠状动脉及其分支的走向可有多种变异。在多数人中,左冠状动脉主要供左心室的前部,右冠状动脉主要供应左心室的后部和右心室。左冠状动脉的血液流经毛细血管和静脉后,主要经由冠状窦回流入右心房,而右冠动脉的血液则主要经较细的心前静脉直接回流入右心室。另外还有一小部分冠脉血液可通过心最小静脉直接流入左、右心房和心室腔内。
心肌的毛细血管网分布极为丰富。毛细血管数和心肌纤维数的比例为1:1。在心肌横截面上,每平方毫米面积内约有2500-3000根毛细血管。因此心肌和冠脉血液之间的物质交换可能很快地进行。冠状动脉之间有侧支互相吻合。在人类,这种吻合支戊内膜下较多。正常心脏的冠脉侧较细小,血流量很少。因此当冠状动脉突然阻塞时,不易很快建立侧支循环,常可导致心肌梗塞。但如果冠状动脉阻塞是缓慢形成的,则侧支可逐渐扩张,并可建立新的侧支循环,起代偿作用。
(二)冠脉血流的特点
在安静状态下,人冠脉血流量为每百克心肌每分钟60-80ml。中等体重的人,总的冠脉血流量为225ml/min,占心输出量的4%-5%。冠脉血流量的多少主要取决于心肌的活动,故左心室单位克重心肌组织的血流量大于右心室。当心肌活动加强,冠脉达到最大舒张状态时,冠脉血流量可增加到每百克心肌每分钟300-400ml。
由于肮脏血管的大部分分支深埋于心肌内,心脏在每次收缩时对埋于其内的血管产生压迫,从而影响冠脉血流。图4-29示狗的左、右冠状动脉血流在一个心动周期中的变化。在左心室等容收缩期,由于心肌收缩的强烈压迫,左冠状动脉血流急剧减少,甚至发生倒流。在左心室射血期,主动脉压升高,冠状动脉血压也随着升主,冠脉血流量增加。到慢速射血期,冠脉血流量又有下降。心肌舒张时,对冠脉血管的压迫解除,故冠脉血流的阻力显著减小,血流量增加。在等容舒张期,冠脉血流量突然增加,在舒张期的早期达到最高峰,然后逐渐回降,在左心室深层,心肌收缩对冠血流的影响更为明显。左心房收缩时对冠脉血流也可产生一定的影响,但并不显著。一般说来,左心室在收缩期血流量大约只有舒张期的20%-30%。当心肌收缩加强时,心缩期血流量所占的比例更小。由此可见,动脉舒张压的高低和心舒期的长短是影响冠脉血流量的重要因素。体循环外财阻力增大时,动脉舒张压升高,冠脉血流量增多。心率加快时,由于心动周期的缩短主要是心舒期缩短,故冠脉血流量也减少。右心室肌肉比较薄弱,收缩时对血流的影响不如左心室明显。在安静情况下,右心室收缩期的血流量和舒张期的血流量相差不多,或甚至多于后者。
(三)冠脉血流量的调节
对冠脉血流量进行调节的各种因素中,最重要的是心肌本身的代谢水平。交感和副交感神经也支配冠脉血管平滑肌,但它们的调节作用是次要的。
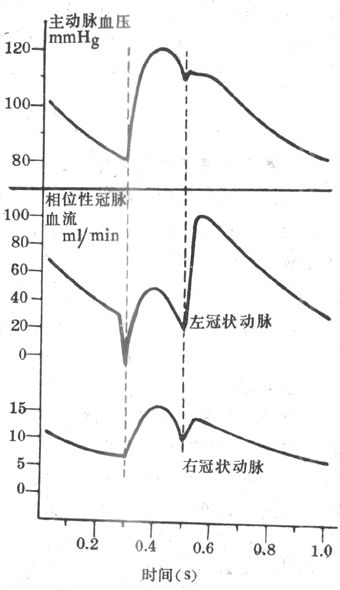
图4-29 一个心动周期中左、右冠状动脉血流变化情况(1mmHg=0.133kPa)
1.心肌代谢水平对冠脉血流量的影响 心肌收缩的能量来源几乎唯一地依靠有氧代谢。心肌因连续不断地进行舒缩,故耗氧量较大,即使在人体处于安静状态时,动脉血流经心脏后,其中65%-75%的氧被心肌摄取。因此心脏的动脉血和静脉血的含氧量差很大,换句话说,心肌提高从单位血液中摄取氧的潜力较不小。在肌肉运动、精神紧张等情况下,心肌代谢活动增强,耗氧量也随之增加。此时,机体主要通过冠脉血管舒张,即增加冠脉血流量来满足心肌对氧的需求。实验证明,冠脉血流量是和心肌代谢水平成正比的。在没有神经支配和循环激素作用的情况下,这种关系仍旧存在。目前认为,心肌代谢增强引起冠脉血管舒张的原因并非低氧本身,而是由于某些心肌代谢产物的增加。在各种代谢产物中,腺苷可能起最重要的作用。当心肌代谢增强而使局部组织中氧分压降低时,心肌细胞中的ATP分解为ADP和AMP。在冠脉血管周围的间质细胞中有5′-核苷酸酶,后者可使AMP分解产生腺苷。腺苷具有强烈的舒张小动脉的作用。腺苷生成的后,在几秒钟内即被破坏,因此不会引起其它器官的血管舒张。心肌的其它代谢产物如H+、CO2乳酸等,虽也能使冠脉舒张,但作用较弱。此外,缓激肽和前列腺素E等体液因素也能使冠脉血管舒张。
2.神经调节 冠状动脉受迷走神经和交感神经支配。迷走神经兴奋对冠状动脉的直接作用是引起舒张。但迷走神经兴奋又使心率减慢,心肌代谢率降低,这些因素可抵消迷走神经对冠状动脉的直接舒张作用。在动物实验中,如果使心率保持不变,则刺激迷走神经引起冠脉舒张。刺激心交感神经时,可激活冠脉平滑肌的α肾上腺素能受体,使血管收缩,但交感神经兴奋又同时激心肌的β肾上腺素能受体,使心率加快,心肌收缩加强,耗氧量增加,从而使冠脉舒张。给予β肾上腺素能受体拮抗剂后,刺激交感神经表现出直接的冠脉收缩反应。冠脉平滑肌上也有β肾上腺素能受体,后者被激活时引起冠脉舒张。交感神经兴奋对冠的β肾上腺至少能受体的激动一般不很明显。一些药物如异丙基肾上腺素对冠脉β肾上腺素能受体作用明显。
总之,在整体条件下,冠脉血流理主要是由心肌本身的代谢水平来调节的。神经因素对冠脉血流的影响在很短时间内就被心肌代谢改变所引起的血流变化所掩盖。
3.激素调节肾上腺素和去甲肾上腺素可通过增强心肌的代谢活动和耗氧量使冠脉血流量增加;也可直接作用于冠脉血管α或β肾上腺素能受体,引起冠脉血管收缩或舒张。甲状腺素增多时,心肌代谢加强,耗氧量增加,使冠状动脉舒张,血流量增加。大剂量血管升压素使冠状动脉收缩,冠脉血流量减少。血管紧张素Ⅱ也能使冠状动脉收缩,冠脉血流量减少。
二、肺循环
肺循环的功能是使血液在流经肺泡进和肺泡之间进行气体交换。呼吸性小支气管以上的呼吸道组织的营养物质收体循环的支气管的末梢之间有吻合支沟通。因此,有一部分支气管静脉血液可经过这些吻合支进入静脉和左心房,使主动脉血液中掺入1%-2%的静脉血。
(一)肺循环的生理特点
右心室的每分输出量和左心室的基本相同。肺动脉及其分支都较粗,管壁较主动脉及其分支薄。肺循环的全部血管都在胸腔内,而胸腔内的压力低于大气压。这些因素使肺循环有与体循环不同的一些特点。
1.血流阻力和血压 肺动脉管壁厚度仅为主动脉的三分之一,其分支短而管径较粗,故肺动脉的可扩张性较高,对血流的阻力较小。肺循环动脉部分总的阻力和静脉部分总的阻力大致相等,故血流在动脉部分的压力降落和在静脉部分的压力降落相等。肺循环毛细血管压大致在右心室压和左心房压数值的中点。由于肺循环血管对血流的阻力小,所以,虽然右心室的每分输出理和左心室每分输出量相等,但肺动脉压远较主动脉压为低。右心室压和肺动脉压可用插入导管的方法直接测量。在正常人,右心室收缩压平均约2.9kPa(22mmHg),舒张压为0-0.13kPa(0-1mmHg)。肺动脉的收缩压和右心室收缩压相同,平均为2.2kPa(22mmHg),舒张压为1.1kPa(8mmHg),平均压约1.7kPa(13mmHg)。用间接方法可测得肺循环行细血管平均压为0.9kPa(7mmHg)。肺循环的终点,即肺静脉和左心房内压为0.13-0.53kPa(1-4mmHg),平均约0.27kPa(2mmHg)。
2.肺的血容量 肺部的血容量约为450ml,占全身血量的9%。由于肺组织和肺血管的可扩张性大,故肺部血容量的变化范围较大。在用力呼气时,肺部血容量减少至约200ml;而在深吸气地可增加到约1000ml。由于肺的血容量较多,而且变化范围较大,故肺循环血管起着贮血库的作用。当机体失血时,肺循环可将一部分血液转移至体循环,起代偿作用。在每一个呼吸周期中,肺循环的血容量也发生周期性的变化,并对左心室输出量和动脉血压发生影响。在吸气时,由腔静脉回流入右心房的血量增多,右心室射出的血量也就增加。由于肺扩张时可将肺循环的血管牵拉扩张,使其容量增大,能容纳较多的血液而由肺静脉回流入左心房的血液则减少。但在几次心搏后,扩张的肺循环血管已被充盈,故肺静脉回流入左心房的血量逐渐增加。在呼气时,发生相反的过程。因此,在吸气开始时,动脉血压下降,到吸气相反相的后半期降至最低点,以后逐渐回升,在呼气相的后半期达到最高点。在呼吸周期中出现的这种血压波动,称为动脉血压的呼吸波。
3.肺循环毛细血管外的液体交换 如前所述,肺循环毛细血管平均约0.9kPa(7mmHg),而血浆胶体渗透压平均3.3kPa(25mmHg),故将组织中的液体吸收入毛细血管的力量较大。现在一般认肺部组织液的压力为负压。这一负压使肺泡膜和毛细血管管壁互相紧密相贴,有利于肺胞和血液之间的气体交换。组织液负压还有利于吸收肺泡内的液体,使肺泡内没有液体积聚。在某些病理情况下,如左心衰竭时,肺静脉压力升高,肺循环毛细血管压也随着升高,就可使液体积聚在肺泡或肺的组织间隙中,形成肺水肿。
(二)肺循环血流量的调节
1.神经调节 肺循环血管受交感神经和迷走神经支配。刺激交感神经对肺血管的直接作用是引起收缩和血流阻力增大。但在整体情况下,交感神经兴奋时体循环的血管收缩,将一部分血液挤入肺循环,使肺循环内血容量增加。循环血液中的儿茶酚胺也有同样的效应。刺激迷走神经可使肺血管舒张。乙酰胆碱也能使肺血管舒张,但在流经肺部后即分解失活。
2.肺泡气的氧分压 肺泡气的氧分压对肺部血管的舒缩活动有明显的影响。急性或慢性的低氧都能使肺部血管收缩,血流阻力增大。引起肺血管收缩的原因是肺泡气的氧分压低而不是血管内血液的氧张力低。当一部分肺泡内气体的氧分压低时,这些肺泡周围的微动脉收缩。在肺泡气的CO2分压升高时,低氧引起的肺部微动脉的收缩更加显著。可见肺循环血管对局部低氧发生的反应和体循环血管不同。肺部血管对低氧发生缩血管反应的机制,目前还不完全清楚。有人推测低氧可能使肺组织产生一种缩血管物质,也有人认为必需有血管内皮存在才能发生这种缩血管反应。肺泡氯低氧引起局部缩血管反应,具有一定的生理意义。当一部分肺泡因通气不足而氧分压降低时,这些肺泡周围的血管收缩,血流减少,而使较多的血液流经通气充足,肺泡气氧分压高的肺泡。假如没有这种缩血管反应,血液流经通气不足的肺泡时,血液不能充分氧合,这部分含氧较低的血液回流入左心房,就会影响体循环血液的含氧量。当吸入气氧分压过低时,例如在高海拔地区,可引起肺循环动脉广泛收缩,血流阻力增大,故肺动脉压显著升高。长期居住在高海拔地区的人,常可因肺动脉高压使右心室负荷长期加重而导致右心室肥厚。
3.血管活性物质对肺血管的影响 肾上腺素、去甲肾上腺素、血管紧张素Ⅱ、血栓素A2、前列腺素F2α等能使肺循环的微动脉收缩。组胺、5-羟色胺能使肺循环静脉收缩,但在流经肺循环后即分解失活。
三、脑循环
脑组织的代谢水平高,血流量较多。在安静情况下,每百克脑的血流量为50-60ml/min。整个脑的血流量约为750ml/min。可见,脑的比重虽仅占体重的约2%,但血流量却占心输出量的15%左右。脑组织的耗氧量也较大。在安静情况下,每百克脑每分钟耗氧3-3.5ml;或者说,整个脑的耗氧量约占全身耗氧量的20%。
(一)脑循环的特点
脑位于颅腔内。颅腔是骨性的,其容积是固定的。颇腔内为脑、脑血管和脑脊液所充满,三者的容积的总和也是固定的。由于脑组织是不可压缩的,故脑血管舒缩程度受到相当的限制,血流量的变化较其它器官的为小。
脑循环的毛细血管壁内皮细胞相互接触紧密,并有一定的重叠,管壁上没有小孔。另外,毛细血管和神经元之间并不直接接触,而为神经胶质细胞怕隔开。这一结构特征对于物质在血液和脑组织之间的扩散起着屏障的作用,称为血-脑屏障(blood-brain barrier)。
(二)脑血流量的调节
1.脑血管的自身调节 脑血流量取决于脑的动、静脉的压力差和脑血管的血流阻力。在正常情况下,颈内静脉压接近于右心房压,且变化不大,故影响血流量的主要因素是颈动脉压。政党情况下脑循环的灌注压为10.6-13.3kPa(80-100mmHg)。平均动脉压降低或颅内压升高都可以使脑的灌注压降低。但当平均动脉压在8.0-18.6kPa(60-140mmHg)范围内变化时,脑血管可通过自身调节的机制使脑血流量保持恒定。平均动脉压降低到8.0kPa(60mmHg)以下时,脑血流量就会显着减少,引起脑的功能障碍。反之,当平均动脉压超过脑血管自身调节的上限时,脑血流量显著增加。
2.CO2和O2分压对及脑血流量的影响血液CO2分压升高时,脑血管舒张,血流量增加。CO2过多时,通过使细胞外液H+浓度升高而使脑血管舒张。过度通气时,CO2呼出过多,动脉血CO2分压过低,脑血流量减少,可引起头晕等症状。血液O2分压降低时,也能使脑血管舒张。
3.脑的代谢对脑血流的影响 脑的各部分的血流量与该部分脑组织的代谢活动程度有关。实验证明,在同一时间内脑的没部分的血流量是不同的,当脑的某一部分活动加强时,该部分的血流量就增多。例如在握拳时,对侧大脑皮层运动区的血流量就增加;阅读时脑的许多区域血流量增加,特别是皮层枕叶和颞叶与语言功能有关的部分血流量增加更为明显。代谢活动加强引起的局部脑血流量增加的机制,可能是通过代谢产物如H+离子、K+离子、腺苷,以及氧分子降低,引起脑血管舒张的。
4.神经调节 颈上神经节发出的去甲云彩上腺素后纤维,其末梢分布至脑的动脉和静脉,并分布至软脑膜的血管,还有少量分布至脑实质的血管。脑实质内的小血管有起自蓝斑去甲肾上腺素神经元的轴突末梢的分布。副交感乙酰胆碱能神经末梢也分布至脑血管。此外,脑血管政治家血管活性肠肽等神经肽纤维末梢分布。神经对脑血管活动的调节作用不很明显。刺激或切除支配脑血管的交感或副交感神经,脑血流量没有明显变化。在多种心血管反射中,脑血流量一般变化都很小。
(三)脑脊液的生成和吸收
脑脊液存在于脑室系统、脑周围的脑池和蛛网膜下腔内,可被视为脑和脊髓的组织液和淋巴。成年人的脑脊液总量约150ml。每天生成的脑脊液约800ml,为脑脊液总量的5-6倍。但同时有等量的脑脊液被吸收入血液,可见脑脊液的更新率较高。
脑脊液主要由侧脑室、第三脑室和第四脑室的脉络丛分泌。侧脑室内的脑脊液经室间孔流入第三脑室,再经过导水管进入第四脑室,然后进入蛛网膜下腔。除脉丛外,室管膜细胞也能分泌脑脊液。软脑膜血管和脑的毛细血管滤过的液体,一部分被重吸收,其余的则沿着血管周围间隙进入蛛网膜下腔,成为脑脊液的一部分。
脑脊液主要通过蛛网膜绒毛被吸收入静脉的血液内。蛛网膜绒毛有活瓣状的细微的管道,其直径为4-12μm。当蛛网膜下腔的压力高于静脉窦的压力时,这些管道就开放。这时,脑脊液(包括其中所含的蛋白质分子甚至小的颗粒如红细胞等)可进入静脉窦血液。当蛛网膜下深的压力低于静脉窦压力时,管道关闭,液体不能由静脉窦向蛛网膜下腔倒流。脑脊液压力的高低取决于其生成和吸收之间的平衡关系。正常人在到卧位时,脑脊液压平均为1.3kPa(10mmHg)。当脑脊液有吸收受到阻碍时,脑脊液压就会升高,并影响脑血流和脑的功能。
脑脊液的主要功能是在脑、脊髓和颇腔、椎管之间起缓冲的作用,有保护性意义。脑浸浴于脑脊液中,由于浮力的作用,使脑的重量减轻到仅50g左右。另外,脑脊液还作为脑和血液之间进行物质交换的中介。脑组织中没有淋巴管,由毛细血管漏出的少量蛋白质,主要经过血管周围间隙进入蛛肉膜下腔的脑脊液中,然后通过蛛网膜绒毛回入血液。
(四)血-脑脊液屏障和血-脑屏障
脑脊液主要是由脉络丛分泌的,但其成分和血浆不脑脊液中蛋白质的含量极微,葡萄糖含量也较血浆为少,但Na+和Mg2+的浓度较血浆中的高,K+、HCO3-和Ca2+的浓度则较血浆中的低。可见,血液和脑脊液之间物质的转运并不是被动的过程,而是主动转运过程。另外,一些大分子物质较难从血液进入脑脊液,仿佛在血液和脑脊液之间存在着某种特殊的屏障,故称之为血-脑脊液屏障(blood-cerebrospinal fluid barrier)。这种屏障对不同物质的通透性上不同的。例如O2、CO2等脂溶性物质可很容易地通过屏障,但许多离子的通透性则较低。血-脑脊液屏障的基础是无孔的毛细血管壁和脉?
■[此处缺少一些内容]■
的浓度较低,即使在实验中使血浆K+浓度加倍,脑脊液中K+浓度仍能保持在正常水平。因此脑内神经元的兴奋性不会因血浆中K+浓度的变化而发生明显的变化。由于血-脑屏障的存在,循环血液中的乙酰胆碱、去甲肾上腺素、多巴胺、甘氨酸等物质就不易进入脑,否则,血浆中这些物质浓度的改变将会明显地扰乱脑内神经元的政党功能活动。
需要指出,脑的某些部分,如下丘脑第三脑室周围和延髓后缘区等处的室周器官,血-脑屏障比较薄弱,毛细血管壁对许多物质的通透性高于脑的其它部分。因此循环血液中的有些物质,如血管紧张素Ⅱ和其它肽类物质,可以在这些部位进入脑内,作用于相应的受体,引起各种效应。另外,当脑组织发生缺氧、损伤等情况以及在脑肿瘤部位,毛细血管壁的通透性增加,故平时不易透过血-脑屏障的物质进入受损部位的脑组织。在临床上可以用同位素标记的白蛋白注入体内,这些蛋白质进入正常脑组织的速度很慢,但较易进入脑肿瘤组织,因此可用这种方法匀来检查脑瘤的部位。在用药物治疗神经系统疾病时,必须明确所用的药物是否容易通过血-脑屏障。
在脑室系统,脑脊液和脑组织之间为室管所分隔;在脑的表面,脑脊液和脑组织之间为软脑膜所分隔。室管膜和软脑膜的通透性很高,脑脊液中的物质很容易通过室管膜或软脑膜进入脑组织。因此,在临床上可将不易通过血-脑屏障的药物直接注入脑脊液,使之能较快地进入脑组织。
参考资料
1.徐丰彦,张镜如主编,人体生理学。北京:人民卫生出版社,1989
2.何瑞荣主编,心血管生理学 北京:人民卫生出版社,1987
3.骆鸿,何瑞荣,延髓腹外侧部对心血管活动的调节。生理科学进展1987;18:224-229
4.姚泰,中枢神经系统对血压的调节,生理科学进展,1989;20:276-283
5.韩济生主编,神经科学纲要,第47章,北京:北京医科大学、中国协和医科大学联合出版社,1983
6.Little RC.Physiology of the Heart andCirculation3rd ed,Year Book Medical Publishers,In,Chica-go,1985
7.Fozzard HA etal.The Heart and Cardiovascular Systen-Scientific Foundations,Raven Press,NewYork,1986
8.Noble D.Thesurprising teart, a revieq of recent progress in cardiac electrophysiology. JPhysiol 1984;353:1-50
9.Reuter H.Ionchannels in cardiac cell menbrane.Ann Rev Physiol 1984;46:473-484
10.Irisawa H et al.Pacemaker currents in mammalian nodal cells .J Mol cell Cardiol 1984;16:777-781
11.WinegradS.Regulation of cardiac contractle proteins.Circ Res 1984;55:565-574
12.Siegl PKS.Overviewof cardiac inotropic mechanisms.J Cardiovasc Pharmacol 1986;8(suppl 9):S1-S10
13 .Carafoli EIntracellular calcium homeostasis.Ann Rev Biochem 1987;56:395-433
14.Brown JH etal.Phosphoinositide-generated second messengers in cardiac signaltransduction.Trends in Cardiovascular Medicine 1922;2:199-213
15.BallermannVJ,Zeidel ML,Gunning ME,Brenner BM,Vasoactive piptides and the kidney.in Kidney4thed, Brenner BM, Rector Jr FC ed,Chapter 14,Wb SaundersCo,Philadelphia,1991
16.Cowley JrAW,Long-term control of arterial blood pressure.Physiol Rev 1992;72:231-300
17.Guyton AC.Textbookof Medical Physiology 8thed,Wb Saunders Co,Philadelphia,1991
18.JohnsonLR.Essential Medical Physiology,Raven Press,New York,1992
19.Mione MC,RalevicV,Burnstock G.Peptdes and vasomotor mechanisms.Pharmac Ther 1990;46:429-468
20 Patton HD,FuchsAF,Hille B,Scher AM,Steiner R.Textbook of Physiology 21sted,WBSaunders Co,Philadelphia,1989
第五章 呼吸
机体与外界环境之间的气体交换过程,称为呼吸。通过呼吸,机体从大气摄取新陈代谢所需要的O2,排出所产生的CO2,因此,呼吸是维持机体新陈代谢和其它功能活动所必需的基本生理过程之一,一旦呼吸停止,生命也将终止。
在高等动物和人体,呼吸过程由三个相互衔接并且同进进行的环节来完成(图5-1):外呼吸或肺呼吸,包括肺通气(外界空气与肺之间的气体交换过程)和肺换气(肺泡与肺毛细血管之间的气体交换过程);气体在血液中的运输;内呼吸或组织呼吸,即组织换气(血液与组织、细胞之间的气体交换过程),有时也将细胞内的氧化过程包括在内。可见呼吸过程不仅依靠呼吸系统来完成,还需要血液循环系统的配合,这种协调配合,以及它们与机体代谢水平的相适应,又都受神经和体液因素的调节。
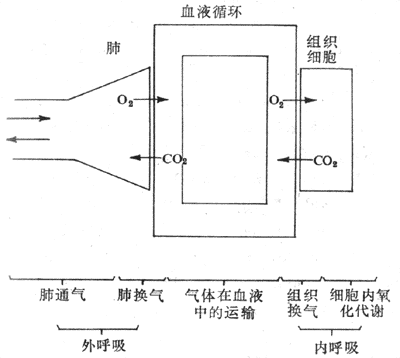
图5-1 呼吸全过程示意图
第一节 肺 通 气
肺通气(pulmonary ventilation)是肺与外界环境之间的气体交换过程。实现肺通气的器官包括呼吸道、肺泡和胸廓等。呼吸道是沟通肺泡与外界的通道;肺泡是肺泡气与血液气进行交换的主要场所;而胸廓的节律性呼吸运动则是实验通气的动力。
一、呼吸道的主要功能
呼吸首(气道)包括鼻、咽、喉(上呼吸道)和气管、支气管及其在肺内的分支(下呼吸道)。随着呼吸道的不断分支,其结构和功能均发生一系列变化,气道数目增多,口径减小,总横断面积增大,管壁变薄,这些变化有重要的生理意义。
(一)调节气道阻力
通过调节气道阻力从而调节进出肺的气体的量、速度和呼吸功(详见肺通气原理)。
(二)保护功能
环境气温、湿度均不恒定,而且可含尘粒和有害气体,这些都交危害机体健康。
但是,呼吸道具有对吸入气体进入加温、湿润、过滤、清洁作用和防御反射等保护功能。
1.加温湿润作用 主要在鼻和咽,而气管和支气管的作用较小。一般情况下,外界空气的温度和温度都较肺同为低。由于鼻、咽粘膜有丰富的血流,并有粘液腺分泌粘液,所以吸入气在达气管时已被加温和被水蒸气所饱和,变为温暖而湿润的气体进入肺泡。如果外界气温高于体温,则通过呼吸道血流的作用,也可以使吸入气的温度下降到体温水平。呼吸道的这种空气调节功能对肺组织有重要的保护作用。经气管插管呼吸的病人,失去了呼吸道的空气调节功能,可使呼吸道上皮、纤毛及腺体等受到损伤,因此应给病人呼吸湿润的空气为宜。
2.过滤清洁作用 通常通过呼吸道的过滤和清洁作用,阻挡和清除了随空气进入呼吸道的颗粒、异物,使进入肺泡的气体几乎清洁无菌。呼吸道有各种不同的机制防止异物到达肺泡。其一在上呼吸道。鼻毛可以阻挡较大颗粒进入,而鼻甲的形状则使许多颗粒直接撞击在粘膜上或因重力而沉积在粘膜上。这样,直径大于10μm的颗粒几乎完全从鼻腔空气中被清除掉。其二在气管、支气管和细支气管。直径在2-10μm的颗粒可通过鼻腔而进入下呼吸道,但这里管壁粘膜有分泌粘液的杯状细胞和纤行上皮细胞。所分泌的沾液覆盖在纤毛上。许多纤毛有力地、协调地和有节奏地摆动,将粘液层和附着于其上的颗粒向喉咽方向移动。每次摆动可移动粘液层达16μm,若每秒钟纤毛摆动20次,则每分钟可使粘液层移动约19mm。纤毛推动粘液层及所附着的颗粒到达咽部后,或被吞咽或被咳出。吸入气干燥或含有刺激性物质,如二氧化硫等,可以损害纤毛的运动,影响呼吸道的防御功能。其三是巨噬细胞。直径小于2μm的小颗粒可以进入呼吸性细支气管、肺泡管和肺泡,巨噬细胞可以吞噬吸入的颗粒和细菌,然后带着它的吞噬物向上游走到细支气管壁上的粘液层,随粘液排出。肺泡巨噬细胞生活在氧分压较市制肺泡中,当通气量减少或氧分压降低时,其功能将减退。此外,呼吸道的分泌物中还含有免疫球蛋白和其它物质,有助于防止感染和维持粘膜的完整性。
呼吸道受到机械或化学刺激时,可以引起防御反射,将在呼吸调节的一节中叙述。
二、肺通气原理
气体进入肺取决于两方面因素的相互作用:一是推动气体流动的动力;一是阻止其流动的阻力。前者必须克服后者,方能实现肺通气,正如心室射血的动力必须克服循环系统的阻力才能推动血液流动一样。
(一)肺通气的动力
气体进出肺是由大气和肺泡气之间存在着压力差的缘故。在自然呼吸条件下,此压力差产生于肺的张缩所引起的肺容积的变化。可是肺本身不具有主动张缩的能力,它的张缩是由胸廓的扩大和缩小所引起,而胸廓的扩大和缩小又是由呼吸肌的收缩和舒张所引起。当吸气肌收缩时,胸廓扩大,肺随之扩张,肺容积增大,肺内压暂时下降并低于大气压,空气就顺此压差而进入肺,造成吸气(inspiration)。反之,当吸气肌舒张和(或)呼气肌收缩时,胸廓缩小,肺也随之缩小,肺容积减小,肺内压暂时升高并高于大气压,肺内气便顺此压差流出肺,造成呼气(expiration)。呼吸肌收缩、舒张所造成的胸廓的扩大和缩小,称为呼吸运动。呼吸运动是肺通气的原动力。
1.呼吸运动 引起呼吸运动的肌为呼吸肌。使胸廓扩大产生吸气动作的肌肉为吸气肌,主要有膈肌和肋间外肌;使胸廓缩小产生呼气动作的是呼气肌,主要有肋间内肌和腹壁肌。此外,还有一些辅助呼吸肌,如斜角肌、胸锁乳突肌和胸背部的其它肌肉等,这些肌肉只在用力呼吸时才参与呼吸运动。
(1)吸气运动:只有在吸气肌收缩时,才会发生吸气运动,所以吸气总是主动过程。膈形状似钟罩,静止时向上隆起,位于胸腔和腹腔之间,构成胸腔的底。膈肌收缩时,隆起的中心下移,从而增大了胸腔的上下径,胸腔和肺容积增大,产生吸气。膈下移的距离视其收缩强度而异,平静吸气时,下移约1-2cm,深吸气时,下移可达7-10cm。由于胸廓呈圆锥形,其横截面积上部较小,下部明显加大。因此,膈稍稍下降就可使胸腔容积大大增加。据估计,平静呼吸时因膈肌收缩而增加的胸腔容积相当于总通气量的4/5,所以膈肌的舒缩在肺通气中起重要作用。膈肌收缩而膈下移时,腹腔内的器官因受压迫而使腹壁突出,膈肌舒张是时,腹腔内脏恢复原们画面为膈肌舒缩引起的呼吸运动伴以腹壁的起伏,所以这种型式的呼吸称为腹式呼吸(abdominal breathing)。
肋间外肌的肌纤维起自上一肋骨的近脊椎端的下缘,斜向前下方走行,止于下一肋骨近胸骨端的止缘。由于脊椎的位置是固定的,而胸骨可以上下移动,所以当肋间外肌收缩时,肋骨和胸骨都向上提,肋骨下缘还向外侧偏转,从而增大了胸腔的前后径和左右径,产生吸气。肋间外肌收缩越强,胸腔容积增大越多。在平静呼吸中肋间外肌所起的作用较膈肌为小。由肋间肌舒缩使肋骨和胸骨运动所产生的呼吸运动,称为胸式呼吸(thoracic breathing)。腹式呼吸和胸式呼吸常同时存在,其中某种型式可占优势;只有在胸部或腹部活动受到限制时,才可能单独出现某一种型式的呼吸。
(2)呼气运动:平静呼气时,呼气运动不是由呼气肌收缩所引起,而是因膈股长肋间外肌舒张,肺依靠本身的回缩力量而回位,并牵引胸廓缩小,恢复其吸气开始前的位置,产生呼气。所以平静呼吸时,呼气是被动的。用力呼吸时,呼气肌才参与收缩,使胸廓进一步缩小,呼气也有了主动的成分。肋间内肌走行方向与肋间外肌相反,收缩时使肋骨和胸骨下移,肋骨还向内侧旋转,使胸腔前后、左右缩小,产生呼气。腹壁肌的收缩,一方面压迫腹腔器官,推动膈上移,另一方面也牵拉下部的肋骨向下向内移位,两者都使胸腔容积缩小,协助产生呼气。
(3)平静呼吸和用力呼吸:安静状态下的呼吸称为平静(平和)呼吸(eupnea)。其特点是呼吸运动较为平衡均匀,每分钟呼吸频率约12-18次,吸气是主动的,呼气是被动的。机体活动时,或吸入气中的二氧化碳含量增加或氧含量减少时,呼吸将加深、加快,成为深呼吸或用力呼吸,这时不仅有更多的吸气肌参与收缩,收缩加强,而且呼气肌也主动参与收缩。在缺氧或二氧化碳增多较严重的情况下,会出现呼吸困难(dyspnea),这时,不仅呼吸大大加深,而且出现鼻翼扇动等,同时主观上有不舒服的困压感。
2.肺内压 肺内压是指肺泡内的压力。在呼吸暂停、声带开放、呼吸道畅通时,肺内压与大气压相等。吸气之初,肺容积增大,肺内压暂进下降,代于大气压,空气在此压差推动下进入肺泡,随着肺内气体逐渐增加,肺内压也逐渐升高,至吸气末,肺内压已升高到和大气压相等,气流也就停止(图5-2)。反之,在呼气之初,肺容积减小,肺内压暂时升高并超过大气压,肺内气体便流出肺,使肺内气体逐渐减少,肺内压逐渐下降,至呼气末,肺内压又降到和大气压相等。
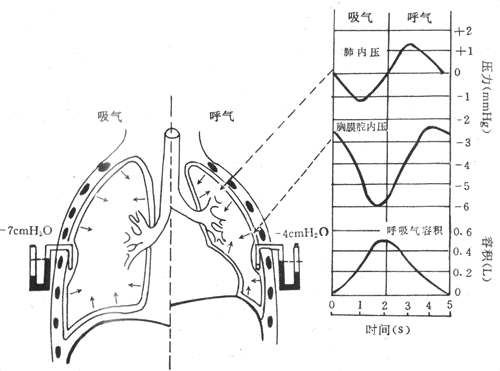
图5-2 吸气和呼气时,肺内压、胸膜腔内压及呼吸气容积的变化过程(右)和胸膜腔内压直接测量示意图(左)
(1mmHg=0.133kPa,1cmH20g0.098kPa)
呼吸过程中肺内压变化的程度,视呼吸的缓急、深浅和呼吸道是否通畅而定。若呼吸慢,呼吸道通畅,则肺内压变化较小;若呼吸较快,呼吸道不够通畅,则肺内压变化较大。平静呼吸时,呼吸缓和,肺容积的变化也较小,吸气时,肺内压较大气压约低0.133-0.266kPa(1-2mmHg),即肺内压为-0.266—-0.133kPa(-2—-1mmHg);呼气时较大气压约高0.133-0.266kPa(1-2mmHg)。用力呼吸时,呼吸深快,肺内压变化的程度增大。当呼吸道不够通畅时,肺内压的升降将更大。例如紧闭声门,尽力作呼吸动作,吸气时,肺内压可为-13.3——-3.99kPa(-100—-30mmHg),呼气时可达7.89-18.62kPa(60-140mmHg)。
由此可见,在呼吸过程中正是由于肺内压的周期性交替升降,造成肺内压和大气压之间的压力差,这一压力差成为推动气体进出肺的直接动力。一旦呼吸停止,便可根据这一原理,用人为的方法造成肺内压和大气压之间的压力差来维持肺通气,这便是人工呼吸。人工呼吸的方法很多,如用人工呼吸机进入正压通气;简便易行的口对口的人工呼吸;节律地举臂压背或挤压胸廓等。但在旅行人工呼吸时,首先要保持呼吸道畅通,否则,对肺通气而言,操作将是无效的。
3.胸膜腔和胸膜腔内压 如上所述,在呼吸运动过程中随胸腔廓的运动而运动。肺为何能随胸廓而运动呢?这是因为在肺和胸廓之间存在一密闭的胸膜腔和肺本身有可扩张性的缘故。胸膜有两层,即紧贴于肺表面的脏层和紧贴于胸廓内壁的壁层。两层胸膜形成一个密闭的潜在的腔隙,为胸膜腔。胸膜腔内仅有少量浆液,没有气体,这一薄层浆液有两方面的作用。一是在两层胸膜之间起润滑作用。因为浆液的粘滞性很低,所以在呼吸运动过程中,两层胸膜可以互相滑动,减小磨擦。二是浆液分子的内聚力使两层胸膜贴附在一起,不易分开,所以肺就可以随胸廓的运动而运动。因此,胸膜腔的密闭性和两层胸膜间浆液分子的内聚力有重要的生理意义。如果胸膜腔破裂,与大气相通,空气将立即进入胸膜腔,形成气胸,两层胸膜彼此分开,肺将因其本身的回缩力而塌陷。这时,尽管呼吸运动仍在进行,肺却减小或失去了随胸廓运动而运动的能力,其程度视气胸的程度和类型而异。显然,气胸时,肺的通气功能受到妨害,严重者,应紧急处理。
胸膜腔内的压力为胸膜腔内压(intrapleural pressure),可用两种方法进行测定。一种是直接法,将与检压计相连接的注射针头斜刺入胸膜腔内,检压计的液而即可直接指示胸膜腔内的压力(图5-2左)。直接法的缺点是有刺破胸膜脏层和肺的危险。另一种方法是间接法,让受试者吞下带有薄壁气囊的导管至下胸部的食管,由测量呼吸过程中食管内压变化来间接地指示胸膜腔内压变化。这是因为食管在胸内介于肺和胸壁之间,食管壁薄而软,在呼吸过程中两者的变化值基本一致。故可以测食管内压力的变化以间接反映胸膜腔内压的变化。
测量表明胸膜腔内压比大气压低,为负压。平静呼气末胸膜腔内压约为-0.665—-0.399kPa(-5—-3mmHg),吸气末约为-1.33—-0.665kPa(-10—-5mmHg)(图5-2)。关闭声门,用力吸气,胸膜腔内压可降至-11.97kPa(-90mmHg),用力呼气时,可升高到14.63kPa(110mmHg)。胸膜腔内负压不便作用于肺,牵引其扩张,也作用于胸腔内其它器官,特别是壁薄而可扩张性大的腔静脉和胸导管等,影响静脉血和淋巴液的回流。
胸膜腔内压为何是负压?从分析作用于胸膜腔的力来说明。有两种力通过胸膜脏层作用于胸膜腔:一是肺内压,使肺泡扩张;一是肺的弹性回缩力,使肺泡缩小(图5-2左,箭头所示)。因此,胸膜腔内的压力实际上是这两种方向相反的力的代数和,即:
胸膜爱内压=肺内压-肺弹性回缩力
在吸气末和呼气末,肺内压等于大气压,因而
胸膜腔内压=大气压-肺弹性回缩力
若以1个大气压为0位标准,则
胸膜腔内压= -肺弹性回缩力
如果肺弹性回缩力是0.665kPa(5mmHg),胸膜腔内压就是-0.665kPa(-5mmHg),实际的压力值便是101.08kPa—0.665kPa=100.415kPa(760mmHg—5mmHg=755mmHg)。可见,胸膜腔负压是由肺的弹性回缩力(其来源见下文,肺的弹性阻力和顺应性)造成的。吸气时,肺扩张,肺的弹性回缩力增大,胸膜腔负压也更负。呼气时,肺缩小,肺弹性回缩力也减小,胸膜腔负压也减少。但是,为什么在呼气末胸膜腔内压仍然为负?这是因为胎儿出生后,胸廓生长的速度比肺快,以致胸廓经常牵引着肺,即便在胸廓因呼气而缩小时,仍使肺处于一定程度的扩张状态,只是扩张程度小些而已。所以,正常情况下,肺总是表现出回缩倾向,胸膜腔内压因而经常为负。
综上所述,可将肺通气的动力概括如下:呼吸肌的舒缩是肺通气的原动力,它引起胸廓的张缩,由于胸膜腔和肺的结构功能特征,肺便随胸廓的张缩而张缩,肺容积的这种变化又造成肺内压和大气压之间的压力差,此压力差直接推动气体进出肺。
(二)肺通气的阻力
肺通气的动力需要克服肺通气的阻力方能实现肺通气。阻力增高是临床上肺通0气障碍最常见的原因。肺通气的阻力有两种:弹性阻力(肺和胸廓的弹性阻力),是平静呼吸时主要阻力,约占总阻力的70%;非弹性阻力,包括气道阻力,惯性阻力和组织的粘滞阻力,约占总阻力的30%,其中又以气道阻力为主。
1.用同等大小的外力作用时,弹性阻力大者,变形程度小;弹性阻力小者,变形程度大。一般用顺应性(compliance)来度量弹性阻力。顺应性是指在外力作用下弹性组织的可扩张性,容易扩张者顺应性大,弹性阻力小;不易扩张者,顺应性小,弹性阻力大。可见顺应性(C)与弹性阻力(R)成反变关系:
C=1/R
顺应性用单位压力变化(△P)所引起的容积变化(△V)来表示,单位是L/cmH2O,即 C=[(△V/△P)L ]/[cmH2O ]。
(1)肺弹性阻力和肺顺应性:肺具有弹性,在肺扩张变形时所产生的弹性回缩力,其方向与肺扩张的方向相反,因为是吸气的阻力,即肺弹性回缩力是肺的弹性阻力。肺的弹性阻力可用肺顺应性表示:

跨肺压是肺内压与胸膜腔内压之差。
1)肺静态顺应性曲线:测定肺顺应性时,进行分步吸气(或打气入肺)或分步呼气(或从肺内抽气),每步吸气或呼气后,屏气,放松呼吸肌,测定肺容积的变化和胸膜腔内压(因为这时呼吸道内没有气流流动,肺内压等于大气压,所以只测胸膜腔内压就可知道跨肺压)。然后绘制容积-压力(V-P)曲线(图5-3),就是肺的顺应性曲线。因为测定是在屏气无气流的情况下进行的,所在是肺静态顺应性。曲线的斜率反映不同肺容量下顺应性或弹性阻力的大小。曲线斜率大,顺应性大,弹性阻力小;曲线斜率小,则意义相反。正常成年人在平静呼气末,肺容积约为肺总量的40%左右时,肺顺应性正好位于曲线的中段,此段斜率最大,故平静呼吸时肺弹性阻力小,呼吸省力。此时健康成年人CL约为0.2L/cmH2O。当肺充血、肺组织纤维化化或肺泡表面活性物质(见后文)减少时,肺的弹性阻力增加,患者吸气困难;肺气肿时,肺弹性成分大量破坏,肺回缩量减小,弹性阻力减小,患者呼气困难。
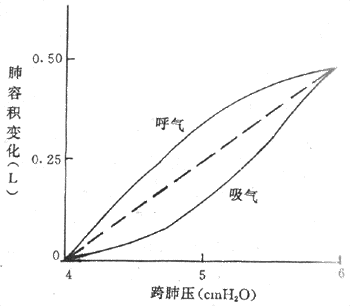
图5-3 肺的静态顺应性曲线(1cmH2O=0.098kPa)
2)比顺应性:肺顺应性还受肺总量的影响。肺总量大的,其顺应性较大;反之,较小。现举例说明。设若用0.5kPa(5cmH2O)的压力将1L的气体注入一个人的两肺,计算得出全肺顺应性为0.2L/cmH2O。如果左、右两肺的容积和顺应性是一样的,那么同样0.5kPa的压力,将同样1L气体送入肺内,每侧肺容量仅增加0.5L,计算出每侧肺的顺应性只有 0.1L/cmH2O,而不是0.2L/cmH2O。这是因为吸入同等容积的气体,在肺总量较大者,其扩张程度较小,弹性回缩也也较小,弹性阻力小,仅需较小的跨肺压变化即可,顺应性大;而在肺总量较小者,其扩张程度大,弹性加缩力也大,弹性阻力大,需较大的跨肺压变化,故顺应性小。由于不同个体间肺总量存在着差别,在比较其顺应性时必须排除肺总量的影响,进行标准化,测定单位肺容量下的顺应性,即比顺应性(specific compliance)。比顺应性=测得的肺顺应性(L/cmH2O)/肺总量(L)。
3)肺弹性阻力的来源:肺弹性阻力来自肺组织本身的弹性加回缩力和肺泡内侧的液体层同肺泡内气体之间的液-气界面的表面张力所产生的回缩力,两者均使肺具有回缩倾向,故成为肺扩张的弹性阻力。
肺组织的弹性阻力主要来自弹力和胶原纤维,当肺扩张时,这些纤维被牵拉便倾向于回缩。肺扩张越大,对纤维的牵拉程度也越大,回缩力也越大,弹性阻力也越大,反之则小。
图5-4示离体的肺在充气和充生理盐水时各自的顺应性曲线。可见扩张充气的肺比扩张充生理盐水的肺所需的跨肺压力大得多,前者约为后者的3倍。这是因为充气时,在肺泡内衬液和肺泡气之间存在液-气界面,从而产生表面张力。球形液-气界面的表面张力方向是向中心的,倾向于使肺泡缩小,产生弹性阻力。而充生理盐水时,没有液-气界面,因此不存在表面张力作用,仅肺组织的弹性回缩所产生的阻力作用。由此可见,肺组织的弹性阻力仅约占肺总弹性阻力的1/3,而表面张力的约占2/3。因此,表面张力对肺的张缩有重要的作用。
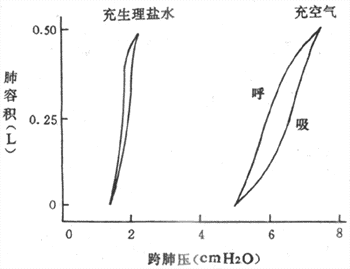
图5-4 充空气和充生理盐水时肺的顺应性曲线(1cmH2O=0.098kPa)
根据Laplace定律,P=2T/r(P是肺内的压力,T是肺泡表面张力,r是肺泡半径)。如果大、小肺泡的表面张力量样的,那么,肺泡内压力将随肺泡半径的大小而反变。小的肺泡,压力大;小的肺泡,压力小。如果这些肺泡彼此连通,结果小肺泡内的气体将流入大肺泡,小肺泡塌陷,大肺泡膨胀,肺泡将失去稳定性(图5-5)。但实际并未发生这种情况,这是因为肺泡存在着降低表面张力作用的表面活性物质的缘故。
肺泡表面活性物质(alveolar surfactant)是复杂的脂蛋白混合物,主要成分是二棕榈酰卵磷脂(dipalmitoyl lecithin,DPL或 dipalmitoylphosphatidyl choline choline,DPPC),由肺泡Ⅱ型细胞合成并释放,分子的一端是非极性疏水的脂肪酸,不溶于水,另一端是极性的易溶于水。因此,DPPC分子垂直排列于液-气界面,极性端插入水中,非极性端伸入肺泡气中,形成单分子层分布在液-气界面上,并随肺泡的张缩而改变其密度。正常肺泡表面活性物质不断更新,以保持其正常的功能。
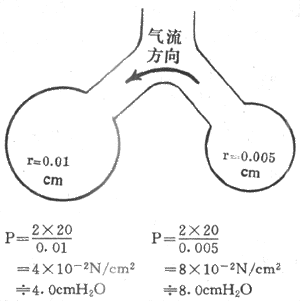
图5-5 相联通的大小不同的液泡内压及气流方向示意图1cmH2O=0.098kPa
肺泡表面活性物质降低表面张力的作用,有重要的生理功能。表面活性物质使肺泡液-气界面的表面张力降至10-4N/cm以下,比血浆的5×10-4N/cm低得多,这样,就减弱了表面张力对肺毛细血管中液体的吸引作用,防止了液体渗入肺泡,使肺泡得以保持相对干燥。此外,由于肺泡表面活性物质的密度大,降低表面张力的作用强,表面张力小,使小肺泡内压力不致过高,防止了小肺泡的塌陷;大肺泡表面张力则因表面活性物质分子的稀疏而不致明显下降,维持了肺内压力与小肺泡相等,不致过度膨胀,这样就保持了大小肺泡的稳定性,有利于吸入气在肺内得到较为均匀的分布。
成年人患肺炎、肺血栓等疾病时,可因表面活性物质减少而发生肺不张。初生儿也可因缺乏表面活性物质,发生肺不张和肺泡内表面透明质膜形成,造成呼吸窘迫综合症,导致死亡。现在已可应用抽取羊水并检查其表面活性物质含量的方法,协助判断发生这种疾病的可能性,采取措施,加以预防。例如,如果含量缺乏,则可延长妊娠时间或用药物(糖皮质类固醇)促进其合成。因此,了解肺泡Ⅱ型细胞的成熟过程及其表面活性物质的代谢和调节有重要的理论和实际意义。
(2)胸廓的弹性阻力和顺应性:胸廓也具有弹性,呼吸运动时也产生弹性阻力。但是,因胸廓弹性阻力增大而使肺通气发生障碍的情况较为少见,所以临床意义相对较小。胸廓处于自然位置时的肺容量,相当于肺总是的67%左右,此时胸廓毫元变化,不表现有弹性回缩力。肺容量小于总量的67%,胸廓被牵引向内而缩小,胸廓的弹性回缩力向外,是吸气的动力,呼气的弹性阻力;肺容量大于肺总量的67%时,胸廓被牵引向外而扩大,其弹性回缩力向内,成为吸气的弹性阻力,呼气的动力。所以胸廓的弹性回缩力既可能是吸气的弹性阻力,也可能是吸气的动力,视胸廓的位置而定,这与肺的不同,肺的弹性回缩力总是吸气的弹性阻力。
![]()
跨壁压为胸膜腔内压与胸壁外大气压之差。正常人胸廓顺应性也是0.2L/cmH20。胸廓顺应性可因肥胖、胸廓畸形、胸膜增厚和腹内占位病变等而降低。
因为肺和胸廓的弹性阻力呈串联排列,所以肺和胸廓的总弹性阻力是两者弹性阻力之和,如以顺应性来表示,即:
![]()
所以总顺应性为 0.1L/cmH20。
2. 非弹性阻力 非弹性阻力包括惯性阻力、粘滞阻力和气道阻力。惯性阻力是气流在发动、变速、换向时因气流和组织的惯性所产生的阻止运动的因素。平静呼吸时,呼吸频率低、气流流速慢,惯性阻力小,可忽略不计。粘滞阻力来自呼吸时组织相对位称所发生的磨擦。气道阻力来自气体流经呼吸道时气体分子间和气体分子与气道之间的磨擦,是非弹性阻力的主要成分,约占80%-90%。非弹性阻力是气体流动时产生的,并随流速加快而增加,故为动态阻力。
气道阻力(airway resistance)可用维持单位时间内气体流量所需压力差来表示:
![]()
健康人,平静呼吸时的总气道阻力为1-3cmH20/L·S-1,主要发生在鼻(约占总阻力50%),声门(约占25%)及气管和支气管(约占15%)等部位,仅10%的阻力发生在口径小于2mm的细支气管。
气道阻力受气流流速、气流形式和管径大小影响。流速快,阻力大;流速慢,阻力小。气流形式有层流和湍流,层流阻力小,湍流阻力大。气流太快和管道不规则容易发生湍流。如气管内有粘液、渗出物或肿瘤、异物等时,可用排痰、清除异物、减轻粘膜肿胀等方法减少湍流,降低阻力。气道管径大小是影响气道阻力的另一重要因素。管径缩小,阻力大增,因为R∝1/r4(见循环系统)。气道管径又受四方面因素折影响:
(1)跨壁压:这里跨壁压是指呼吸道内外的压力差。呼吸道内压力高,跨壁压增大,管径被动扩大,阻力变小;反之则增大。
(2)肺实质对气道壁的外向放射状牵引:小气道的弹力纤维和胶然后纤维与肺泡壁的纤维彼此穿插,这些纤维象帐蓬的拉线一样对气道发挥索引作用,以保持那些没有软骨支持的细支气管的通畅。
(3)自主神经系统对气道管壁平滑肌舒缩活动的调节:呼吸道平滑肌受交感、副交感双重神经支配,两者均有紧张性。副交感神经使气道平滑肌收缩,管径变小,阻力增加;交感神经使平滑肌舒张,管径变大,阻力降低,临床上常用拟上腺素能药物解除支气管痉挛,缓解呼吸困难,近来发现呼吸道平滑肌的舒缩还受自主神经释放的非乙酰胆碱的共存递质的调制,如神经肽(血管活性肠肽、神经肽Y、速激肽等)。它们或作用于接头前受体,调制递质的释放、或作用于接头后,调制对递质的反应或直接改变效应器的反应。
(4)化学因素的影响:儿茶酚胺可使气道平滑肌舒张;前列腺素F2α可使之收缩,而E2使之舒张;过敏反应时由肥大细胞释放的组胺和慢反应物质使支气管收缩;吸入气CO2含理的增加可以刺激支气管、肺的C类纤维,反射性地使支气管收缩,气道阻力增加。近来的研究发现气道上皮可合成、释放内皮素,使气道平滑肌收缩。哮喘病人肺内皮素的合成和释放增加,提示内皮素可能参与哮喘的病理生理过程。
在上述四种因素中,前三种均随呼吸而发生周期性变化,气道阻力也因而出现周期性改变。跨壁压增大(因胸膜内压下降),交感神经兴奋都能使气道口径增大,阻力减小;呼气时发生相反的变化,使气道口径变小,阻力增大,这也是为何支气管哮喘病人呼气比吸气更为困难的主要原因。
(三)呼吸功
在呼吸过程中,呼吸肌为克服弹性阻力和右面弹性阻力而实现肺通气所作的功为呼吸功。通常以单位时间内压力变化乘以容积变化来计算,单位是kg·m。正常人平静呼吸时,呼吸功不大,每分钟约为0.3-0.6kg·m,其中2/3用来克服弹性阻力,1/3用来克服非弹性阻力。劳动或运动时,呼吸频率、深度增加,呼气也有主动成分的参与,呼吸功可增至10kg·m。病理情况下,弹性或非弹性阻力增大时,也可使呼吸功增大。
平静呼吸时,呼吸耗能仅占全身耗能的3%。剧烈运动时,呼吸耗能可升高25倍,但由于全身总耗能也增大15-20倍,所以呼吸耗能仍只占总耗能的3%-4%。
三、基本肺容积和肺容量
了解肺通气量的简单方法是用肺量计记录进出肺的气量。图5-6示呼吸时肺容量变化的曲张。
(一)基本肺容积
图5-6左侧示肺的四种基本容积,它们互不重叠,全部相加等于肺的最大容量。
1.潮气量 每次呼吸时吸入或呼出的气量为潮气量(tidal volume,TV)。平静呼吸时,潮气量为400-600ml,一般以500ml 计算。运动时,潮气量将增大。
2.补吸气量或吸气贮备 平静吸气末,再尽力吸气所能吸入的气量为补吸气量(inspiratory reserve volume,IRV),正常成年人约为1500-200ml。
3.补呼气量或呼气贮备量 平静呼气末,再尽力呼气所能呼出的气量为补呼气量(espiratory reserve volume,ERV),正常成年人约为900-1200ml。
4.余气量或残气量 最大呼气末尚存留于肺中不能再呼出的气量为余气量(res idual volume,RV)。只能用间接方法测定,正常成人约为1000-1500ml。支气管哮喘和肺气肿患者,余气量增加。目前认为余气量是由于最大呼气之末,细支气管,特别是呼吸性细支气管关闭所致。
(二)肺容量
是基本肺容积中两项或两项以上的联合气量(图5-5右)。
1.深吸气量 从平静呼气末作最大吸气时所能吸入的气量为深吸气量(inspiratory capacity),它也是潮气量和补吸气量之和,是衡量最大通气潜力的一个重要指示。胸廓、胸膜、肺组织和呼吸肌等的病变,可使深吸气量减少而降低最大通气潜力。
2.功能余气量 平静呼气末尚存留于肺内的气量为功能余气量(functional residual capacity,FRC),是余气量和补呼气量之和。正常成年人约为2500ml,肺气肿患者的功能余气量增加,肺实质性病变时减小。功能余气量的生理意义是缓冲呼吸过程中肺泡气氧和二氧化碳分压(PO2和PCO2)的过度变化。由于功能余气量的稀释作用,吸气时,肺内PO2不至突然升得太高,PCO2不致降得太低;呼气时,肺内PO2则不会降得太低,PCO2不致升得太高。这样,肺泡气和动脉血液的PO2和PCO2就不会随呼吸而发生大幅度的波动,以处于气体交换。
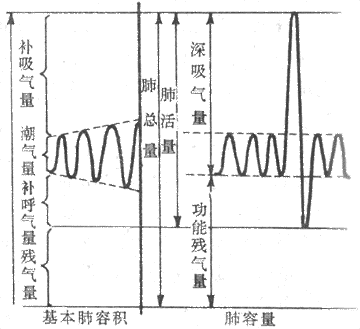
图5-6 基本肺容积和肺容量图解
3.肺活量和时间肺活量 最大吸气后,从肺内所能呼出的最大气量称作肺活(vital capacity,VC),是潮气量、补吸气量和补呼气量之和。肺活量有较大的个体差异,与身材大小、性别、年龄、呼吸肌强弱等有关。正常成年男性平均约为3500ml,女性为2500ml。
肺活是反映了肺一次通气的最大能力,在一定程度上可作为肺通气功能的指标。但由于测定肺活量时不限制呼气的时间,所以不能充分反映肺组织的弹性状态和气道的通畅程度,即通气功能的好坏。例如,某些病人肺组织弹性降低或呼吸道狭窄,通气功能已经受到损害,但是如果延长呼气时间,所测得的肺活量是正常的。因此,提出时间肺活量(timed vital capacity),也称用力呼气量的概念,用来反映一定时间内所能呼出的气量。时间肺活量为单位时间内呼出的气量占肺活量的百分数。测定时,让受试者先作一次深吸气,然后以最快的速度呼出气体,同时分别测量第1、2、3s末呼出的气量,计算其所占肺活量的百分数,分别称为第1、2、3s的时间肺活量(图5-7),正常人各为83%、96%和99%肺活量。时间肺活量是一种动态指标,不仅反映肺活量容量的大小,而且反映了呼吸所遇阻力的变化,所以是评论肺通气功能的较好指标。阻塞性肺疾病患者往往需要5-6秒或更长的时间才能呼出全部肺活量。
图5-7 时间肺活量
A:正常时间肺活量 B:气道狭窄时的时间肺活量
4.肺总量 肺所能容纳的最大气量为肺总量(total lung capacity,TLC),是肺活量和余气量之和。其值因性别、年龄、身材、运动锻炼情况和体位而异。成年男性平均为5000ml,女性3500ml。
四、肺通气量
(一)每分通气量
每分通气量(minute ventilation volume)是指每分钟进或出肺的气体总量,等于呼吸频率乘潮气量。平静呼吸时,正常成年人呼吸频率每分12-18次,潮气量500ml,则每分通气量6-9L。每分通气量随性别、年龄、身材和活动量不同而有差异。为便于比较,最好在基础条件下测定,并以每平方米体表面积为单位来计算。
劳动和运动时,每分通气量增大。尽力作深快呼吸时,每分钟所能吸入或呼出的最大气量为最大通气量。它反映单位时间内充分发挥全部通气量,是估计一个人能进行多大运动量的生理指标之一。测定时,一般只测量10s或15s最深最快的呼出或吸入量,再换算成每分钟的,即为最大通气量。最大通气量一般可达70-120L。比较平静呼吸时的每分通气量和最大通气量,可以了解通气功能的贮备能力,通常用通气贮量百分比表示:
通气贮量百分比=[(最大通气量-每分平静通气量)/最大通气量 ]×100%
正常值等于或大于93%。
(二)无效腔和肺泡通气量
每次吸入的气体,一部分将留在从上呼吸道至呼吸性细支气管以前的呼吸道内,这部分气体均不参与肺泡与血液之间的气体交换,故称为解剖无效腔(anatomical dead space),其容积约为150ml。进入肺泡内的气体,也可因血流在肺内分布不均而未能都与血液进入气体交换,未能发生气体交换的这一部分肺泡容量称为肺泡无效腔。肺泡无效腔与解剖无效腔一起合称生理无效腔(physiollgical dead space)。健康人平卧时生理无效腔等于或接近于解剖无效腔。
由于无效腔的存在,每次吸入的新鲜空气不能都到达肺泡进入气体交换。因此,为了计算真正有效的气体交换,应以肺泡通气量为准。肺泡通气量(alveolar ventilation)是每分钟吸入肺泡的新鲜空气量,等于(潮气量-无效腔气量)×呼吸频率。如潮气量是500ml,无效腔气量是150ml,则每次呼吸仅使肺泡内气体更新1/7左右。潮气量和呼吸频率的变化,对肺通气和肺泡通气有不同的影响。在潮气量减半和呼吸频率加倍或潮气量加倍而呼吸频率减半时,肺通气量保持不变,但是肺泡通气量却发生明显的变化,如表5-1所示。故从气体交换而言,浅而快的呼吸是不利的。
表5-1 不同呼吸频率和潮气量时的肺通气量和肺泡通气量
| 呼吸频率 | 潮气量 | 肺通气量 | 肺泡通气量 |
| (次/Imin) | (ml) | (ml/min) | (ml/min) |
| 16 | 500 | 8000 | 5600 |
| 8 | 1000 | 8000 | 6800 |
| 32 | 250 | 8000 | 3200 |
高频通气 近年来,临床上在某些情况下(如配合支气管镜检查,治疗呼吸衰竭等)使用一种特殊形式的人工通气,即高频通气。这是一种频率很高,潮气量很低的人工通气,其频率可为每分钟60-100次或更高,潮气量小于解剖无效腔,但却可以保持有效的通气和换气,这似乎与上述浅快呼吸不利于气体交换的观点矛盾。目前,对于高频通气何以能维持有效的通气和换气还不太清楚,可能其通气原理与通常情况下的通气原理不尽相同,有人认为它和气体对流的加强及气体分子扩散的的加速有关。高频通气的临床应用和通气原理都有待进一步研究。
第二节 呼吸气体的交换
肺通气使肺泡不断更新,保持了肺泡气PO2、PCO2的相对稳定,这是气体交换得以顺利进行的前提。气体交换包括肺换气和组织换气,在这两处换气的原理一样。
一、气体交换原理
(一)气体的扩散
气体分子不停地进行着无定向的运动,其结果是气体分子从分压高处向分压低处发生净转移,这一过程称为气体扩散,于是各处气体分压趋于相等。机体内的气体交换就是以扩散方式进行的。单位时间内氧化扩散的容积为气体扩散速率(diffusion rate,D),它受下列因素的影响。
1.气体的分压差 在混合气体中,每种气体分子运动所产生的压力为各该气体的分压,它不受其它气体或其分压存在的影响,在温度恒定时,每一气体的分压只决定于它自身的浓度。混合气的总压力等于各气体分压之和。
气体分压可按下式计算:
气体分压=总压力×该气体的容积百分比
两个区域之间的分压差(△P)是气体扩散的动力,分压差大,扩散快。
2.气体的分子量和溶解度质量轻的气体扩散较快。在相同条件下,各气体扩散速率和各气体分子量(MW)的平方根成反比。溶解度(S)是单位分压下溶解于单位容积的溶液中的气体的量。一般以1个大气压,38℃时,100ml液体中溶解的气体的ml数来表示。溶解度与分子量平方根之比(S/***)为扩散系数(diffusion coefficient),取决于气体分子本身的特性。CO2的扩散系数是O2的20倍,主要是因为CO2在血浆中的溶解度(51.5)约为O2的(2.14)24倍的缘故,虽然CO2的分子量(44)略大于O2的(32)。
3.扩散面积和距离扩散面积越大,所扩散的分子总数也越大,所以气体扩散速率与扩散面积(A)成正比。分子扩散的距离越大,扩散经全程所需的时间越长,因此,扩散速率与扩散距离(d)成反比。
4.温度 扩散速率与温度(T)成正比。在人体,体温相对恒定,温度因素可忽略不计。综上所述,气体扩散速率与上述诸因素的关系是:
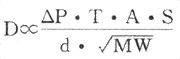
(二)呼吸气和人体不同部位气体的分压
既然气体交换的动力是分压差,则有必要首先了解进行气体交换各有关部位的气体组成和分压。
1.呼吸气和肺泡气的成分和分压 人体吸入的气体是空气。空气的主要成分是O2、CO2和N2,具有生理意义的是O2、和CO2。空气中各气体的容各百分比一般不因地域不同而异,但分压却因总大气压的变动而改变。高原大气压降低,各气体的分压也低。吸入的空气在呼吸道内被水蒸气所饱和,所在呼吸道内吸入气的成分已不同于大气,因此各成分的分压也发生相应的改变。
从肺内呼出的气体为呼出气,它来自两部分:无效腔的吸入气和来肺泡的肺泡气,是这两部分气体混合。
上述各部分气体的成分和分压如表5-2所示。
2.血液气体和组织气体的分压(张力)液体中的气体分压称为气体的张力(P),其数值与分压的相同。表5-3示血液和组织中的PO2和PCO2。不同组织的PO2和PCO2不同,同一组织的PO2和PCO2还受组织活动和水平的影响,表中值仅是安静状态下的大致估计值。
表5-2 海平面各气体的容积百分比ml%和分压kPa(mmHg)
| 大 气 容积百分比 |
分压 | 吸 入 气 容积百分比 |
分压 | 呼 出 气 容积百分比 |
分压 | 肺泡气 容积百分比 |
分压 | |
| O2 | 20.84 | 21.15 | 19.67 | 19.86 | 15.7 | 15.96 | 13.6 | 13.83 |
| (159.0) | (149.3) | (120.0) | (104.0) | |||||
| CO2 | 0.04 | 0.04 | 0.04 | 0.04 | 3.6 | 3.59 | 5.3 | 5.32 |
| (0.3) | (0.3) | (27.0) | (40.0) | |||||
| N2 | 78.62 | 79.40 | 74.09 | 74.93 | 74.5 | 75.28 | 74.9 | 75.68 |
| (597.0) | (563.4) | (566) | (569) | |||||
| H2O | 0.50 | 0.49 | 6.20 | 6.25 | 6.20 | 6.25 | 6.20 | 6.25 |
| (3.7) | (47) | (47) | (47) | |||||
| 合计 | 100.0 | 101.08 | 100.0 | 101.08 | 100 | 101.08 | 100 | 101.08 |
| (760) | (760) | (760) | (760) |
N2在呼吸过程中并无增减,只是因O2和CO2百分比的改变,使N2的百分比发生相对改变
表5-3 血液和组织中气体的分压kPa(mmHg)
| 动脉血 | 混合静脉血 | 组织 | |
| PO2 | 12.9-13.3 | 5.32 | 4 |
| (97-100) | (40) | (30) | |
| PCO2 | 5.32 | 6.12 | 6.65 |
| (40) | (46) | (50) |
二、气体在肺的交换
(一)交换过程
混合静脉血流经肺毛细血管时,血液PCO2是5.32kPa(40mmHg),比肺泡气的13.83kPa(104mmHg)低,肺泡气中O2便由于分压的差向血液扩散,血液的PCO2便逐渐上升,最后接近肺泡气的PCO2。CO2则向相反的方向扩散,从血液到肺泡,因为混合静脉血的PCO2是6.12kPa(46mmHg),肺泡的PCO2是5.32kPa(40mmHg)。(图5-8)。O2和CO2的扩散都极为迅速,仅需约0.3s即可达到平衡。通常情况下血液流经肺毛细血管的时间约0.7s,所以当血液流经肺毛细血管全长约1/3时,已经基本上完成交换过程(图5-9)。可见,通常情况下肺换气时间绰绰有余。
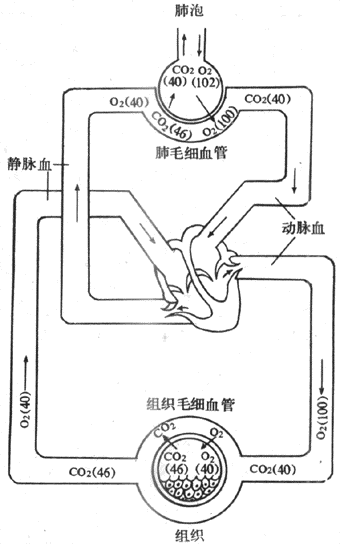
图5-8 气体交换示意图 数字为气体分压mmHg(1mmHg=0.133kPa)
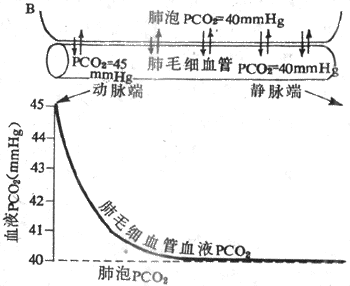
图5-9 肺毛细血管血液从肺泡摄取O2(A)和向肺泡排出CO2(B)的过程((1mmHg=0.133kPa))
(二)影响肺部气体交换的因素
前面已经提到气体扩散速率受分压差、扩散面积、扩散距离、温度和扩散系数的影响。这里只需具体说明肺的扩散距离和扩散面积以及影响肺部气体交换的其它因素,即通气/血流比值的影响。
1.呼吸膜的厚度 在肺部肺泡气通过呼吸膜(肺泡-毛细血管膜)与血液气体进行交换。气体扩散速率与呼吸膜厚度成反比关系,膜越厚,单位时间内交换的气体量就越少。呼吸膜由六层结构组成(图5-10);含表面活性物质的极薄的液体层、很薄的肺泡上皮细胞层、上皮基底膜、肺泡上皮和毛细血管膜之间很小的间隙、毛细血管的基膜和毛细血管内皮细胞层。虽然呼吸膜有六层结构,但却很薄,总厚度不到1μm,有的部位只有0.2μm,气体易于扩散通过。此外,因为呼吸膜的面积极大,肺毛细血管总血量不多,只60-140ml,这样少的血液分布于这样大的面积,所以血液层很薄。肺毛细血管平均直径不足8μm,因此,红细胞膜通常能接触至毛细血管壁,所以O2、CO2不必经过大量的血浆层就可到达红细胞或进入肺泡,扩散距离短,交换速度快。病理情况下,任何使呼吸膜增厚或扩散距离增加的疾病,都会降低扩散速率,减少扩散量,如肺纤维化、肺水肿等,可出现低氧血症;特别是运动时,由于血流加速,缩短了气体在肺部的交换时间,这时呼吸膜的厚度和扩散距离的改变显得更有重要性。
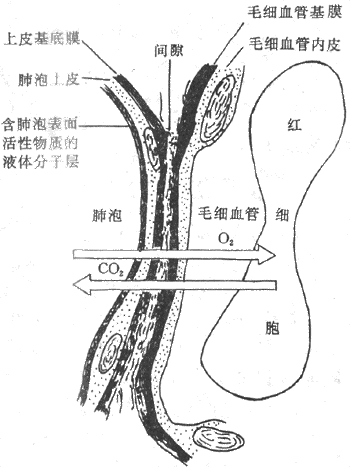
图5-10 呼吸膜结构示意图
2.呼吸膜的面积 气体扩散速率与扩散面积成正比。正常成人肺有3亿左右的肺泡,总扩散面积约70m2。安静状态下,呼吸膜的扩散面积约40m2,故有相当大的贮备面积。运动时,因肺毛细血管开放数量和开放程度的增加,扩散面积也大大增大。肺不张、肺实变、肺气肿或肺毛细血管关闭和阻塞均使呼吸膜扩散面积减小。
3.通气/血流比值的影响通气/血流比值(ventilation/perfusion ratio)是指每分肺通气量(VA)和每分肺血流量(Q)之间的比值(VA/Q),正常成年人安静时约为4.2/5=0.84。不难理解,只有适宜的VA/Q才能实现适宜的气体交换,这是因为肺部的气体交换依赖于两个泵协调工作。一个是气历史意义,使肺泡通气,肺泡气得以不断更新,提供O2,排出CO2;一个是血泵,向肺循环泵入相应的血流量,及时带起摄取的O2,带来机体产生的CO2。如果VA/Q比值增大,这就意味着通气过剩,血流不足,部分肺泡气未能与血液气充分交换,致使肺泡无效腔增大。反之,VA/Q下降,则意味着通气不足,血流过剩,部分血液流经通气不良的肺泡,混合静脉血中的气体未能得到充分更新,未能成为动脉血就流回了心脏。犹如发生了动-静脉短路,只不过是功能性的而不是解剖结构所造成的动-静脉短路。由此可见,VA/Q增大,肺泡无效腔增加;VA/Q减小,发生功能性动-静脉短路。两者都妨碍了有效的气体交换,可导致血液缺O2或CO2潴留,但主要是血液缺O2。这是因为,动、静脉血液之间O2分压差远远大于CO2的,所以动-静脉短路时,动脉血PO2下降的程度大于PCO2升高的程度;CO2的扩散系数是O2的20倍,所以CO2的扩散较O2为快,不易潴留;动脉血PO2下降和PCO2升高,可以刺激呼吸,增加肺泡通气量,有助于CO2的排出,却几乎无助于O2摄取,这是由氧离曲线和CO2解离曲线的特点所决定的(见第三节)。肺气肿病人,因许多细支气管阻塞和肺泡壁的破坏,上述两种VA/Q异常都可以存在,致使肺换气速率受到极大损害,这是造成肺换气功能异常最常见的一种疾病。
健康成人就整个肺而言VA/Q是0.84。但是肺内肺泡通气量和肺毛细血管血流量的分布不是很均匀的,因此,各个局部的通气/血流比值也不相同,用整个肺的VA/Q就不以反映出来。例如人友直立位时,由于重力等因素的作用,肺尖部的通气和血流都较肺底的小,不过血流量的减少更为显著,所以肺尖部的通气/血流比值增大,产生中度肺泡无效腔,而肺底的比值减小,产生功能性动-静脉短路(图5-11)。虽然正常情况下存在着肺泡通气和血流的不均匀分布,但从总体上说,由于呼吸膜面积远远超过气体交换的实际需要,所以并未明显影响O2的摄取和CO2的排出。
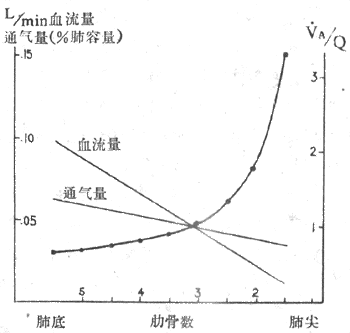
图5-11 正常人直立时肺通气和血流量的分布VA/Q,通气/血流比值
(三)肺扩散容量
气体在0.133kPa(1mmHg)分压差作用下,每分钟通过呼吸膜扩散的气体的ml数为肺扩散容量(pulmonary diffusion capacyty,DL),即:
DL=V/(PA-PC)
V是每分钟通过呼吸膜的气体容积(ml/min),PA是肺泡气中该气体的平均分压,Pc是肺毛细血管血液内该气体的平均分压。肺扩散容量是测定呼吸气通过呼吸膜的能力的一种指标。正常人安静时氧的肺扩散容量平均约为20ml/min·0.133kPa,CO2的为O2的20倍。运动时DL增加,是因为参与气体交换的肺泡膜面积和肺毛细血管血流量增加以及通气、血流的不均分布得到改善所致,DL可因有效扩散面积减小扩散距离增加而降低。
三、气体在组织的交换
气体在组织的交换机制、影响因素与肺泡处相似,所不同者是交换发生于液相(血液、组织液、细胞内液)之间,而且扩散膜两侧的O2和CO2的分压差随细胞内氧化代谢的强度和组织血流量而异血流量不变时,代谢强、耗O2多,则组织液CO2低,PCO2高;代谢率不变时,血流量大,则PO2高,PCO2低。
在组织处,由于细胞有氧代谢,O2被利用并产生CO2,所以PO2可低至3.99kPa(30mmHg)以下,PCO2可高达6.65kPa(30mmHg)以上.动脉血流经组织毛细血管时,便顺分压差由血液向细胞扩散,CO2则由细胞内血液扩散(图5-8),动脉血因失去O2和得到CO2而变成静脉血。
第三节 气体在血液中的运输
从肺泡扩散入血液的O2必须通过血液循环运送到各组织,从组织散入血液的CO2的也必须由血液循环运送到肺泡。下述O2和CO2在血液中运输的机制。
一、氧和二氧化碳在血液中存在的形式
O2和CO2的都以两种形式存在于血液:物理溶解的和化学结合的。
气体在溶液中溶解的量与分压和溶解度成正比,和温度成反比。温度38℃时,1个大气压(760Hg,101.08kPa)的 O2和 CO2和在100ml血液中溶解的量分别是2.36ml和48ml。按此计算,静脉血 PCO2和为6.12kPa(46mmHg),则每100ml血液含溶解的CO2为(48×6.12)/101.08=2.9ml;动脉血PO2为13.3kPa(100mmHg),每100ml血液含溶解的O2为(2.36×13.3)/101.08=0.31ml。可是,血液中实际的O2和O2为CO2含量比这数字大得多(表5-4),以溶解形式存在的O2、CO2比例极少,显然单靠溶解形式来运输O2、CO2不能适应机体代谢的需要。例如,安静状态下人体耗O2量约为250ml/min,如只靠物理溶解的O2来提供,则需大大提高心输出量或提高肺泡内的PO2,这对机体极其不利,所幸在进化过程中形成了O2、CO2为极为有效地化学结合的运输形式,大大减轻了对心脏和呼吸器官的苛求。
表5-4血液O2和CO2的含量(ml/100ml 血液)
| 物理溶解的 | 动脉血化学结合的 | 合计 | 物理溶解的 | 混合静脉血 化学结合的 |
合计 | |
| O2 | 0.31 | 20.0 | 20.31 | 0.11 | 15.2 | 15.31 |
| CO2 | 2.53 | 46.4 | 48.93 | 2.91 | 50.0 | 52.91 |
虽然溶解形式的O2、CO2很少,但也很重要。因为在肺或组织进行气体交换时,进入血液的O2、CO2都是先溶解,提高分压,再出现化学结合;O2、CO2从血液释放时,也是溶解的先逸出,分压下降,结合的再分离出现补充所失去的溶解的气体。溶解的和化学结合的两者之间处于动态平衡。
二、氧的运输
血液中的O2以溶解的和结合的两种形式存在。溶解的量极少,仅占血液总O2含量的约1.5%,结合的占 98.5%左右。O2的结合形式是氧合血红蛋白(HbO2)。血红蛋白(hemoglobin,Hb)是红细胞内的色蛋白,它的分子结构特征使之成为极好的运O2工具。Hb还参与CO2的运输,所以在血液气体运输方面Hb占极为重要的地位。
(一)Hb分子结构简介
每1Hb分子由1个珠蛋白和4个血红素(又称亚铁原卟啉)组成(图5-12)。每个血红素又由4个吡咯基组成一个环,中心为一铁原子。每个珠蛋白有4条多肽链,每条多肽链与1个血红至少连接构成Hb的单体或亚单位。Hb是由4个单体构成的四聚体。不同Hb分子的珠蛋白的多肽链的组成不同。成年人Hb(HbA)的多肽链是2条α链和2条β链,为α2β2结构。胎儿Hb(HbF)是2条α链和2条γ链,为α2γ2结构。出生后不久HbF即为HbFA所取代。多肽链中氨基酸的排列顺序已经清楚。每条α链含141个氨基酸残基,每条β链含146个氨在酸残基。血红素的Fe2+均连接在多肽链的组氨基酸残基上,这个组氨酸残基若被其它氨基酸取代,或其邻近的氨基酸有所改变,都会影响Hb的功能。可见蛋白质结构和功能密切相关。
Hb的4个单位之间和亚单位内部由盐键连接。Hb与O2的结合或解离将影响盐键的形成或断裂,使Hb四级结构的构型发生改变,Hb与O2的亲和力也随之而变,这是Hb氧离曲线呈S形和波尔效应的基础(见下文)。
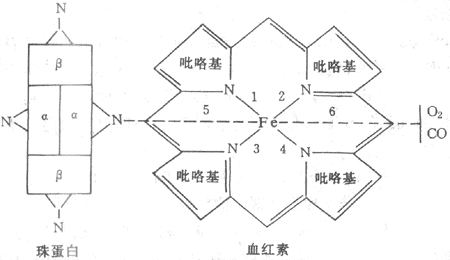
图5-12 血红蛋白组成示意图
(二)Hb与O2结合的特征
血液中的O2主要以氧合Hb(HbO2)形式运输。O2与Hb的结合有以下一些重要特征:
1.反应快、可逆、不需酶的催化、受PO2的影响。当血液流经PO2高的肺部时,Hb与 O2结合,形成HbO2;当血液流经PO2低的组织时,HbO2迅速解离,释放O2,成为脱氧Hb:
![]()
2.Fe2+与O2结合后仍是二价铁,所以该反应是氧合(oxygenation),不是氧化(oxidation)。
3.1分子Hb可以结合4分子O2。Hb分子量是64000-67000道尔顿(d),所以1gHb可以结合1.34-1.39mlO2,视Hb纯度而异。100ml血液中,Hb所能结合的最大O2量称为Hb的氧容量。此值受Hb浓度的影响;而实际结合的O2量称为Hb的氧含量,其值可受PO2的影响。Hb氧含量和氧容量的百分比为Hb氧饱和度。例如,Hb浓度在15g/100ml血液时,Hb的氧容量=15×1.34=Hb 20.1ml/100ml血液,如Hb的氧含量是20.1ml,则Hb氧饱和度是100%。如果Hb氧含量实际是15ml,则Hb氧饱和度=15/20×100%=75%。通常情况下,溶解的O2极少,故可忽略不计,因此,Hb氧容量,Hb氧含量和Hb氧饱和度可分别视为血氧容量(osygen capacity)、血氧含量(oxygen content)和血氧饱和度(oxygen saturatino)。HbO2呈鲜红色,去氧Hb呈紫蓝色,当体表表浅毛细血管床血液中去氧Hb 含量达5g/100ml血液以上时,皮肤、粘膜呈浅蓝色,称为紫绀。
4.Hb与O2的结合或解离曲线呈S形,与Hb的变构效应有关。当前认为Hb有两种构型:去氧Hb为紧密型(tense form,T型),氧合Hb为疏松型(relaxed form,R型)。当O2与Hb的Fe2+结合后,盐键逐步断裂,Hb 分子逐步由T型变为R型,对O2的亲和力逐步增加,R型的O2亲和力为T型的数百倍。也就是说,Hb 的4个亚单位无论在结合O2或释放O2时,彼此间有协同效应,即1个亚单位与O2结合后,由于变构效应的结果,其它亚单位更易与O2结合;反之,当HbO2的1个亚单位释出O2后,其它亚单位更易释放O2。因此,Hb氧离曲线呈S形。
(三)氧离曲线
氧离曲线(oxygen dissociation curve)或氧合血红蛋白解离曲线是表示PO2与Hb 氧结合量或Hb氧饱和度关系的曲线(图5-13)。该曲线既表示不同PO2时,O2与Hb 的结合情况。上面已经提到的曲线呈S形,是Hb变构效应所致。同时曲线的S形还有重要的生理意义,下面分析氧离曲线各段的特点及其功能意义。
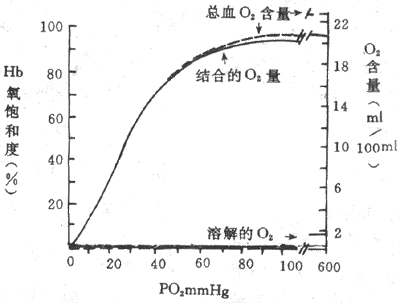
图5-13 氧离曲线
(实线,在Ph7.4,PCO240mmHg,温度37℃时测定的)
同时示溶解的O2和在Hb浓度为15g/100ml血液时的总血O2含量(1mmHg=0.133kPa)
1.氧离曲线的上段 相当于PO27.98-13.3kPa(60-100mmHg),即PO2较高的水平,可以认为是Hb与O2结合的部分。这段曲线较平坦,表明PO2的变化对Hb氧饱和度影响不大。例如PO2为13.3kPa(100mmHg)时(相当于动脉血PO2),Hb氧饱和度为97.4%,血O2含量约为19.4ml%;如将吸入气PO2提高到19.95kPa(150mmHg),Hb氧饱和度为100%,只增加了2.6% ,这就解释了为何VA/Q不匹配时,肺泡通气量的增加几乎无助于O2的摄取;反之,如使PO2下降到9.31kPa(70mmHg),Hb氧饱和度为94%,也不过只降低了3.4%。因此,即使吸入气或肺泡气PO2有所下降,如在高原、高空或某些呼吸系统疾病时,但只要PO2不低于7.98kPa(60mmHg),Hb氧饱和度仍能保持在90%以上,血液仍可携带足够量的O2,不致发生明显的低血氧症。
2.氧离曲线的中段 该段曲线较陡,相当于PO25.32-7.98kPa(40-60mmHg),是HbO2释放O2的部分。PO25.32kPa(40mmHg),相当于混合静脉血的PO2,此时Hb氧饱和度约为75%,血O2含量约14.4ml%,也即是每100ml血液流过组织时释放了5mlO2。血液流经组织液时释放出的O2容积所占动脉血O2含量的百分数称为O2的利用系数,安静时为25%左右。以心输出量5L计算,安静状态下人体每分耗O2量约为250ml。
3.氧离曲线的下段 相当于PO22-5,32kPa(15-40mmHg),也是H bO2与O2解离的部分,是曲线坡度最陡的一段,意即PO2稍降,HbO2就可大大下降。在组织活动加强时,PO2可降至2kPa(15mmHg),HbO2进一步解离,Hb氧饱和度降至更低的水平,血氧含量仅约4.4ml%,这样每100ml血液能供给组织15mlO2,O2的利用系数提高到75%,是安静时的3倍。可见该段曲线代表O2贮备。
(四)影响氧离曲线的因素
Hb与O2的结合和解离可受多种因素影响,使氧离曲线的位置偏移,亦即使Hb对O2的亲和力发生变化。通常用P50表示Hb对O2的亲和力。P50是使Hb氧饱和度达50%时的PO2,正常为3.52 kPa(26.5mmHg)。P50增大,表明Hb对O2的亲和力降低,需更高的PO2才能达到50%的Hb氧饱和度,曲线右移;P50降低,指示Hb对O2的亲和力增加,达50%Hb氧饱和度所需的PO2降低,曲线左移。影响Hb与O2亲和力或P50的因素有血液的Ph、PCO2、温度和有机磷化物(图5-14)。
1.Hb与PCO2的影响pH降低或升PCO2升高,Hb对O2的亲和力降低,P50增大,曲线右移; pH升高或PCO2降低,Hb对O2的亲和力增加,P50降低,曲线左移。酸度对Hb氧亲和力的这种影响称为波尔效应(Bohr effect)。波尔效应的机制,与 pH改变时h b构型变化有关。酸度增加时,H+与Hb多肽链某些氨基酸残基的基团结合,促进盐键形成,促使Hb分子构型变为T型,从而降低了对O2的亲和力,曲线右移;酸度降低时,则促使盐键断裂放出H+,Hb变为R型,对O2的亲和力增加,曲线左移。PCO2的影响,一方面是通过PCO2改变时,pH也改变间接效应,一方面也通过CO2与Hb结合而直接影响Hb与O2的亲和力,不过后一效应极小。
波尔效应有重要的生理意义,它既可促进肺毛细血管的氧合,又有利于组织毛细血管血液释放O2。当血液流经肺时,CO2从血液向肺泡扩散,血液PCO2下降,[H+]也降低,均使Hb对O2的亲和力增加,曲线左移,在任一PO2下Hb氧饱和度均增加,血液运O2量增加。当血液流经组织时,CO2从组织扩散进入血液,血液PCO2和[H+]升高,Hb对O2的亲和力降低,曲线右移,促使HbO2解离向组织释放更多的O2。
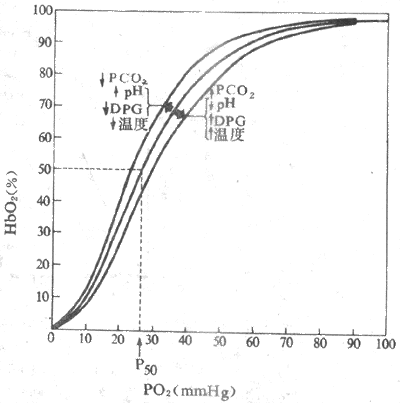
图5-14 影响氧离曲线位置的主要因素(1mmHg=0.133kPa)
2.温度的影响 温度升高,氧离曲线右移,促使O2释放;温度降低,曲线左移,不利于O2的释放。临床低温麻醉手术时应考虑到这一点。温度对氧离曲线的影响,可能与温度影响了H+活度有关。温度升高H+活度增加,降低了Hb对O2的亲和力。当组织代谢活跃是局部组织温度升高,CO2和酸性代谢产物增加,都有利于Hb02解离,活动组织可获得更多的O2以适应其代谢的需要。
3.2,3-二磷酸甘油酸 红细胞中含有很多有机磷化物,特别是2,3-二磷酸甘油酸(2.3-diphospoglyceric acid,2,3-DPG),在调节Hb和O2的亲和力中起重要作用。2,3-DPG浓度升高,Hb对O2亲和力降低,氧离曲线右移:2,3-DPG浓度升降低,Hb对O2的亲和力增加,曲线左移。其机制可能是2,3-DPG与Hbβ链形成盐键,促使Hb变成T型的缘故。此外,2,3-DPG可以提高[H+],由波尔效应来影响Hb对O2的亲和力。
2,3-DPG是红细胞无氧糖酵解的产物。高山缺O2,糖酵解加强,红细胞 2,3-DPG增加,氧离曲线右移,有利于O2的释放,曾认为这可能是能低O2适应的重要机制。可是,这时肺泡PO2也降低,红细胞内过多的2,3-DPG也妨碍了Hb与O2的结合。所以缺O2时,2,3-DPG使氧离曲线右移是否有利,是值得怀疑的。
4.Hb自身性质的影响除上述因素外,Hb与O2的结合还为其自身性质所影响。Hb的Fe2+氧化成Fe3+,失去运O2能力。胎儿Hb和O2的亲和力大,有助于胎儿血液流经胎盘时从母体摄取O2。异常Hb 也降低运O2功能。CO与Hb结合,占据了O2的结合位点,HbO2下降。CO与Hb的亲和力是O2的250倍,这意味着极低的PCO,CO就可以从HbO2中取代O2,阻断其结合位点。此外,CO还有一极为有害的效应,即当CO与Hb分子中某个血红素结合后,将增加其余3个血红素对O2的亲和力,使氧离曲线左移,妨碍O2的解离。所以CO中毒既妨碍Hb与O2的结合,又妨碍O2的解离,危害极大。
总之,血液Hb的运O2量可受多种因素影响:包括PO2、Hb本身的性质和含量、pH、PCO2、温度、2,3-DPG和CO等,pH降低,PCO2升高,温度升高,2,3-DPG增高,氧离曲线右移;pH升高,PCO2、温度、2,3-DPG降低和CO中毒,曲线左移。
三、二氧化碳的运输
(一)CO2的运输
血液中CO2也 以溶解和化学结合的两种形式运输。化学结合的CO2主要是碳酸氢盐和氨基甲酸血红蛋白。表5-5示血液中各种形式CO2的含量(ml/100ml 血液)、运输量(%)和释出量(%)。溶解的CO2约占总运输量的5%,结合的占95%(碳酸氢盐形式的占88%,氨基甲酸血红蛋白形式占7%)。
从组织扩散入血CO2首先溶解于血浆,一小部分溶解的CO2缓慢地和水结合生成碳酸,碳酸又解离成碳酸氢根和氢离子,H+被血浆缓冲系统缓冲,pH无明显变化。溶解的CO2也与血浆蛋白的游离氨基反应,生成打官司基甲酸血浆蛋白,但形成的量极少,而且动静脉中的含量相同,表明它对CO2的运输不起作用。
在血浆中溶解的CO2绝大部分扩散进入红细胞内,在红细胞内主要以下述结合形式存在:
表5-5 血液中各种形式CO2的含量(ml/100ml血液)、运输量(%)和释出量(%)
| 动脉血 | 静脉血 | 差值 | 释出题 | |||
| 含量 | 运输量 | 含量 | 运输量(动、静永血间) | |||
| CO2总量 | 48.5 | 100 | 52,5 | 100 | 4.0 | 100 |
| 溶解的CO2 | 2.5 | 5.15 | 2.8 | 5.33 | 0.3 | 7.5 |
| HCO3形式的CO2 | 43.0 | 88.66 | 46.0 | 87.62 | 3.0 | 75 |
| 氨基甲酸血红 蛋白的CO2 |
3.0 | 6.19 | 3.7 | 7.05 | 0.7 | 17.5 |
运输量(%)是指各种形式的CO2含量/CO2总含量×100%
释放量(%)是指各种形式的CO2在肺释放量/CO2总释放量×100%
1.碳酸氢盐 从组织扩散进入血液的大部分CO2,在红细胞内与水反应生成碳酸,碳酸又解离成碳酸氢根和氢离子,反应极为迅速,可逆(图5-15)。这是因为红细胞内含有较高浓度的碳酸酐酶,在其催化下,使反应加速5000倍,不到1s即达平衡。在此反应过程中红细胞内碳酸氢根浓度不断增加,碳酸氢根便顺浓度梯度红细胞膜扩散进入血浆。红细胞负离子的减少应伴有同等数量的正离子的向外扩散,才能维持电平衡。可是红细胞膜不允许正离子自由通过,小的负离子可以通过,于是,氯离子便由血浆扩散进入红细胞,这一现象称为氯离子转移(chloride shift)。在红细胞膜上有特异的HCO3—CI-载体,运载这两类离子跨膜交换。这样,碳酸氢根便不会在红细胞内堆积,有利于反应向右进行和CO2的运输。在红细胞内,碳酸氢根与K+结合,在血浆中则与Na+结合成碳酸氢盐。上述反应中产生的H+,大部分和Hb结合,Hb 是强有力的缓冲剂。
![]()
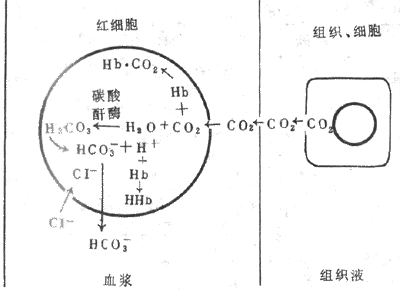
图5-15 CO2在血液中的运输示意图
在肺部,反应向相反方向(左)进行。因为肺泡气PCO2比静脉血的低,血浆中溶解的CO2首先扩散入肺泡,红细胞内的HCO3+H+生成H2CO3,碳酸酐酶又催化H2CO3分解成CO2和H2O,CO2又从红细胞扩散入血浆,而血浆中的HCO3便进入红细胞以补充消耗的HCO3,CI-则出红细胞。这样以HCO3形式运输的CO2,在肺部又转变成CO2释出。
2.氨基甲酸血红蛋白 一部分CO2与Hb的氨基结合生成氨基甲酸血红蛋白(carbaminohemoglobin),这一反应无需酶的催化、迅速、可逆,主要调节因素是氧合作用。
![]()
HbO2与CO2结合形成HbNHCOOH的能力比去氧Hb的小。在组织里,解离释出O2,部分HbO2变成去氧Hb,与CO2结合生成HbNHCOOH。此外,去氧Hb 酸性较HbO2弱,去氧Hb和H+结合,也促进反应向右侧进行,并缓冲了pH的变化。在肺的HbO2生成增多,促使HHbNHCOOH解离释放CO2和H+,反应向左进行。氧合作用的调节有重要意义,从表5-5可以看出,虽然以氨基甲酸血红蛋白形式运输的CO2仅占总运输量的7%,但在肺排出的CO2中却有17.5%是从氨基甲酸血红蛋白释放出来的。
(二)CO2解离曲线
CO2解离曲线(carbon dioxide dissociation curve)是表示血液中CO2含量与PCO2关系的曲线(图5-16)。与氧离曲线不同,血液CO2含量随PCO2上升而增加,几乎成线性关系而不是s 形,而且没有饱和点。因此,CO2解离曲线的纵坐标不用饱和度而用浓度来表示。
图5-16的A点是静脉血PO25.32kPa(40mmHg),PCO26kPa(45mmHg)时的CO2含量,约为52ml%;B点是动脉血PO213.3kPa(100mmHg),PCO25.32kPa(40mmHg)时的CO2含量,约为48ml%,血液流经肺时通常释出CO24ml/100ml血液。
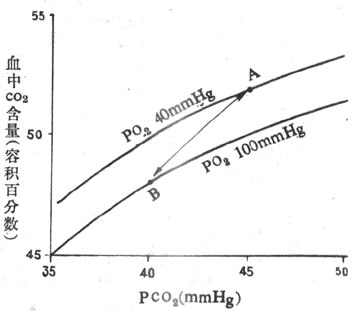
图5-16 CO2解离曲线
A:静脉血 B:动脉血(1mmHg=0.133kPa)
(三)氧与Hb的CO2运输的影响
O2与Hb结合将促使CO2释放,这一效应称作何尔登效应( Haldane effect)。从图5-16可以看出,在相同PCO2下,动脉血(HbO2)携带的CO2比静脉血少。这主要是因为HbO2酸性较强,而脱氧Hb酸性较弱的缘故。所以脱氧Hb易和CO2结合生成 HbNHCOOh ,也易于和H+结合,使H2CO2解离过程中产生的H+被及时移去,有利于反应向右进行,提高了血液运输CO2的量。于是,在组织中,由于HbO2释出O2而成去氧Hb,经何尔登效应促使血液摄取并结合CO2;在肺,则因Hb与 O2结合,促使CO2释放。可见O2和CO2的运输不是孤立进行的,而是相互影响的。CO2通过波尔效效影响O2的结合和释放,O2又通过何尔登效应影响CO2的结合和释放。两者都与Hb的理化特性有关
第四节 呼吸运动的调节
呼吸运动是一种节律性的活动,其深度和频率随体内、外环境条件的改变而改变例如劳动或运动时,代谢增强,呼吸加深加快,肺通气量增大,摄取更多的O2,排出更多的CO2,以与代谢水平相适应。呼吸为什么能有节律地进行?呼吸的浓度和频率又如何能随内、外环境条件而改变?这些总是是本节的中心。
一、呼吸中枢与呼吸节律的形成
呼吸中枢是指中枢神经系统内产生和调节呼吸运动的神经细胞群。多年来,对于这些细胞群在中枢神经系统内的分布和呼吸节律产生和调节中的作用,曾用多种技术方法进行研究。如早期的较为粗糙的切除、横断、破坏、电刺激等方法,和后来发展起来的较为精细的微小电毁损、微小电刺激、可逆性冷冻或化学阻滞、选择性化学刺激或毁损、细胞外和细胞内微电极记录、逆行刺激(电刺激轴突,激起冲动逆行传导至胞体,在胞体记录)、神经元间电活动的相关分析以及组织化学等方法。有管些方法对动物呼吸中枢做了大量的实验性研究,获得了许多宝贵的资料,形成了一些假说或看法。
(一)呼吸中枢
呼吸中枢分布在大脑皮层、间脑、脑桥、延髓和脊髓等部位。脑的各级部位在呼吸节律产生和调节中所起作用不同。正常呼吸运动是在各级呼吸中枢的相互配合下进行的。
1.脊髓 脊髓中支配呼吸肌的运动神经元位于第3-5颈段(支配膈肌)和胸段(支配肌间肌和腹肌等)前角。很早就知道在延髓和脊髓间横断脊髓,呼吸就停止。所以,可以认为节律性呼吸运动不是在脊髓产生的。脊髓只是联系上(高)位脑和呼吸肌的中继站和整合某些呼吸反射的初级中枢。
2.下(低)位脑干 下(低)位脑干指脑桥和延髓。横切脑干的实验表明,呼吸节律产生于下位脑干,呼吸运动的变化因脑干横断的平面高低而异(图5-17)。
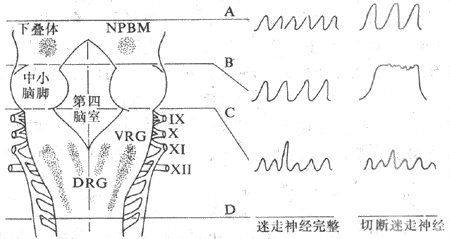
图5-17脑干呼吸有关核团(左)和在不同平面横切脑干后呼吸的变化(右)示意图
DRG:背侧呼吸组 VRH:腹侧呼吸组NPBM:臂旁内侧核A、B、C、D为不同平面横切
在动物中脑和脑桥之间进行横切(图5-17,A平面),呼吸无明显变化。在延髓和脊髓之间横切(D平面),呼吸停止。上述结果表明呼吸节律产生于下位禽干,上位脑对节律性呼吸不是必需的。如果在脑桥上、中部之间横切(B平面),呼吸将变慢变深,如再切断双侧迷走神经,吸气便大大延长,仅偶尔为短暂的呼气所中断,这种形式的呼吸称为长吸呼吸。这一结果是提示脑桥上部有抑制吸气的中枢结构,称为呼吸整中枢;来自肺部的迷走传入冲动也有抑制吸气的作用,当延髓失去来自这两方面对吸气活动的抑制作用后,吸气活动不能及时中断,便出现长吸呼吸。再在脑桥和延髓之间横切(C平面),不论迷走神经是否完整,长吸式呼吸都消失,而呈喘息样呼吸,呼吸不规则,或平静呼吸,或两者交替出现。因而认为脑桥中下中有活化吸气的长吸中枢;单独的延髓即可产生节律呼吸。孤立延髓的实验进一步证明延髓可独立地产生节律呼吸。于是在20-50年代期间形成了三级呼吸中枢理论;脑桥上部有呼吸调整中枢,中下部有长吸中枢,延髓有呼吸节律基本中枢。后来的研究肯定了早期关于延髓有呼吸节律基本中枢和脑桥上部有呼吸调整中枢的结论,但未能证实脑桥中下部存在着结构上特定的长吸中枢。
近年来,用微电极等新技术研究发现,在中枢神经系统内有的神经元呈节律性放电,并和呼吸周期相关,这些神经元被称为呼吸相关神经元或呼吸神经元。这些呼吸神经元有不同类型。就其自发放电的时间而言,在吸气相放电的为吸气神经元,在呼气相放电的为呼气神经元,在吸气相放电并延续至呼气相的为吸气-呼气神经元,在呼气相放电并延续到吸气相者,为呼气-吸气神经元,后两类神经元均系跨时相神经元。
在延髓,呼吸神经元内主要集中在背侧(孤束核的腹外侧部)和腹侧(疑核、后疑核和面神经后核附近的包氏复合体)两组神经核团内,分别称为背侧呼吸组(dorsal respiratory group,DRG)和腹侧呼吸组(ventralrespiratory group,VRG)(图5-17)。背侧呼吸组的神经元轴突主要交叉到对侧,下行至脊髓颈段,支配膈运动神经元。疑核呼吸神经元的轴突由同侧舌咽神经和迷走神经传出,支配咽喉部呼吸辅助肌。后疑核的呼吸神经元绝大部分交叉到对侧下行,支配脊髓肌间内、外肌和腹肌的运动神经元,商品化纤维也发出侧支支配膈肌的运动神经元。包氏复合体主要含呼气神经元,它们的轴突主要与背侧呼吸组的吸气神经元形成抑制性联系,此外也有轴突支配脊髓的膈运动神经元。
由于延髓呼吸神经元主要集中在背侧呼吸组和腹侧呼吸组,所以曾推测背侧呼吸组和腹侧呼吸组是产生基本呼吸节律的部位。可是,后来的某些实验结果不支持这一看法。有人用化学的或电解的毁损这些区域后,呼吸节律没有明显变化,这些结果提示背侧呼吸组和腹侧呼吸组可能不是呼吸节律唯一发源地,呼吸节律可能源于多个部位,产生呼吸节律的神经结构相当广泛,所以不容易因局灶损害而丧失呼吸节律。
在脑桥上 部,呼吸神经元相对集中于臂旁内侧核和相邻的Kolliker-Fuse(KF)核,合称PBKF核群。PBKF和延髓的呼吸神经核团之间有双向联系,形成调控呼吸的神经元回路。在麻醉猫,切断双侧迷走神经,损毁PBKF可出现长吸,提示早先研究即已发现的呼吸调整中枢乃位于脑桥的BPKF,其作用为限制吸气,促使吸气向呼气转换。
3.上位脑 呼吸还受脑桥以上部位的影响,如大脑皮层、边缘系统、下丘脑等。
大脑皮层可以随意控制呼吸,发动说、唱等动作,在一定限度内可以随意屏气或加强加快呼吸。大脑皮层对呼吸的调节系统是随意呼吸调节系统,下位脑干的呼吸调节系统是自主节律呼吸调节系统。这两个系统的下行通路是分开的。临床上有时可以观察到自主呼吸和随意呼吸分离的现象。例如在脊髓前外侧索下行的处主呼吸通路受损后,自主节律呼吸甚至停止,但病人仍可进行随意呼吸。患者靠随意呼吸或人工呼吸来维持肺通气,如未进行人工呼吸,一旦病人入睡,可能发生呼吸停止。
(二)呼吸节律形成的假说
呼吸节律是怎样产生的,尚未完全阐明,已提出多种假说,当前最为流行的是局部神经元回路反馈控制假说。
中枢神经系统里有许多神经元没有长突起向远处投射,只有短突起在某一部位内形成局部神经元回路联系。回路内可经正反馈联系募集更多神经元兴奋,以延长兴奋时间或加强兴奋活动;也可以负反馈联系,以限制其活动时间或终止其活动。平静呼吸时,由于吸气是主动的,所以许多学者更多地是去研究吸气中如何发生的,又如何转变为呼气的。有人提出中枢吸气活动发生器和吸气切断机制(inspiratory off-switch mechanism)的看法,认为在延髓有一个中枢吸气活动发生器,引发吸气神经元呈斜坡样渐增性放电,产生吸气;还有一个吸气切断机制,使吸气切断而发生呼气。在中枢吸气活动发生器作用下,吸气神经元兴奋,其兴奋传至①脊髓吸气肌运动神经元,引起吸气,肺扩张;②脑桥臂旁内侧核,加强其活动;③吸气切断机制,使之兴奋。吸气切断机制接受来自吸气神经元,脑桥背旁内侧核,和肺牵张感觉器的冲动。随着吸气相的进行,来自这三方面的冲动均逐渐增强,在吸气切断机制总合达到阈值时,吸气切断机制兴奋,发出冲动到中枢吸气活动发生器或吸气神经元,以负反馈形式终止其活动,吸气停止,转为呼气(图5-18)。切断迷走神经或毁损脑桥臂旁内侧核或两者,吸气切断机制达到阈值所需时间延长,吸气因面延长,呼吸变慢。因此,凡可影响中枢吸气活动发生器、吸气切断机制阈值或达到阈值所需时间的因素,都可影响呼吸过程和节律。
关于呼气如何转入吸气,呼吸加强时呼气又如何成为主动的,目前了解料少。
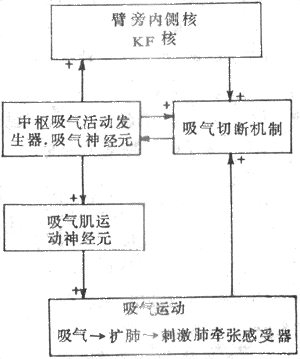
图5-18 呼吸节律形成机制简化模式图
+:表示兴奋 -:表示抑制
二、呼吸的反射性调节
呼吸节律虽然产生于脑,但其活动可受来自呼吸器官本身以及骨骼肌、其它器官系统感觉器‘传入冲动的反射性调节,下述其中的一些重要反射
(一)肺牵张反射
1868年Breuer和Hering发现,在麻醉动物肺充气或肺扩张,则抑制吸气;肺放气或肺缩小,则引起吸气。切断迷走神经,上述反应消失,所以是反射性反应。由肺扩张或肺缩小引起的吸气抑制或兴奋的反射为黑-伯反射(Hering-Breuer reflex)或肺牵张反射。它有两种成分:肺扩张反射和肺缩小反射。
1.肺扩张反射 是肺充气或扩张时抑制吸气的反射。感觉器位于从气管到细支气管的平滑肌中,是牵张感受器,阈值低,适应慢。当肺扩张牵拉呼吸道,使之也扩张时,感觉器兴奋,冲动经迷走神经走神经粗纤维传入延髓。在延髓内通过一定的神经联系使吸气切断机制兴奋,切断吸气,转入呼气。这样便加速了吸气和呼气的交替,使呼吸频率增加。所以切断迷走神经后,吸气延长、加深,呼吸变得深而慢。
有人比较了8种动物的肺扩张反射,发现有种属差异,兔的最强,人的最弱。在人体,当潮气量增加至800ml以上时,才能引起肺扩张反射,可能是由于人体肺扩张反射的中枢阈值较高所致。所以,平静呼吸时,肺扩张反射不参与人的呼吸调节。但在初生婴儿,存在这一反射,大约在出生4-5天后,反射就显著减弱。病理情况下,肺顺应性降低,肺扩张时使气道扩张较大,刺激较强,可以引起该反射,使呼吸变浅变快。
2.肺缩小反射 是肺缩小时引起吸气的反射。感受器同样位于气道平滑肌内,但其性质尚不十分清楚。肺缩小反向在较强的缩肺时才出现,它在平静呼吸调节中意义不大,但对阻止呼气过深和肺不张等可能起一定作用。
(二)呼吸肌本体感受性反射
肌梭和腱器官是骨骼肌的本体感受器,它们所引起的反射为本体感受性反射。如肌梭受到牵张刺激时可以反射性地引起受刺激肌梭所在肌的收缩,为牵张反射,属本体感受性反射(参见第十章第四节)。呼吸肌也有牵张反射的主要依据是:在麻醉猫,切断双侧迷走神经,颈7横断脊髓,牵拉膈肌,膈肌肌电活动啬;切断动物的胸脊神经背根,呼吸运动减弱;人类为治病需要曾做类似手术,术后相应呼吸肌的活动发生可恢复的或可部分恢复的减弱。说明呼吸肌本体感受性反射参与正常呼吸运动的调节,在呼吸肌负荷改变时将发挥更大的作用。但是,这些依据不是无懈可击的。因为背根切断术不仅切断了本本感受器的传入纤维,也切断了所有经背根传入的其它感受器的传入纤维。近来的研究表明来自呼吸肌其它感受器的传入冲动也可反射性地影响呼吸。因此,对呼吸肌本体感受性反射应做更深更深入细致的研究,如研究分别兴奋不同感受器或传入纤维时对呼吸的效应。
(三)防御性呼吸反射
在整个呼吸道都存在着感受器,它们是分布在粘膜上皮的迷走传入神经末梢,受到机械或化学刺激时,引起防御性呼吸反射,以清除激惹物,避免其进入肺泡。
1.咳嗽反射 是常见的重要防御反射。它的感受器位于喉、气管和支气管的粘膜。大支气管以上部位的感受器对机械刺激敏感,二级支气管以下部位的对化学刺激敏感。传入冲动经迷走神经传入延髓,触发一系列协调的反射反应,引起咳嗽反射。
咳嗽时,先是短促或深吸气,接着声门紧闭,呼气肌强烈收缩,肺内压和胸膜腔内压急速上升,然后声门突然打开,由于气压差极大,气体更以极高的速度从肺内冲出,将呼吸道内异物或分泌物排出。剧烈咳嗽时,因胸膜腔内压显著升高,可阻碍静脉因流,使静脉压和脑脊液压升高。
2.喷嚏反射 是和咳嗽类似的反射,不同的是:刺激作用于鼻粘膜感受器,传入神经是三叉神经,反射效应是腭垂下降,舌压向软腭,而不是声门关闭,呼出气主要从鼻腔喷出,以清除鼻腔中的刺激物。
(四)肺毛细血管旁(J-)感受器引起的呼吸反射
J-感受器位于肺泡毛细血管旁,在肺毛细血管充血、肺泡壁间质积液时受到刺激,冲动经迷走神经无髓C纤维传入延髓,引起反射性呼吸暂停,继以浅快呼吸,血压降低,心率减慢。J-感受器在呼吸调节中的作用尚不清楚,可能与运动时呼吸加快作肺充血、肺水肿时的急促呼吸有关。
(五)某些穴位刺激的呼吸效应
针刺人中窕可以急救全麻手术过程中出现的呼吸停止。针刺动物人中可以使膈肌呼吸运动增强,电刺激家兔人中对膈神经和管髓呼吸神经元电活动有特异性影响。有人观察到在麻醉意外事件发生呼吸暂停时,刺激素可以兴奋呼吸。穴位的呼吸效应及其机制值得探讨。
(六)血压对呼吸的影响
血压大幅度变化时可以反射性地影响呼吸,血压升高,呼吸减弱减慢;血压降低,呼吸加强加快。
三、化学因素对呼吸的调节
化学因素对呼吸的调节也是一种呼吸的反射性调节,化学因素是指动脉血或脑脊液中的O2、CO2和H+。机体通过呼吸调节血液中的O2、CO2和H+的水平,动脉血中O2、CO2和H+水平的变化又通过化学感受器调节着呼吸,如此形成的控制环维持着内环境这些因素的相对稳定。
(一)化学感受器
化学感觉器是拂晓春适宜刺激化学物质的感受器。参与呼吸调节的化学感受器因其所在部位的不同,分为外周化学感受器和中枢化学感受器。
1.外周化学感受器 颈动脉体和主动脉体是调节呼吸和循环的重要外周化学感受器。在动脉血PO2降低、PCO2或H+浓度([H+])升主时受到刺激,冲动经窦神经和迷走神经传入延髓,反射性地引起呼吸加深加快和血液循环的变化。虽然颈、主动脉体两者都参与呼吸和循环的调节,但是颈动脉体主要调节呼吸,而主动脉体在循环调节方面较为重要。由于颈动脉体的有利的解剖位置,所以,对外周化学感受器的研究主要集中在颈动脉体。
颈动脉体含Ⅰ型细胞(球细胞)和Ⅱ型细胞(鞘细胞),它们周围包绕以毛细血管窦。血液供应十分丰富。Ⅰ型细胞呈球形,有大量囊泡,内含递质,如乙酰胆碱、儿茶酚胺、某些神经活性肽等。Ⅱ型细胞数量较少,没有囊泡。Ⅱ型细胞包绕着Ⅰ型细胞、神经纤维和神经末梢,功能上类似神经胶质细胞,与颈动脉体其它成分之间没有特化的接触。窦神经的传入纤维末梢分支穿插于Ⅰ、Ⅱ型细胞之间,与Ⅰ型细胞形成特化接触,包括单向突触、交互突触、缝隙边接等(图5-19),传入神经末梢可以是突触前和(或)突触后成分。交互突触构成Ⅰ型细胞与传入神经之间的一种反馈环路,借释放递质调节化学感受器的敏感性。此外,颈动脉体还有传出神经支配,借调节血流和化学感受器以改变化学感受器的活动。
用游离的颈动脉体,记录其传入神经单纤维的动作,观察改变灌流液成分时动作频率的变化,可以了解颈动脉体所感受的刺激的性质以及刺激与反应之间的关系。结果发现当灌流液PO2下降,PCO2或[H+]升高时,传入冲动增加。如果保持灌流血液的PO2正常的13.3kPa(100mlHg),仅减少血流量,传入冲动也增加。困为血流量下降时,颈动脉体从单位血液中摄取的O2量相对增加,细胞外液 PO2因供O2少于耗 O2而下降。但在贫血或CO中毒时,血 O2含量虽然下降,但PO2正常,只需血流量充分,化学感受器传入冲动并不增加,所以化学感受器所感受的刺激是PO2,而不是动脉血O2含量,而且是感受器所处环境的PO2。从实验中还可看出上述三种刺激对化学感受器有相互增强的作用。两种刺激同进作用时比单一刺激的效应强。这种协同作用有重要意义,因为机体发生循环或呼吸衰竭时,总是PCO2升高和PO2降低同进存在,它们的协同作用加强了对化学感受器的刺激,从而促进了代偿性呼吸增强的反应。
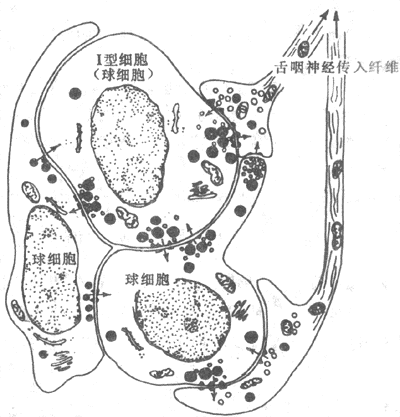
图5-19 颈动脉体组织结构示意图
图中未显示Ⅱ细胞
目前认为,Ⅰ型细胞起着化学感受器的作用。当它们受到刺激时,细胞浆内[Ca2+]升高。触发递质释放,引起传入神经纤维兴奋。PO2降低与 PCO2或[H+]升高引起细胞内[Ca2+]升高机制不同。PO2降低可抑制细胞 K+通道的开放,K+外流减少,细胞膜去极化,从而促使电压依从性Ca2+通道开放,Ca2+进入细胞。而PCO2或[H+]升高时,进入细胞内的H+增多,激活了细胞的Na+-H+交换机制,Na+进入细胞,使细胞内[Na+]长高,继而使细胞的Na+-Ca2+交换机制活动啬,Na+出细胞,Ca2+进细胞内,引起细胞浆内[Ca2+]升高。还有资料表明,少部分胞浆内Ca2+可能来自细胞内的Ca2+贮器。
2.中枢化学感受器 摘除动物外周化学感受器或切断其传入神经后,吸入CO2仍能加强通气。改变脑脊液CO2和H+浓度也能刺激呼吸。过去认为这是CO2直接刺激呼吸中枢所致年代以来,用改变脑表面灌流液成分和pH、局部冷阻断、电凝固损伤、电刺激、记录神经元电活动、离体脑组织块的电生理研究等方法在多种动物做了大量实验,结果表明在延髓有一个不同于呼吸中枢,但可影响呼吸的化学感受器,称为中枢化学感受器,以另于外周化学感受器。
中枢化学感受器 位于延髓腹外侧浅表部位,左右对称,可以分为头、中、尾三个区(图5-20A)。头端和尾端区都有化学感受性,中间区不具有化学感受性,不过,局部阻滞或损伤中间区后,可以使动物通气量降低,并使头端、尾端区受刺激时的通气反应消失,提示中间区可能是端区和尾 端区传入冲动向脑干呼吸中枢投射的中继站。应用胆碱能激动剂和拮抗剂的研究结果表明,在中枢化学感受器传递环节中可能有胆碱能机制参与。
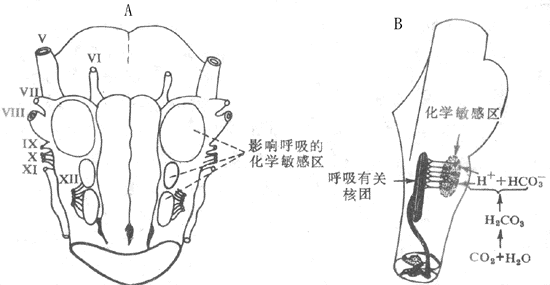
图5-20 中枢化学感受器
A示延髓腹外侧的三个化学敏感区 B示血液或
脑脊液PCO2升高时,刺激呼吸的中枢机制
中枢化学感受器的生理刺激是脑脊液和局部细胞外液的H+。因为如果保持人工脑脊液的pH不变,用含高浓度CO2的人工脑脊液灌流脑室时所引起的通气增强反应消失,可见有效刺激不是CO2本身,而是CO2所引起的[H+]的增加。在体内,血液中的CO2能迅速通过血脑屏障,使化学感受器周围液体中的[H+]升高,从而刺激中枢化学感受器,再引起呼吸中枢的兴奋(图5-20B)。可是,脑脊液中碳酸酶含量很少,CO2与水的水合反应很慢,所以对CO2的反应有一定的时间延迟。血液中的H+不易以通过血液屏障,故血液pH的变化对中枢化学感受器的直接作用不大,也较缓慢。
中枢化学感受器与外周化学感受器不同,它不感受缺O2的刺激,但对CO2的敏感性比外周的主,反应潜伏期较长。中枢化学感受器的作用可能是调节脑脊液的[H+],使中枢神经系统有一稳定的pH环境,而外周化学感受器的作用主要是在机体低O2时,维持对呼吸的驱动。
(二)CO2、H+和O2对呼吸的影响
1.CO2的影响 很早已经知道,在麻醉动物或人,动脉血液PCO2降得很低时可发生呼吸暂停。因此,一定水平的PCO2对维持呼吸和呼吸中枢的兴奋性是必要的,CO2是调节呼吸的最重要的生理性体液因子。
吸入含CO2的混合气,将使肺泡气PCO2长高,动脉血PCO2也随之升高,呼吸加深加快,肺通气量增加(图5-21)。通过肺通气量的增大可能增加CO2的清除,肺泡气和动脉血PCO2还可维持于接近正常水平。但是,当吸入气CO2陡升,CO2堆积,压抑中枢神经系统的活动,包括呼吸中枢,发生呼吸困难、头痛、头昏,甚至昏迷,出现CO2麻醉。对CO2的反应,有个体差异,还受许多因素影响,如疾病或药物。总之CO2在呼吸调节中是经常起作用的最重要的化学刺激,在一定范围内动脉血PCO2的升高,可以加强对呼吸的刺激作用,但超过一定限度则有压抑和麻醉效应。
CO2刺激呼吸是通过两条途径实现的,一是通过刺激中枢化学感受器再兴奋呼吸中枢:二是刺激外周化学感受器,冲动窦神经和迷走神经传入延髓呼吸有关疑团,反射性地使呼吸加深、加快,增加肺通气。但两条途径中前者是主要的。因为去掉外周化学感受器的作用之后,CO2的通气反应仅下降约20%,可见中枢化学感受器在CO2通气反应中起主要作用;动脉血PCO2只需升高 0.266kPa(2mmHg)就可刺激中枢化学感受器,出现通气加强反应,如刺激外周化学感受器,则需升高1.33kPa(10mmHg)。不过,在下述情况下,外周化学感受器的作用可能是重要的:因为中枢化学感受器的反应慢,所以当动脉血PCO2突然大增时,外周化学感受器在引起快速呼吸反应中可起重要作用;当中枢化学感受器到抑制,对CO2的反应降低时,外周化学感受器就起重要作用。
2.H+的影响 动脉血[H+]增加,呼吸加深加快,肺通气增加;[H+]降低,呼吸受到抑制(图5-21)。H+对呼吸的调节也是通过外周化学感受器和中枢化学感受器实现的。中枢化学感受器对H+的敏感性较外周的高,约为外周的25倍。但是,H+通过血液屏障的速度慢,限制了它对中枢化学感受器的作用。脑脊液中的H+才是中枢化学感受器的最有效的刺激。
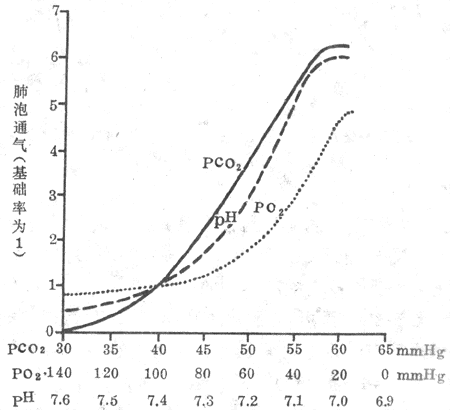
图5-21 动脉血液PCO2、PO2、pH改变对肺泡通气
的影响仅改变一种体液因素而保持另二因素于
正常水平时的情况(1mmHg=0.133kPa)
3.O2的影响 吸入气PO2降低时,肺泡气PO2都随之降低,呼吸加深、加快,肺通气增加(图5-21)。同CO2一样,对低O2的反应也有个体差异。一般在动脉PO2下降到10.64kPa(80mmHg)以下时,肺通气才出现可觉察到的增加,可见动脉血PO2对正常呼吸的调节作用不大,仅在特殊情况下低O2刺激才有重要意义。如严重肺气肿、肺心病患者,肺换气受到障碍,导致低O2和CO2潴留。长时间CO2潴留使中枢化学感受器对CO2的刺激作用发生适应,而外周化学感受器对低O2刺激适应很慢,这时低O2对外周化学感受器的刺激成为驱动呼吸的主要刺激。
低O2对呼吸的刺激作用完全是通过外周化学感受器实现的。切断动物外周化学感受器的传入神经或摘除人的颈动脉体,急性低O2的呼吸刺激反应完全消失。低O2对中枢的直接作用是压抑作用。但是低O2可以通过对外周化学感受器的刺激而兴奋呼吸中枢,这样在一定程度上可以对抗低O2对中枢的直接压抑作用。不过在严重低O2时,外周化学感受性反射已不足以克服低O2对中枢的压抑作用,终将导致呼吸障碍。在低O2时吸入纯O2,由于解除了外周化学感受器的低O2刺激,会引起呼吸暂停,临床上给O2治疗时应予以注意。
(三)PCO2、H+和PO2在影响呼吸中的相互作用
图5-21示保持其它两个因素不变而只改变其中一个因素时的单因素通气效应。可以看出PO2下降对呼吸的影响较慢、较弱,在一般动脉血PO2变化范围内作用不大,要在PO2低于10.64kPa(80mmHg)后,通气量才逐渐增大。PCO2和H+与低O2不同,只要略有升高,通气就明显增大,PCO2的作用尤为突出。
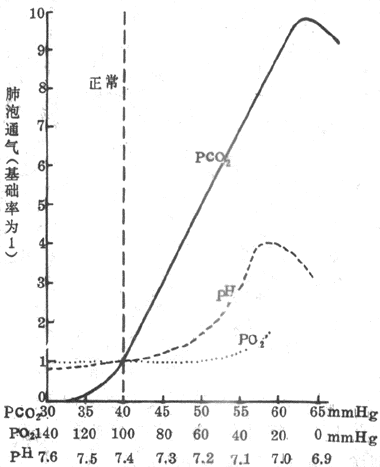
图5-22 动脉血液PCO2升高、PCO2降低、pH降低对肺泡通气率的影响
(1mmHg=0.133kpa)
但实际情况不可能是单因素的改变,而其它因素不变。往往是一种因素的改变会引起其余一、两种因素相继改变或存在几种因素的同时改变,三者间相互影响、相互作用,既可因相互总和而加大,也可因相互抵消而减弱。图5-22为一种因素改变,另两种因素不加控制时的情况。可以看出:PCO2升高时,[H+]也随之升高,两者的作用总和起来,使肺通气较单独PCO2升高时为大。[H+]增加时,因肺通气增大使CO2排出,PCO2下降,抵消了一部分H+的刺激作用;CO2含量的下降,也使[H+]有所降低。两者均使肺通气的增加较单独[H+]升高时为小。PO2下降时,也因肺通气量增加,呼出较多的CO2,使PCO2和[H+]下降,从而减弱了低O2的刺激作用。
四、周期性呼吸
周期性呼吸是异常呼吸型之一,表现为呼吸加强加快与减弱减慢交替出现。最常见的有陈-施呼吸和比奥(Biot)呼吸。
(一)陈-施呼吸(潮式呼吸)
陈-施呼吸(Cheyne-Stokes respiration)的特点是呼吸逐渐增强增快又逐渐减弱减慢与呼吸暂停交替出现,每个周期约45s到3min。
当前认为陈-施呼吸产生的基本机制是因为某种原因呼吸受到刺激,肺通气量增加,呼出过多的CO2,肺泡气PCO2下降,肺部血液PCO2也下降,片刻之后,这种低PCO2血液到达脑部?
■[此处缺少一些内容]■
尚不清楚,可能是疾病已侵及延髓,损害了呼吸中枢。
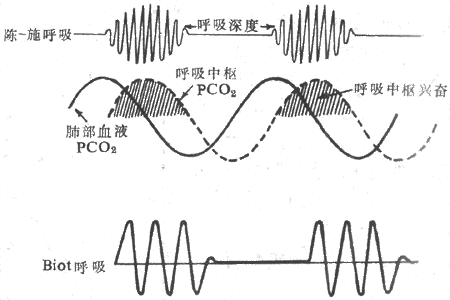
图5-23 异常呼吸型式示意图
五、运动时呼吸的变化及调节
运动时机体代谢增高,血液循环和呼吸系统都将发生一系列变化以适应增高了的机体代谢的需要。这时,呼吸加深加快,肺通气量增大,其增加的程度随运动量而异。潮气量可双安静时的500ml升到2000ml,呼吸频率可从 12-18次/min升至50次/min,每分通气量可升达100L以上,O2的摄入量和CO2排出量也都相应增加。
运动时肺通气量的增加有一个过程。运动之始,通气量骤升,继之以缓慢的升高,随后达一稳态水平。运动停止时,也是通气量先骤降,继以缓慢下降,然后恢复到运动前的水平(图5-24)。
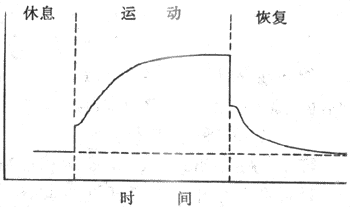
图5-24 运动时的肺通气量变化
运动时呼吸的变化是大家都很熟悉的生理现象,但其变化机制却至今仍未阐明,不过与其它生理活动一样,也是在神经和体液机制调节下发生的。
一般认为运动开始时通气骤升也条件反射有关,是在运动锻炼过程中形成的。因为只是给予运动暗示,并未开始运动,也可出现通气量增大的反应,而且与运动者过去的经验、精神状态、实验条件等有关。此外,运动时,运动肌肉、关节的本体感受器受到刺激,其传入冲动也可以反射性地刺激呼吸。因为,仅被动运动肢体,就可引起快速通气反应,阴断活动肢体的传入神经,反应消失;脊髓胸12水平截瘫病人,被动运动膝关节,不能产生快速通气反应。近年来神经组化研究表明,在延髓孤束周围有较稠密的肌传入末梢,这可能是运动肢体引起运动初期快速通气反应的解剖学基础。
运动时动脉血pH、PCO2PO2的波动的作用。中度运动时,虽然动脉血pH、PCO2PO2的均值保持相对的稳定,但它们却都能随呼吸厕于周期性波动,波动的幅度随运动强度而变化,运动强,波动幅度大;运动弱,波动幅度小。运动时,这种波动幅度的增大,可能在运动通气反应中起重要作用。动物实验中,设法在不影响血液气体平均分压的同时,缓冲上述周期性波动,动物通气量下降。记录猫颈动脉窦神经化学感受器传入冲动,发现其频率与呼吸周期同步消、涨。体液因素的这种作用是通过化学感受器实现的。
运动停止后,通气未立即恢复到安静水平。这是因为运动时,O2供小于O2耗,欠下了“O2债”。所以运动停止后,一段时间内,O2耗仍大于安静时的,以偿还O2债,待偿还后,通气才恢复。这时维持通气增强的刺激主要是H+。
参考资料
1.陈子彬延髓中枢化学感受器及其在呼吸调节中的作用,生理科学进展1982;13:300-305
2.张衡 关于哺乳动物呼吸中枢的现代观点生理科学进展 1986;17:210-215
3.郑煜 哺乳动物呼吸节律研究简介 生物学通报 1992;10:11-14
4.Numm JF,陈毓槐等译 应用呼吸生理学北京科学出版社,1987
5.Garby L,Meldon J,周殿松译 血液呼吸功能 北京 科学出版社 1987
6.BarnesPJ.Modulation of neurotransmission in airways.Physiol Rey 1992;72:699-729
7.Hay DWP,HenryPJ,Goldie RG.Endothelin and the respiratory system.TIPs 1993;14:29-32
8.gonzalez C,AlmarazL,Obeso A,Rigual R.Oxygen and acid chemoreception in the carotid bodychemoreceptors .TINS1992;15:146-153
9.CohenMI.Neurogenesis of respiratory rhythm in the mammal.Physiol Rev1979;59:1105-1173
10.Slonim NB,HamiltonLH.Respiratory Physiology 5thed,Cv Mosby Co,St Louis,1987
第六章 消化和吸收
第一节 概述
人的消化器官由长约8-10m的消化道及与其相连的许多大、小消化腺组成。消化器官的主要生理功能是对食物进行消化和吸收,从而为机体新陈代谢提供了必不可少的物质和能量来源。
消化是食物在消化道内被分解为小分子的过程。消化的方式有两种。一种是通过消化道肌肉的舒缩活动,将食物磨粹,并使之与消化液充分混合,以及将食物不断地向消化道的远端推送;这种方式称机械消化。另一种消化方式是通过消化腺分泌的消化液完成的。消化液中含在各种消化酶,能分别分解蛋白质、脂肪和糖类等物质,使之成为小分子物质(表6-1);这种消化方式称化学性消化。正常情况下,这两种方式的消化作用是同时进行,互相配合的。食物经过消化后,透过消化道的粘膜,进入血液和淋巴循环的过程,称为吸收。消化和吸收是两个相辅相成、紧密联系的过程。不能被消化和吸收的食物残渣,最后以粪的形式排出体外。
表6-1 消化液的成分及其作用
| 消化液 | 分泌量(L/d) | PH | 主要成分 | 酶的底物 | 酶的水解产物 |
| 唾液 | 1.0-1.5 | 6.6-7.1 | 沾液 | ||
| α-淀粉酶 | 淀粉 | 麦芽糖 | |||
| 胃液 | 1.5-2.5 | 0.9-1.5 | 粘液、盐酸 | ||
| 胃蛋白酶(原) | 蛋白质 | 、胨、多肽 | |||
| 内因子 | |||||
| 胰液 | 1.0-2.0 | 7.8-8.4 | HCO3 | ||
| 胰蛋白酶(原) | 蛋白质 | 氨基酸、寡肽 | |||
| 糜蛋白酶(原) | |||||
| 羧基肽酶(原) | 肽 | 氨基酸 | |||
| 核糖核酸酶 | RNA | 单核苷酸 | |||
| 脱氧核糖核酸酶 | DNA | ||||
| α-淀粉酶 | 淀粉 | 麦芽糖、寡糖 | |||
| 胰脂肪酶 | 甘油三酯 | 脂肪酸、甘油、甘油一酯 | |||
| 胆固醇酯酶 | 胆固醇酯 | 脂肪酸、胆固醇 | |||
| 磷脂酶 | 磷脂 | 脂肪酸、溶血磷脂 | |||
| 胆汁 | 0.8-1.0 | 6.8-7.4 | 胆盐 | ||
| 胆固醇 | |||||
| 胆色素 | |||||
| 小肠液 | 1.0-3.0 | 7.6 | 粘液 | ||
| 肠激酶 | 胰蛋白酶原 | 胰蛋白酶 | |||
| 大肠液 | 0.5 | 8.3 | 粘液 | ||
| HCO3 |
一、消化道平滑肌的特性
在整个消化道中,除口、咽、食管上端和肛门外括约肌是骨骼肌外,其余部分是都是由平滑肌组成的。消化道通过这些肌肉的舒缩活动,完成对食物的机械性消化,并推动食物的前进;消化道的运动对于食物的化学性消化和吸收,也有促进作用。
(一)消化道平滑肌的一般特性
消化道平滑肌具有肌组织的共同特性,如兴奋、自律性、传导性和收缩性,但这些特性的表现均有其自己的特点。
1.消化道平滑肌的兴奋较骨骼肌为低。收缩的潜伏期、收缩期和舒张期所占的时间比骨骼肌的长得多,而且变异很大。
2.消化道平滑肌在离体后,置于适宜的环境内,仍能进行良好的节律性运动,但其收缩很缓慢,节律性远不如心肌规则。
3.消化道平滑肌经常保持在一种微弱的持续收缩状态,即具有一定的紧张性。消化道各部分,如胃、肠等之所以能保持一定的形状和位置,同平滑肌的紧张性在重要的关系;紧张性还使消化道的管腔内经常保持着一定的基础压力;平滑肌的各种收缩活动也就是在紧张性基础上发生的。
4.消化道平滑肌能适应实际的需要而作很很大的伸展。作为中空的容纳器官来说,这一特性具有重要生理意义。它的消化道有可能容纳好几倍于自己原初体积的食物。
5.消化道平滑肌对电刺激较不敏感,但对于牵张、温度和化学刺激则特别敏感,轻微的刺激常可引起强烈的收缩。消化道平滑肌的这一特性是与它所处的生理环境分不开的,消化道内容物对平滑肌的牵张、温度和化学刺激是引起内容物推进或排空的自然刺激因素。
(二)消化道平滑肌的电生理特性
消化道平滑肌电活动的形式要比骨骼肌复杂得多,其电生理变化大致可分为三种,即静息膜电位、慢波电位和动作电位。
1.静息膜电位消化道平滑肌的静息膜电位很不稳定,波动较大,其实测值为-60—-50Mv,静息电位主要由K+的平衡电位形成,但Na+、CI-、Ca2+以及生电性钠泵活动也参与了静息膜电位的产生。
2.慢波电位消化道的平滑肌细胞可产生节律性的自发性去极化;以静息膜电位为基础的这种周期性波动,由于其发生频率较慢而被称为慢波电位,又称基本电节律(basal electric rhythm,BER)。消化道不部位的慢波频率不同,在人类,胃的慢波频率为3次/min,十二指肠为12次/min,回肠末端为8-9次/min。慢波的波幅约为10-15mV,持续时间由数秒至十几秒。
用细胞内微电极记录时,慢波多表现为单向波,包括初期的快速去极化和缓慢的复极化平台。关于慢波产生的离子基础尚未完全清楚。目前认为,它的产生可能与细胞膜上生电性钠泵的活动具有波动性有关,当钠泵的活动暂时受抑制时,膜便发生去极化;当钠泵活动恢复时,膜的极化加强,膜电位便又回到原来的水平。实验证明,用抑制钠泵的药物哇巴因后,胃肠平滑肌的慢波电位消失。
在通常情况下,慢波起源于消化道的纵行肌,以电紧张形式扩布到环行肌。由于切断支配胃肠的神经,或用药物阻断神经冲动后,慢波电位仍然存在,表明它的产生可能是肌源性的。慢波本身不引起肌肉收缩,便它可以反映平滑肌兴奋性的周期变化。慢波可使静息膜电位接近于产生动作电位的阈电位,一旦达到阈电位,膜上的电压依从性离子通道便开放而产生动作电位。
3.动作电位平滑肌的动作电位与神经和骨骼肌的动作电位的区别在于:①锋电位上升慢,持续时间长;②平滑肌的动作电位不受钠通道阻断剂的影响,但可被Ca2+通道阻断剂所阻断,这表明它的产生主要依赖Ca2+的内流;③不滑肌动作电位的复极化与骨骼肌相同,都是通过K+的外流,所不同的是,不滑肌K+的外向电流与Ca2+的内向电流在时间过程上几乎相同,因此,锋电位的幅度低,而且大小不等。
由于平滑肌动作电位发生时Ca2内流的速度已足以引起平滑肌的收缩,因此,锋电位与收缩之间存在很好的相关性,每个慢波上所出现锋电位的数目,可作为收缩力大小的指标。
慢波、动作电位和肌肉收缩的关系可简要归纳为:平滑肌的收缩是继动作电位之后产生的,而动作电位则是在慢波去极化的基础上发生的。因此,慢波电位本身虽不能引起平滑肌的收缩,但却被认为是不滑肌的起步电位,是平滑肌收缩节律的控制波,它决定蠕动的方向、节律和速度(图6-1)。
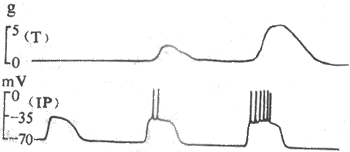
图6-1 消化道不滑肌的电活动
下面的曲线为细胞内电极记录的基本电节律,
在第二和第三个波的支极化期,出现数目不同的动作电位;
上面的曲线为肌肉收缩,收缩波只出现在动作电位时,
动作电位数目越多,收缩的幅度也越大T:张力 IP:细胞内电位
二、消化腺的分泌功能
人每日由各种消化腺分泌的消化液总量达6-8L(表6-1)。消化液主要由有机物、离子和水组成。消化液的主要功能为:①稀释食物,使之与血浆的渗透压相等,以利于吸收;②改变消化腔内的pH,使之适应于消化酶活性的需要;③水解复杂的食物成分,使之便于吸收;④通过分泌粘液、抗体和大量液体,保护消化道粘膜,防止物理性和化学性的损伤。
分泌过程是由腺细胞主动活动的过程,它包括由血液内摄取原料、在细胞内合成分泌物,以及将分泌物由细胞内排出等一连串的复杂活动。对消化腺分泌细胞的刺激-分泌耦联的研究表明,腺细胞膜上往往存在着多种受体,不同的刺激物与相应的受体结合,可引起细胞内一系列的生化反应,最终导致分泌物的释放(图6-2)。
图6-2 胰腺分泌细胞内的两种介导机制
三、胃肠的神经支配及其作用
神经系统对胃肠功能的调节较为复杂,它通过植物性神经和胃肠的内在神经两个系统相互协调统一而完成的(图6-3)。
图6-3 消化系统的局部和中枢性反射通路
胃肠的内在神经是由存在于食管至肛门的管壁内的两种神经丛组成的。一种是位于胃肠壁粘膜下神经丛(Meissner神经丛);另一种是位于环行肌与纵行肌层之间的肌间神经丛(或称Auerbach神经丛)。内在神经丛包含无数的神经元和神经纤维,据估计,内在神经丛中约有108个神经元,包括感觉神经元、中间神经元和运动神经元。内在神经丛的神经纤维(包括进入消化管壁的交感和副交感纤维)则把胃肠壁的各种感受器及效应细胞与神经元互相连接,起着传递感觉信息、调节运动神经元的活动和启动、维持或抑制效应系统的作用。目前认为,消化管壁内的神经丛构成了一个完整的、相对独立的整合系统,在胃肠活动的调节中具有十分重要的作用。
支配胃肠的自主神经被称为外来神经,包括交感神经和副交感神经。交感神经从脊髓胸腰段侧角发出,经过腹腔神经节、肠系膜神经节或腥下神经节,更换神经元后,节后纤维分布到胃肠各部分,主要通过三种途径影响胃肠活动:①终止于内在神经元的肾上腺素能纤维;②分布于某些肌束的肾上腺素能纤维;③分布至血管平滑肌的肾上腺素血管纤维。由交感神经节后纤维释放至内在神经元表面的去甲肾上腺素,可抑制神经元的兴奋活动,从而抑制其向前传导的活动。这样,由交感神经发放的冲动,可抑制通过内在神经丛或迷走神经传递的反射。
图6-4 胃肠胆碱能、肾上腺素能及肽能神经的支配及作用模式图
I:抑制 E:兴奋 CNS:中枢神经系统
——:胆碱能神经质 ----:肾上腺素能神经 +++肽能神经
副交感神经通过迷走神经和盆神经支配胃肠。到达胃肠的纤维都是节前纤维,它们终止于内在神经丛的神经元上。内在神经丛的多数副交感纤维是兴奋性胆碱能纤维,少数是抑制性纤维;而在这些抑制性纤维中,多数既不是胆碱能,也不是肾上腺素能纤维,它们的末梢释放的递质可能是肽类物质,因而被称为肽能神经(图6-4)。由肽能神经末梢释放的递质不是单一的肽,而可能是不同的肽,如血管活性肽(VIP)、P物质、脑啡肽和生长抑素等。目前认为,胃的容受性舒张、机械刺激引起的小肠充血等,均为神经兴奋释放VIP所致,VIP能神经的作用主要是舒张平滑肌、舒张血管和加强小肠、胰腺的分泌活动(图6-5)。
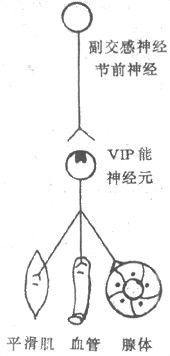
图6-5 血管活性肠肽(VIP)能神经支配的设想图
四、胃肠激素
在胃肠的粘膜层内, 不仅存在多种外分泌腺体,还含有十种内分泌细胞,这些细胞分泌的激素统称为胃肠激素(gastrointestinal hormone)。胃肠激素在化学结构上都是由氨基酸残基组成的肽类,分子量大多数在5000以内。
(一)胃肠内分泌细胞的形态及分布
用细胞免疫细胞化学的方法已经证明,从胃到大肠的粘膜层内,存在有40多种内分泌细胞,它们分散地分布在胃肠粘膜的跑非内分泌细胞之间。由于胃肠粘膜的面积巨大,胃肠内分泌细胞的总数很大有大,大大地超过了体内所有内分泌腺的总和。因此,消化道已不仅仅是人体内的消化器官,它也是体内最大最复杂的内分泌器官(表6-2)。
胃肠内分泌细胞在形成上有两个明显的特点,一是细胞内的分泌颗粒均分布在核和基底之间,故属于基底颗粒细胞。不同的内分泌细胞的分泌颗粒大小、形状和密度均不同。胃肠内分泌细胞的另一特点是,大部分细胞呈锥形,其顶端有绒毛突起,伸入胃肠腔内(图6-6),微绒毛可直接感受胃肠内食物成分和pH的一刺激而引起细胞的分泌活动。只有少数胃肠内分泌细胞无微绒毛,它们与胃肠腔无直接接触,它们的分泌可由神经兴奋或局部内环境的变化而引起,而与胃肠腔内的食物成分无关。这两种类型的细胞,前者被称为开放型细胞,后者为闭合型细胞。
表6-2 主要胃肠内分泌细胞的名称、分布和分泌产物
| 细胞名称 | 分泌产物 | 分布部位 |
| A细胞 | 胰高血糖素 | 胰岛 |
| B细胞 | 胰岛素 | 胰岛 |
| D细胞 | 生长 抑素 | 胰岛、胃、小肠、结肠 |
| G细胞 | 胃泌素 | 胃窦、十二指肠 |
| I细胞 | 胆囊收缩素 | 小肠上部 |
| K细胞 | 抑胃肽 | 小肠上部 |
| Mo细胞 | 胃动素 | 小肠 |
| N细胞 | 神经降压素 | 回肠 |
| PP细胞 | 胰多肽 | 胰岛、胰腺外分沁部分、胃、小肠、大肠 |
| S细胞 | 促胰液素 | 小肠上部 |
图6-6 胃窦粘膜内的G细胞(开放型细胞)示细胞顶端的绒毛
胃肠内分泌细胞在生物化学方面都具有摄取胺前体,进行脱羟而产生肽类或活性胺的能力。具有这种能力的细胞统称为APUD(amine precursor uptake and decarboxy-lation)细胞。除胃肠和胰腺的内分泌细胞外,神经系统、甲状腺、肾上腺髓质、垂体等组织中也含有APUD细胞。
(二)胃肠激素的作用
胃肠激素与神经系统一起,共同调节消化器官的运动、分泌和吸收功能。此外,胃肠激素对体内其它器官的活动也具有广泛的影响。其作用有三个主要方面。
1.调节消化腺的分泌和消化道的运动 这一作用的靶器官包括唾液腺、胃腺、胰腺、肠腺、肝细胞、食管-胃括约肌、胃肠平滑肌及胆囊等。三个主要胃肠激素的作用见表6-3及后文。
表6-3 三种胃肠对消化腺分泌和消化管运动地作用
| 胃酸 | 胰HCO3 | 胰酶 | 肝胆汁 | 小肠液 | 食管-括约肌 | 胃运动 | 小肠运动 | 胆囊收缩 | |
| 胃沁素 | ╂╂ | ╂ | ╂╂ | ╂ | ╂ | ╂ | ╂ | ╂ | ╂ |
| 促胰液素 | — | ╂╂ | ╂ | ╂ | ╂ | — | — | — | ╂ |
| 胆囊收缩素 | ╂ | ╂ | ╂╂ | ╂ | ╂ | — | ╂ | ╂ | ╂╂ |
╂:兴奋 ╂╂:强兴奋 —:抑制
2.调节其它激素的释放已经证明,食物消化时,从胃肠释放的抑胃肽(gastric inhibitory polypeptide,GIP)有很强的刺激胰岛素分泌的作用。因此,口服葡萄糖比静脉注射相同剂量的葡萄糖,能引起更多的胰岛素分泌。进餐时,不仅由于葡萄糖的吸收入血直接作用于胰岛B细胞,促进其分泌胰岛素,而且还可通过抑胃肽及早地把信息传递到胰岛,引起胰岛素较早的分泌;使血糖不至于升得过高而从尿中丢失,这对于有效地保持机体所获得的能源,具有重要的生理意义。
影响其它激素释放的胃肠激素还有:生长抑素、胰多肽、血管活性肽等,它们对生长激素、胰岛素、胰高血糖素、胃沁素等的释放均有调节作用。
3.营养作用 一些胃肠激素具有刺激消化道组织的代谢和促进生长的作用,称为营养作用(trophic action)。例如,胃泌素能刺激胃沁酸部和粘膜和十二指肠粘膜的蛋白质、RNA和DNA的合成。从而促进其生长。给动物长期注射五肽胃泌素(一种人工合成的胃泌素,含有胃泌素活性的最小片段—羧基端的5个氨基酸片段),可引起壁细胞增生。在临床上也观察到,切除胃窦的病人,血清胃泌素水平下降,同时可发生胃粘膜萎缩;相反,在患有胃泌素瘤的病人,血清胃泌素水平很高,这种病人多有胃粘膜增生、肥厚。此外,近年来还发现,小肠粘膜内I细胞释放的胆囊收缩素也具有重要的营养作用,它能引起胰腺内DNA、RAN和蛋白质的合成增加,促进胰腺外分泌组织的生长。
由胃肠内分泌细胞释放的激素主要是通过血液循环运送到靶细胞起作用的,这些出现在血液中的激素,可用放射免疫方法从血液中测定出来。但有一些胃肠激素释放后并不进入血液循环,而是通过细胞外注弥散至邻近的靶细胞,这种传递局部信息的方式也称为旁分泌(paracrine)。由胃窦部或胰岛内的D细胞释放的生长抑素,很可能是以这种方式发挥其对邻近的胃泌素细胞(G细胞)或胰B细胞的抑制性调节作用的。图6-7是胃肠激素这两种作用方式的模式图。
图6-7 胃肠激素作用的两种主要方式
A:经典的内分泌方式 B和C:旁分泌方式
(三)脑-肠肽的概念
近年来的研究证实,一些产生胃肠道的肽,不仅存在于胃肠道,也存在于中枢神经系统内;而原来认为只存在于中枢神经系统的神经肽,也在消化道中发现。这些双重分布的肽被统称为脑-肠肽(braingut peptide)。已知的脑-肠肽有胃泌素、胆囊收缩素、P物质、生长抑素、神经降压素等约20余种。这些肽类双重分布的生理意义已引起人们的重视,例如胆囊收缩素在外周对胰酶分泌和胆汁排放的调节作用及其在中枢对摄食的抑制作用,提示脑内及胃肠内的胆囊收缩素在消化和吸收中具有协调作用。
第二节 口腔内消化
消化过程是从口腔开始的。食物在口腔内停留的时间很短,一般是15-20秒钟。食物在口腔内咀嚼,被唾液湿润而便于吞咽。由于唾液的作用,食物中的某些成分还在口腔内发生化学变化。
一、唾液分泌
人的口腔内有三对大的唾液腺:腮腺、颌下腺和舌下腺,还有无数散在的小唾腺。唾液就是由这些大小唾液腺分泌的混合液。腮腺是由浆液细胞组成的,分泌稀的唾液,;颌下腺和舌下腺是混合腺,即腺泡由浆液细胞和粘液细胞组成。
(一)唾液的性质和成分
唾液为无色无味近于中性(Ph6.6-7.1)的低渗液体。唾液中分约占99%。有机物主要为粘蛋白,还有球蛋白、氨基酸、尿素、尿酸、唾液淀粉酶和溶菌酶等。唾液中的无机物有钠、钾、钙、硫氰酸盐、氯、氨等。此外,唾液中还有一定量的气体,如氧、氮和二氧化碳。
唾液中的粘蛋白几乎全由粘液细胞所分泌,它使唾液具有粘稠性质。浆细胞分泌稀薄的唾液,几乎不含粘蛋白,但浆液腺所分泌的唾液淀粉酶是粘液腺所分泌的4倍。
唾液的渗透压随分泌率的变化而有所不同。在分泌率很低的情况下,其渗透压也低,约为50mOsm/kgH2O;而在最大分泌率时,渗透压可接近血浆,唾液中钠和氯的浓度升高,钾的浓度降低;分泌率低时则出现相反的现象。目前认为,唾液中电解质成分随分泌率变化的原因是分泌液在流经导管时,导管上皮细胞对电解质的吸收不相同而造成的,而分泌液从腺泡细胞中排出时是等渗的,电解质的组成与血浆是相似的。
(二)唾液的作用
唾液可以 湿润与溶解食物,以引起味觉并易于吞咽;唾液还可清洁和保护口腔,它可清除口腔中的残余食物,当有害物质进入口腔时,它可冲淡、中和这些物质,并将它们从口腔粘膜上洗掉,唾液中的溶菌酶还有杀菌作用;在人和少数哺乳动物如兔、鼠等的唾液中,含中唾液淀粉酶(狗、猫、马等的唾液中无此酶),它可使淀粉分解成为麦芽糖。唾液淀粉酶发挥作用的最适ph 在中性范围内,唾液中的氯和硫氰酸盐对此酶有激活作用。食物进入胃后,唾液淀粉酶还可继续使用一段时间,直至胃内容物变为pH约为4.5的酸性反应为止。
(三)唾液分泌的调节
唾液分泌的调节完全是神经反射性的,包括非条件反射和条件反射两种。
引起非条件反射性唾液分泌的正常刺激是食物对口腔机械的、化学的和温度的刺激。在这些刺激的影响下,口腔粘膜和舌的神经末稍(感受器)发生兴奋,冲动沿传入神经纤维(在舌神经、鼓索神经支、舌咽神经和迷走神经中)到达中枢,再由传出神经到唾液腺,引起唾液分泌(图6-8)。
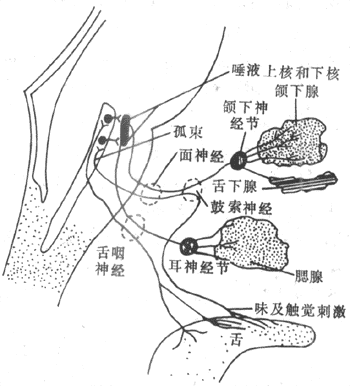
图6-8 唾液腺的神经支配
唾液分泌的初级中枢在延髓,其高级中枢分布于下丘脑和大脑皮层等处。
支配唾液腺的传出神经以副交感神经为主,如第9对脑神经到腮腺,第7对脑神经的鼓索支到颌下腺。刺激这些神经可引起量多而固体少的唾液分泌。副交感神经的对唾液腺的作用是通过其末稍释放乙酰胆碱而实现的,因此,用对抗乙酢胆碱的药物如阿托品,能抑制唾液分泌,而用乙酰胆碱或其类似药物时,可引起大量的唾液分泌。副交感神经兴奋时,还可使唾液腺的血管舒张,进一步促进唾液的分泌。目前认为,副交感神经引起唾液腺附近血管舒张的神经纤维是肽能神经纤维,其末稍释放血管活性肠肽。
支配唾液腺的交感神经是肽能神经纤维,在颈上神经节换神经元后,发出节后纤维分布在唾液腺的血管和分泌细胞上。刺激这些神经引起血管收缩,也可引起唾液分泌,但其分泌作用则随不同的唾液腺而有不同,例如,刺激人的颈交感神经,只引起颌下腺分泌,却不引起腮腺分泌。
人在进食时,食物的形状、颜色、气味,以及进食的环境,都能形成条件反射,引起唾液分泌。“望梅止渴”就是日常生活中条件反射性唾液分泌的一个例子。成年人的唾液分泌,通常都包括条件反射和非条件反射两种成分在内。
二、咀嚼
口腔通过咀嚼运动对食物进行机械性加工。咀嚼是由各咀嚼肌有顺序地收缩所组成的复杂的反射性动作。咀嚼肌包括咬肌、翼内肌、翼外肌和颞肌等,它们的收缩可使下颌向上、向下、向左右及向前方运动,这时,上牙列与下牙列相互接触,可以产生很大的压力以磨粹食物。咀嚼还使食物与唾液充分混合,以形成食团,便于吞咽。
咀嚼肌是骨骼肌,可作随意运动,但在正常情况下,它的运动还受口腔感受器和咀嚼肌内的本体感受器传来的冲动的制约。在咀嚼运动中,颊肌和舌肌的收缩具有重要作用,它们的收缩可将食物置于上下牙列之间,以便于咀嚼。
吸吮也是一个反射动作,吸吮时,口腔壁肌肉和舌肌收缩,使口腔内空气稀薄,压力降低到比大气压力为低0.98-1.47kPa(10-15cmH2O)。凭着口腔内的这个低压条件,液体便可进入口腔。
应当指出,口腔内消化过程不仅完成口腔内食物的机械性和化学性加工,它还能的反射性地引起胃、胰、肝、胆囊等的活动,以及引起胰岛素的分泌等等变化,为以后的消化过程及紧随消化过程的代谢过程,准备有利条件。
三、吞咽
吞咽是一种复杂的反射性动作,它使食团从口腔进入胃。根据食团在吞咽时所经过的部位,可将天咽动作分为下列三期:
第一期:由口腔到咽。这是在来自大脑皮层的冲动的影响下随意开始的。开始时舌尖上举及硬腭,然后主要由下颌舌骨肌的收缩,把食团推向软腭后方而至咽部。舌的运动对于这一期的吞咽动作是非常重要的。
第二期:由咽到食管上端。这是通过一系列急速的反射动作而实现的。由于食团刺激了软腭部的感受器,引起一系列肌肉的反射性收缩,结果使软腭上升,咽后壁向前突出,封闭了鼻回通路;声带内收,喉头升高并向并紧贴会厌,封闭了咽与气管的通路;呼吸暂时停止;由于喉头前移,食管上口张开,食团就从咽被挤入食管。这一期进行得极快,通常约需 0.1s。
第三期:沿食管下行至胃。这是由食管肌肉的顺序收缩而实现的。食管肌肉的顺序收缩又称蠕动(peristalsis),它是一种向前推进的波形运动。在食团的下端为一舒张波,上端为一收缩波,这样,食团就很自然地被推送前进(图6-9)。
图6-9 食管蠕动的模式图
食管的蠕动是一种反射动作。这是由于食团刺激了软腭、咽部和食管等处的感受器,发出传入冲动,抵达延髓中枢,再向食管发出传出冲动而引起的。
在食管和胃之间,虽然在解剖上并不存在括红肌,但用测压法可观察到,在食管也胃贲门连接处以上,有一段长约4-6cm的高压区,其内压力一般比胃0.67-1.33kPa(5-10mmHg),因此是正常情况下阻止胃内容物逆流入食管的屏障,起到了类似生理性括约肌作用,通常将这一食管称为食管-胃括约肌。当食物经过食管时,刺激食管壁上的机械感受器,可反射性地引起食管-胃括约肌舒张,食物便能进入胃内。食物入胃后引起的胃泌素释放,则可加强该括约肌的收缩,这对于防止胃内容物逆流入食管可能具有一定作用。
总之,吞咽是一种典型的、复杂的反射动作,它有一连串的按顺序发生的环节,每一环节由一系列的活动过程组成,前一环节的活动又可引起后一环节的活动。吞咽反射的传入神经包括来自软腭(第5、9对神脑经)、咽后壁(第9对脑神经)、会咽(第10对脑神经)、和食管(第10对脑神经)等外的脑神经的传入纤维。吞咽的基本中枢位于延髓内,支配舌、喉、咽部肌肉动作的传出神经在第5、9、12对脑神经中,支配食管的传出神经是迷走神经。
从吞咽开始至食物到达贲门所需的时间,与食物的性状及人体的体位有关。液体食物约需3-4s,糊状食物约5s,固体食物较慢,约需6-8s,一般不超过15s。
第三节 胃内消化
胃是消化道中最膨大的部分。成人的容量一般为1-2L,因而具有暂时贮存食物的功能。食物入胃后,还受到胃液的化学性消化和胃壁肌肉运动的机械性消化。
一、胃的分泌
胃粘膜是一个复杂的分泌器官,含有三咱管状外分泌腺和多种内分泌细胞。
胃的外分泌腺有:①贲门腺分布在胃与食管连接处的宽约1-4cm的环状区内,为粘液腺,分泌粘液;②泌酸腺分布在占全胃粘膜约2/3的胃底和胃体部。泌酸腺由三种细胞组成:壁细胞、主细胞和粘液颈细胞,它们分别分泌盐酸、胃蛋白酶原和粘液;③幽门腺分布在幽门部,是分泌碱性粘液的腺体。胃液是由这三种腺体和胃粘膜上皮细胞的分泌物构成的。
胃粘膜内至少含有6种内分泌细胞,如分泌胃泌素的G细胞、分泌生长抑素的D细胞和分泌组胺的肥大细胞等。
(一)胃液的性质、成分和作用
纯净的胃液是一种无色而呈酸性反应的液体,Pho 0.9-1.5。正常人每日分泌的胃液量约为1.5-2.5L。胃液的成分包括无机物如盐酸、钠和钾的氯化物等,以及有机物如粘蛋白、消化酶等。与唾液相似,胃液的成分也随分泌的速率而变化,当分泌率增加时,氢离子浓度升高,钠离子浓度下降,但氯和钾的浓度几乎保持恒定(图6-10)。
1.盐酸 胃液中的盐酸也称胃酸,其含量通常以单位时间内分泌的盐酸mmol表示,称为盐酸排出量。正常人空腹时盐酸排出量(基础酸排出量)约为0-5mmol/h。在食物或药物(胃泌素或组胺)的刺激下,盐酸排出量可进一步增加。正常人的盐酸最大排出量可达20-25mmol/h。男性的酸分泌多于女性;盐酸的排出量反映胃的分泌能力,它主要取决于壁细胞的数量(图6-11),但也与壁细胞的功能状态有关。
图6-10 人胃液中电解质浓度与分泌率的关系胃液分泌是用组胺静脉注射引起的
图6-11 胃酸最大排出量与壁细胞数目的关系
由17个人胃的切除部分计算出最大酸排出量和壁细胞数目的关系。
由图显示每100万个壁细胞可产酸约25mEp/h
胃液中H+的最大浓度可达150mmol/L,比血液中H+的浓度高三、四百万倍,因此,壁细胞分泌H+是逆着巨大的浓度梯度进行的,需要消耗大量的能量,能量来源于氧代谢。
泌酸所需的H+来自壁细胞浆内的水。水解离产生H+和OH-,任借存在于壁细胞上分泌小管膜上的H+、K+-ATP酶的作用,H+被主动地转运入小管腔内。
壁细胞分泌小管膜上的H+、K+-ATP酶又称质子泵(proton pump)或称酸泵。H+-K+交换是壁细胞质子泵区别于体内任何其它细胞上的质子泵的显著特征。H+、K+-ATP酶每催化一分子的ATP分解为ADP和磷酸所释放的能量,可驱动一个H+从壁细胞浆进入分泌小管腔和一个K+从小管腔进入细胞浆。H+的分泌必须在分泌小管内存在足够浓度的K+的条件下才能进行。
年来,选拔性干扰胃壁细胞的H+、K+-ATP酶的药物已被用来有效地抑制胃酸分泌,成为一代新型的抗溃疡药物。
已知壁细胞内含有丰富的碳酸酐酶,在它的催化下,由细胞代谢产生的CO2和由血浆中摄取的CO2可迅速地水合而形成H2CO3,H2CO3随即又解离为H+和HCO3。这样,在H+分泌后,留在细胞内的OH-便和由
H2CO3解离的H+结合而被中和,壁细胞内将不致因OH-的蓄积而使pH升高。由H2CO3产生的HCO3则在壁细胞的底侧膜,与CI-并换而进入血液。因此,餐后与大量胃酸分泌的同时,血和尿的pH往往升高而出现“餐后碱潮”。与HCO3交换而进入壁细胞内的CI-则通过分泌小管膜上特异性的CI-通道进入小管腔,与H+形成HCi (图6-12)。
图6-12 壁细胞分泌盐酸的一种假设
胃内的盐酸有许多作用,它可杀死随食物进入胃内的细菌,因而对维持胃和小肠内的无菌状态具有重要意义。盐酸还能激活胃蛋白酶原,使之转变为有活性的胃蛋白酶,盐酸并为胃蛋白酶作用提供了必要的酸性环境。盐酸进入小肠后,可以引起促胰液素的释放,从而促进胰液、胆汁和小肠液的分泌。盐酸所造成的酸性环境,还有助于小肠对铁和钙的吸收。但若盐酸分泌过多,也会对人体产生不利影响,。一般认为,过高的胃酸对胃和十二指肠粘膜有侵蚀作用,因而溃疡病发病的重要原因之一。
2.胃蛋白酶原 胃蛋白酶原是由主细胞合成的,并以不具有活性的酶原颗粒形式贮存在细胞内。当细胞内充满酶原颗粒时,它对新的酶原的全盛2产生负反馈作用。持续的刺激可使主细胞内的颗粒历释放而完全消失,便分泌仍继续进入,说明酶原也可以不经过颗粒的形式直接释放出来。
分泌入胃腔内的胃蛋白酶原在胃酸的作用下,从分子中分离出一个小分子的多肽,转变为具有活性的胃蛋白酶。已激活的胃蛋白酶对胃蛋白酶原也有激活作用。
胃蛋白酶能水解食物中的蛋白质,它主要作用于蛋白质及多肽分子中含苯丙氨酸或酪氨酸的肽键上,其主要分解产物是胨,产生多肽或氨基酸较少。胃蛋白酶只有在酸性较强的环境中才能发挥作用,其最知pH为2。随着pH的升高,胃蛋白酶的活性即降低,当pH升至6以上时,此酶即发生不可逆的变性。
3.粘液和碳酸氢盐 胃的粘液是由表面上皮细胞、泌酸腺的粘液颈细胞,贲门腺和幽门腺共同分泌的,其主要成分为糖蛋白。糖蛋白是由4个亚单位通过二硫键连接形成的(图6-13)。由于糖蛋白的结构特点,粘液具有较高的粘滞性和形成凝胶的特性。在正常人,粘液覆盖在胃粘膜的表面,形成一个厚约500μm的凝胶层,它具有润滑作用,可减少粗糙的食物对胃粘膜的机械性损伤。
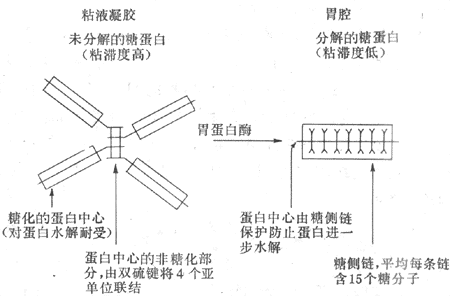
图6-13 胃粘液糖蛋白结构的示意图
胃内HCO3主要是由胃粘膜的非泌酸细胞分泌的,仅有少量的HCO3是从组织间液渗入胃内的。基础状态下,胃HCO3分泌的速率仅为H+分泌速率的5%。进食时其分泌速率的增加通常是与H+分泌速率的变化平行的。由于H+和HCO3在分泌速率和浓度上的巨大差距,分泌的HCO3对办内pH显然不会有多大影响。
长期以来人们一直在思索:胃粘膜处于高酸和胃蛋白酶的环境中,为什么不被消化?近年来“粘液-碳酸氢盐屏障”概念的提出,至少部分地回答了这个问题。这主要是因为。胃粘液的粘稠度约为水的30-260倍,H+和HCO3等离子在粘液层内的扩散速度明显减慢,因此,在胃腔同内的H+向粘液凝胶深层弥散过程中,它不断地与从粘液层下面的上皮细胞分泌并向表面扩散的HCO3遭遇,两种离子在粘液层内发生中和。用pH测量电极测得,在胃粘液层存在一个pH梯度,粘液层靠近胃腔面的一侧呈酸性,pH为7左右(图6-14)。因此,由沾液和碳酸氢盐共同构筑的粘液-碳酸氢盐屏障。能有效地阻挡H+的逆向弥散,保护了胃粘液免受H+的假侵蚀;粘液深层的中性pH环境还使胃蛋白酶丧失了分解蛋白质的作用。
图6-14 胃粘液-碳酸氢盐屏障模式图
正常情况下,胃粘液凝胶层临近胃腔一侧的糖蛋白容易受到胃蛋白酶的作用而水解为4个亚单位,这样,粘液便从凝胶状态变为溶胶状态而进入胃液。但一般来讲,水解的速度与粘膜上皮细胞分泌液的速度相等,这种粘液分泌与降解裼动态平衡,保持了粘液屏障功能的完整性和连续性。
4.内因子泌酸腺的壁细胞除分泌盐酸外,还分泌一种分子量在50000-60000之间的糖蛋白,称为内因子。内因子可与进入胃内的维生素B12结合而促进其吸收。
(二)胃液分泌的调节
胃液分泌受许多因素的影响,其中有的起兴奋性作用,有的则起抑制性作用。进食是胃液分泌的自然刺激物,它通过神经和体液因素调节胃液的分泌。
1.刺激胃酸分泌的内源性物质
(1)乙酰胆碱:大部分支配胃的副交感神经节后纤维末稍释放乙酰胆碱。乙酰胆碱直接作用于壁细胞膜上的胆碱能受体,引起盐酸分泌增加。乙酰胆碱的作用可被胆碱能受体阻断剂(如阿托品)阻断。
(2)胃泌素:胃泌素主要由胃窦粘膜内的G细胞分泌。十二指肠和空肠上段粘膜内也有少量G细胞。胃泌素释放后主要通过血液循环作用于壁细胞,刺激其分泌盐酸。
胃泌素以多种分子形式存在于体内,其主要的分子形式有两种:大胃泌素(G-34)和小胃泌素(G-17)。胃窦粘膜内的胃泌素主要是G-17,十二指肠粘膜中有G-17和G-34约各占一半。从生物效应来看,G-17刺激胃分泌的作用要比G-34强5-6倍,但G-34在体内被清除的速度很慢,它拦衰期约为50min,而G-17通常只有6min。
图6-15 胃泌素的氨基酸序列
划线的部分是其活性片段
人的小胃泌素的氨基酸序列如图6-15,其C端正的4个氨基酸是胃泌素的最小活性片段,因此,用人工合成的四肽或五肽胃泌素是具有天然胃泌全部作用的人工制品。
(三)组胺:胃的泌酸区
粘膜内含有大量的组胺。产生组胺的细胞是存在于固有膜中的肥大细胞。正常情况下,胃粘膜恒定地释放少量组胺,通过局部弥散到达邻近的壁细胞,刺激其分泌。壁细胞上的组胺受体为Ⅱ型受体(H2受体),用甲氰咪呱(cimetidine)及其相类似的药物可以阻断组胺与壁细胞的结合,从而减少胃酸分泌。
以上三种内源性分泌物,一方面可通过各自壁细胞上的特异性受体,独立地发挥刺激胃酸分泌的作用(图6-16);另一方面,三者又相互影响,表现为当以上三个因素中的两个因素同时作用时,胃酸的分泌反应往往比这两个因素单独作用的总和要大,这种现象在生理学上称为加强作用(potentiation)。在整体内,促分泌物之间的相互加强作用是经常存在的,因此,用任何一咱促分泌物的阻断剂,如用甲氰咪呱时,它不仅抑制了壁细胞对组胺的反应,同时也会由于去除了组胺的作用的背景,使壁细胞对胃泌素和乙酰胆碱的反应也有所降低。
图6-16 三种刺激胃酸分泌的内源性物质的作用及相互关系
A:阿托品 CM:甲氰咪呱
2.消化期的胃液分泌 进食后胃液分泌的机制,一般按接受食物刺激的部位,分成三个时期来分析,即头期、胃期和肠期。但必须注意,三个时期的划分是人为的,只是为了便于叙述,实际上,这三个时期几乎是同时开始的、相互重叠的。
(1)头期胃液分泌:头期的胃液分泌是由进食动作引起的,因其传入冲动均来自头部感受器(眼、耳、口腔、咽、食管等)。因而称为头期。
头期胃液分泌的机制曾用慢性方法作了较详细的分析,即用事先旅行过食管切断术并具有胃瘘的狗进行假饲(sham feeding);当食物经过口腔进入食管后,随即从食管的切品流出体外,食物并未进入胃内,但却引起胃液分泌。进一步分析确定,由进食动作所引起的胃液分泌,包括条件反射性和非条例反射性两种分泌。前者是由和食物有关的形象、气味、声音等刺激了视、嗅、听等感受器而引起的;后者则当咀嚼和吞咽食物时,刺激了口腔和咽喉等外的化学和机械感受器而引起的。这些反射的传入途径和由进食引起的唾液分泌的传入途径相同,反射中枢包括延髓、下丘脑、边缘和大脑皮层等。迷走神经是这些反射共同的传出神经。当切断支配胃的迷走神经后,假饲就不再引起胃液分泌。
迷走神经兴奋后,除了通过其末稍释放乙酰胆碱,直接引起腺体细胞分泌外,迷走神经冲动还可引起胃窦粘膜内的G细胞释放胃泌素,后者经过血液循环刺激胃腺颁发(图6-17)。由此可见,头期的胃注分泌并不是纯神经反射性的,而是一种神经-体液性的调节。
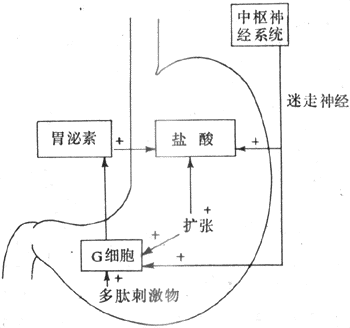
图6-17 引起胃酸分泌的机制
引起胃泌素释放的迷走神经纤维被认为是非胆碱能的,因为阿托品不仅不能阻断,反而使假饲引起的胃泌素释放反应增加。目前对这一现象的解释是,迷走神经中既存在兴奋胃泌素释放的纤维,也存在抑制胃泌素释放的纤维,前者的中介物可能是一种肽类物质,而抑制性纤维则是通过乙酰胆碱能起作用的。阿托品由于阻断了抑制性纤维的作用,因而使胃泌素的释放有所增加。
头期胃液分泌的量和酸度都很高,而胃蛋白酶的含量则尤其高。在人体观察的资料表明,头期胃液分泌的大小与食欲有很大的关系。
(2)胃期胃液分泌:食物入胃后,对胃产生机械性和化学性刺激,继续引起胃液分泌,其主要途径为:①扩张刺激胃底、胃体部的感受器,通过迷走、迷走神经经长反射和壁内神经丛的短反射,引起胃腺分泌;②扩张刺激胃幽门部,通过壁内神经丛,作用于G细胞,引起胃泌素的释放;③食物的化学成分直接作用于G细胞,引起胃泌素的释放。
刺激G细胞释放胃泌素的主要食物化学成分是蛋白质的消化产物,其中包括肽类和氨基酸。G细胞为开放型胃肠内分泌细胞,顶端有绒毛样突起伸入胃腔,可以直接感受胃腔内化学物质的作用。用放射免疫方法测定血浆中胃泌素浓度,正常人空腹时约为30-120pg/ml,在进食蛋白质食物后,血浆胃泌素可升高到50-200pg/ml,在食后2-3小时逐渐恢复至进食前水平。糖类和脂肪类食物不是胃泌素释放的强刺激物。
胃酸分泌的胃液酸度也很高,但胃蛋白酶含量却比头期分泌的胃液为弱。
(3)肠期胃液分泌:将食糜、内的提取液、蛋白胨液由瘘管直接注入十二指肠内,也可引起胃液分泌的轻度增加,说明当食物离开胃进入小肠后,还有继续刺激胃液分泌的作用。机械扩张游离的空肠袢,胃液分泌也增加。
在切断支配胃的外来神经后,食物对小肠的作用仍可引起胃液分泌,提示肠期胃液分泌的机制中,神经反射的作用不大,它主要通过体液调节机制,即当食物与小肠粘膜接触时,有一种或几种激素从小肠粘膜释放出来,通过血液循环作用于胃。已知人的十二指肠粘膜中含有较多的胃泌素;用放射免疫方法测得,在切除了胃窦的病人,进食后血浆胃泌素的浓度仍有升高,说明进食后可引起十二指肠释放胃泌素,它可能是肠期胃泌分泌的体液因素之一。有人认为,在食糜作用下,小肠粘膜还可能释放肠泌酸素的激素,刺激胃酸分泌。此外,由小肠吸收的氨基酸也可能参与肠期的胃液分泌,因为静脉注射混合氨基酸也可引起胃酸分泌。
肠期胃液分泌的量不大,大约占进食后胃液分泌总量的1/10,这可能与食物在小肠内同时还产生许多对胃液起抑制性作用的调节(见后文)有关。
3.胃液分泌的抑制性调节上已述在进食过程中兴奋胃液分泌的机制,而正常消化期的胃液分泌还受到各种抑制性因素的调节,实际表现的胃液分泌正是兴奋和抑制性因素共同作用的结果。在消化期同内,抑制胃液分泌的因素除精神、情绪因素外,主要在盐酸、脂肪和高张溶液三种。
盐酸是胃腺活动的产物,但它对胃腺的活动又具有抑制性作用,因此是胃酸分泌的一种负反馈的调节机制。
当胃窦的pH降到1.2-1.5时,便可能对胃液分泌的产生抑制作用。这种抑制作用的机制可能是盐酸直接抑制了胃窦粘膜中的G细胞,减少胃泌素释放的结果。恶性贫血的病人胃酸分泌很低,他们血浆中胃泌素的浓度却比正常人高20-30倍,如向这种病人胃内注以盐酸,使胃内酸化,血浆胃泌素的浓度即下降,这进一步说明,胃内容物的酸度对胃泌素的释放,以及进而影响胃液分泌具有重要作用。
近年来一些实验资料提示,盐酸在胃内还可能通过引起胃粘膜释放一种抑制性因素——生长抑素,转而抑制胃泌素和胃液的分泌。
当十二接指肠内的pH降到2.5以下时,对胃酸分泌也有抑制作用,但其作用机制目前尚未完全阐明。已知酸作用于小肠粘膜可引起促胰液素释放,后者对胃泌素引起的酸分泌具有明显的抑制作用,因此,促胰液素很可能是十二指肠酸化抑制胃分泌的一种抑制物。此外,十二指肠球部在盐酸刺激下,也可能释放出一种抑制胃分泌的肽类激素——球抑胃素,但球抑胃素结构尚未最后确定。
脂肪是抑制胃液分泌的一个重要因素。脂肪及其消化产物抑制胃分泌的作用发生在脂肪进入十二指肠后,而不是在胃中。早在30年代,我国生理学家林可胜就发现,从小肠粘膜中可提取出一种物质,当由静脉注射后,保使胃液分泌的量、酸度和消化力减低,并抑制胃运动。这个物质被认为是脂肪在小肠内抑制胃分泌的体液因素,而可能是几种具有此种作用的激素的总称。小肠粘膜中存在的抑胃肽、神经降压素等多种激素,都具有类似肠抑素的特性。
十二指肠内高张溶液对胃分泌的抑制作用可能通过两种途径来实现,即汽船活小肠内渗透压感受器,通过肠-胃反射引起胃酸分泌的抑制,以及通过刺激小肠粘膜释放一种或几种抑制性激素而抑制胃液分泌。后一机制尚未阐明。
在胃的粘膜和肌层中,存在大量的前列腺素(详见内分泌章)。迷走神经兴奋和胃泌素都可引起前列腺素释放的增加。前列腺素对进食、组胺和胃泌素等引起的胃液分泌有明显的抑制作用。它可能是胃液分泌的负反馈抑制物。前列腺素还能减少胃粘膜血流,但它抑制胃分泌的作用并非继发于血流的改变。
二、胃的运动
胃既有贮存食物的功能,又具有泵的功能。胃低和胃体的前部(也称头区)运动较弱,其主要功能是贮存食物;胃体的远端和胃窦(也称尾区)则有较明显的运动,其要主功能是磨粹食物、使食物与胃液充分混合,以形成食糜,以及逐步地将食糜排至十二指肠。
(一)胃的容受性舒张
当咀嚼和吞咽时,食物对回、食管等外感受器的刺激,可通过迷走神经反射性地引起胃底和胃体贴骨肉的舒张。胃壁肌肉的这种活动,被称为胃的容受性舒张。容受性舒张使胃腔容量由空腹时的50ml,增加到进食后的1.5L,它适应于大量食物的涌入,而胃内压力变化并不大,从而使胃更好地完成容受和贮存食物的功能,其生理意义是显然的。
胃的容受性舒张是通过迷走神经的传入和传出通路反射地实现的,切断人和动物的双侧迷走神经,容受性舒张即不再出现。在这个反射中,迷走神经和传也通路是抑制性纤维,其末稍释放的递质既非乙酰胆碱,也非去甲肾上腺素,而可能是某种肽类物质。
(二)胃的蠕动
食物进入胃后约5分钟,蠕动即开始。蠕动是从胃的中部开始,有节律地向幽门方向进行。在人,胃蠕动波的频率约每分钟3次,并需1分钟左右到达幽门。因此,通常是一波未平,一波又起。
蠕动波在初起时比较小,在向幽站传播过程中,波的深度和速度都逐步增加,当接近幽站时,明显加强,可将一部分食糜(约1-2ml)排入十二指肠,因此有幽站泵之称。并不是每一个蠕动波都到达幽门,有些蠕动波到胃窦后即行消失。一旦收缩波超越胃内容物,并到达胃窦终末时,由于胃窦终末部的有力收缩,胃内容物部分将被反向地推回到近侧胃窦和胃体部(图6-18)。食糜的这种后退,非常有利于食物和消化液的混合,还可机械地磨碎块状固体食物。总之,蠕动主要的生理意义是:一方面使食物与胃液充分混合,以利于胃液发挥消化作用;另一方面,则可搅拌和粉碎食物,并推进胃内容物通过幽门向十二指肠称行。
图6-18 胃的蠕动
胃的蠕动是受胃不滑肌的基本电节律控制的。胃的基本电节律起源于胃大弯上部,沿纵行肌向幽门方向传播,每分钟约3次。胃肌的收缩通常出现在基本电节律波后6-9s,动作电位后1-2s。神经和体液因素可通过影响胃的基本电节律和动作电位而影响胃的蠕动;迷走神经冲动、胃泌素和胃动素(是近年来从小肠粘膜中分离出来的一种胃肠激素)可使胃的基本电节律和动作电位出现的频率增加,使胃的收缩频率和强度增加;交感神经兴奋、促胰液素和抑胃肽则作用相反。
(三)胃的排空及其控制
食物由胃排入十二指肠的过程称为胃的排空。一般在食物入胃后5分钟即有部分食糜被排入十二指肠。不同食物的排空速度不同,这和食物的物理性状和化学组成都有关系。稀的、流体食物比稠的或固体食物排空快;切碎的、颗粒小的食物比大块的食物排空快;等涌液体比非等渗液体快。在三种主要食物中,糖类的排空时间较蛋白质为短,脂肪类食物排空最慢。对于混合食物,由胃完全排空通常需要4-6小时。
胃的排空率受来处胃和来自十二指肠二方面因素的控制:
1.胃内因素促进排空
(1)胃内食物量对排空率的影响:胃的内容物作为扩张胃的机械刺激,通过壁内神经反射或迷走-迷走神经反射,引起胃运动的加强。一般,食物由胃排空的速率和留在胃内作物量的平方根成正比。
(2)胃泌素对胃排空的影响:扩张刺激以及食物的某些成分,主要是蛋白质消化产物,可引起胃窦粘膜释放胃泌素。胃泌素除了胃酸分泌外,对胃的运动也有中等程度的刺激作用,它提高幽站泵的活动,便使幽站舒张,因而对胃拓空有重要的促进作用。
2.十二指肠因素抑制排空
(1)肠-胃反射对胃运动的抑制:在十二指肠壁上存在多种感受器,酸、脂肪、渗透压及机械扩张,都可刺激这些感受器,反射性地抑制胃运动,引起胃排空减慢。这个反射称为肠-胃反射,其传出冲动可通过迷走神经、壁内神经,甚至还可能通过交感神经等几条途径传到胃。肠-胃反射对酸的刺激特别敏感,当pH降到3.5-4.0时,反射即可引起,它抑制幽门泵的活动,从而阻止酸性食糜进入十二指肠。
(2)十二指肠产生的激素对胃排空的抑制:当过量的食糜,特别是酸或脂肪由胃进入十二指肠后,可引起不小粘膜释放几种不同的激素,抑制胃的运动,延缓胃的排空。促胰液素、抑胃肽等都具有这种作用,统称为肠抑胃素。
上述在十二指肠内具有抑制胃运动的各项因素产东是经常存在的,随着盐酸在肠内被中和,食物消化产物的被吸收,它们对胃的抑制性影响便渐渐消失,胃运动又逐渐增强,因而又推送另一部分食糜进入十二指肠。如此重复,使胃排内容物的排空较好地适应十二指肠内消化入吸收速度。
(四)呕吐
呕吐是将胃及肠内容物从口腔强力驱出的动作。机械的和化学的刺激作用于知根、咽部、胃、大小肠、总胆管、泌尿生殖器官等处的感受器,都可以引起呕吐。视觉和内耳庭的位置感觉发生改变时,也可引起呕吐。
呕吐前常出现恶习、流涎、呼吸急迫和心跳快而不规则等自主神经兴奋的症状。呕吐开始时,先是深吸气,声站紧闭,随着胃和食管下湍舒张,膈肌和腹肌猛烈地收缩,压挤胃的内容物通过食管而进入口腔。呕吐时,十二指肠和空肠上段也变得强烈起来,蠕动增快,并可转为痉挛。由于胃舒张而十二指肠收缩,平时的压力差转倒转,使十二指肠内容物倒流入胃,因此,呕吐物中常混有胆汁和小肠液。
在呕吐动作中,所有的这些活动都是反射性的。传入冲动的是由迷走神经和交感神经的感觉纤维、舌咽神经有及其它神经传入至延髓内的呕吐中枢。由中枢发出的冲动则沿迷走神经、交感神经、膈神经和脊神经等传到胃、小肠、膈肌和腹壁肌等处。呕吐中枢的位置在延髓外侧网状结构的背外侧缘。颅内压增高(脑水肿、肿瘤等情况)可直接刺激该中枢而引起呕吐。呕吐中枢在结构上和功能上与呼吸中枢、心血管中枢均有密切联系,它以协调这些邻近中枢的活动,从而在呕吐时产生复杂的反应。
在延髓呕吐中枢 的附近存在一个特殊的化学感受野,某些中枢性催吐药如阿朴吗啡,实际上是刺激了这个化学感受野,通过它再兴奋呕吐中枢的。
呕吐是一种具有保持意义的防御反射,它可把胃内有害的物质排出。但长期剧烈的呕吐会影响进食和正常消化活动,并且使大量的消化液丢失,造成体内水电解质和酸碱平衡的紊乱。
第四节 小肠内消化
食糜由胃进入十二指肠后,即开始了小肠内的消化。小肠内消化是整个消化过程中最重要的阶段。在这里,食糜受到胰液、胆汁和小肠液的化学性消化以及小肠运动的机械性消化。许多营养物质也都在这一部位被吸收入机体。因此,食物通过小肠,消化过程基本完成。未被消化的食物残渣,从小肠进入大肠。
食物在小肠内停留的时间,随食物的性质而有不同,一般为3-8h。
一、胰液的分泌
胰液是兼有外分泌和内分泌功能的腺体。胰腺的内分泌功能主要与糖代谢的调节有关,将在内分泌章中讨论。胰腺的外分泌为胰液,是由胰腺的腺泡细胞和小的导管管壁细胞所分泌的,具有很强的消化能力。
(一)胰液的成分和作用
胰液是列色无嗅的碱性液体,pH约为7.8-8.4,渗透压约为血浆相等。人每日分泌的胰液量约为1-2L。
胰液中含有无机物和有机物。在无机成分中,碳酸氢盐的含量很高,它是由胰腺内的小的导管细胞分泌的。导管细胞内含有较高浓度的碳酸酐酶,在它的催化下,二氧化碳可水化而产生碳酸,后者经过解离而产生碳酸氢根(HCO3),人胰液中的HCO3的最高浓度为140mmol/L,其浓度随分泌速度的增加而增加(图6-19)。HCO3的主要作用是中和进入十二指肠的胃酸,使肠粘膜免受强酸的侵蚀;同时也提供了小肠内多种消化酶活动的最适宜的pH环境(pH 7-8)。除HCO3外,占第二位的主要负离子是CI-。CI-的浓度随HCO3的浓度的变化而有变化,当HCO3浓度升高时,CI-的浓度就下降。胰液中的正离子有Na+、K+、Ca2+等,它们在胰液中的浓度与血浆中的浓度非常接近,不依赖于分泌的速度。
图6-19 胰液中电解质成分和分泌率的关系
胰液中的有机物主要是蛋白质,含量由0.1%-10%不等,随分泌的速度不同而有不同。胰液中的蛋白质主要由多种消化酶组成,它们是由腺泡细胞分泌的。胰液中的消化酶主要有:
1.胰淀粉酶 胰淀粉酶是一种α-淀粉酶,它对生的或熟的淀粉的水解效率都很高,消化产物为糊精、麦芽糖。胰淀粉酶作用的最知pH为6.7-7.0。
2.胰脂肪酶 胰脂肪酶可分解甘油三酯为脂肪酸、甘油一酯和甘油。它的最适pH为7.5-8.5。
目前认为,胰脂肪酶只有在胰腺分泌的另一种小分子 蛋白质——辅脂酶存在条件下才能发挥作用。胰脂肪酶与辅脂酶在甘油三酯的表面形成一种高亲和度的复合物,牢固地附着在脂肪颗粒表面,防止胆盐把脂肪酶从脂肪表面置换下来。因此,辅脂酶的作用可比喻为附着在甘油三酯表面的“锚”。
胰液中还含有一定量的胆固醇和磷脂酶A2,它们分别水解胆固醇酯和卵磷脂。
3. 胰蛋白酶和糜蛋白酶 这两种酶者是以不具有活性的酶原形式存在于胰液中的。肠液中的肠致活酶可以激活蛋白酶原,使之变为具有活性的胰蛋白酶。此外,酸、胰蛋白酶本身,以及组织液也能使胰蛋白酶原活化。糜蛋白酶原是在胰蛋白酶作用下转化为有活性的糜蛋白酶的。
胰蛋白酶和糜蛋白酶的作用极相似,都能分解蛋白质为胨,当两者一同作用于蛋白质时,则可消化蛋白质为小分子的多肽和氨基酸。
正常胰液中还含有羧基肽酶、核糖核酸酶、脱氧核糖核酸酶等水解酶。羧基肽酶可作用于多肽末端的肽键,释放出具有自由羧基的氨基酸,后两种酶则可使相应的核酸部分地水解为单核苷酸。
由于胰液中含有水解三种主要食物的消化酶,因而是所有消化液中最重要的一种。临床和实验均证明,当胰液分泌障碍时,即使其它消化腺的分泌都正常,食物中的脂肪和蛋白质仍不能完全消化,从而也影响吸收,但糖的消化和吸收一般不受影响。
(二)胰液分泌的调节
在非消化期,胰液几乎是不分泌或很少分泌的。进食开始后,胰液分泌即开始。所以,食物是兴奋胰腺的自然因素。进食时胰液受神经和体液双重控制,但以体液调节为主。
1.神经调节 食物的形象、气味、食物对口腔、食管、胃和小肠的刺激,都可通过神经反射(包括条件反射和非条件反射)引起胰液分泌。反射的传出神经主要是迷走神经。切断迷走神经,或注射阿托品阻断迷走神经的作用,都可显着地减少胰液分泌。迷走神经可通过其末稍释放乙酰胆碱直接作用于胰腺,也可通过引起胃泌素的释放,间接地引起胰腺分泌(图6-20)。迷走神经主要作用于胰腺的腺泡细胞,对导管细胞的作用较弱,因此,迷走神经兴奋引起胰液分泌的特点是:水分和碳酸氢盐含量很少,而酶的含量却很丰富。
内脏大神经对胰液分泌的影响不明显。内脏大神经中的胆碱能纤维可增加胰液分泌,但其上腺素能纤维则因使胰腺血管收缩,对胰液分泌产生抑制作用。
2.体液调节 调节胰液分泌的体液因素主要有促胰液素和胆囊收缩素(也称促胰酶素)两种,分述如下:
(1)促胰液素:当酸性食糜进入小肠后,可刺激小肠粘膜释放促胰液素。小肠上段粘膜含促胰液素较多,距幽门越远,含量越小。产生促胰液素的细胞为S细胞。王志均等曾在具有移植胰的狗身上观察引起促胰液素释放的因素,结果表明,盐酸是最强制刺激因素,其次为蛋白质分解产物和脂酸钠,糖类几乎无没有作用。小肠内促胰液素释放的pH阈值为4.5。迷走神经的兴奋不引起起促胰液素的释放;切除小肠的外来神经,盐酸在小肠内仍能引起胰液分泌,说明促胰液素的释放不依赖于肠管外来神经。
促胰液素主要作用于胰腺小导管的上皮细胞,使其分泌大量的水分和碳酸氢盐,因而使胰液的分泌量大为增加,便酶的含量却很低。
(2)胆囊收缩素:这是小肠粘膜中I细胞释放的一种肽类激素。引起胆囊收缩素释放的因素(由强至弱)为:蛋白质分解产物、脂酸钠、盐酸、脂肪。糖类没有作用。
促进胰液中各种酶的分泌是胆囊收缩素的一个重要作用,因而也称促胰酶素;它的另一重要作用是促进胆囊强烈收缩,排出胆汁。胆囊收缩素对胰腺组织还有营养作用,它促进胰组织蛋白质和核糖核酸的合成。
影响胰液分泌的体液因素还有胃窦分泌的胃泌素、小肠分泌的血管活性肠肽等,它们在作用分别与胆囊的收缩素和促胰液素相似。
近年来的资料表明,促胰液素和胆囊收缩素对胰液分泌的作用是通过不同机制实现的,前者以cAMP为第二信使,后者则通过磷脂酰醇系统,在Ca2+介导下起作用的(图6-20)。
促胰液素和胆囊收缩素之间具有协同作用,即一个激素可加强另一个激素的作用。此外,迷走神经对促胰液素的作用也有加强作用,例如阻断迷走神经后,促胰液素引起的胰液分泌量将大大减少。激素之间,以及激素与神经之间的相互加强作用,对进餐时胰液的大量分泌具有重要意义。
二、胆汁的分泌与排出
胆汁是由肝细胞不断生成的,生成后肝管流出,经胆总管而至十二指肠,或由肝管转入胆囊而存贮于胆囊,当消化时再由胆囊排出至十二指肠。胆汁和胰液、肠液一起,对小肠内的食糜进行化学性消化。
图6-20 胰液分泌的神经体液调节 实线代表水样分泌 虚线代表酶的分泌
(一)胆汁的性质和成分
成年人每日分泌胆汁约800—1000ml ,胆汁的生成量和蛋白质的摄入量有关,高蛋白食物可生成较多的胆汁。
胆汁是一种较浓的具有苦味的有色液汁。人的胆汁(由肝直接分泌的胆汁)呈金黄色或橘棕色;而胆囊胆汁(在胆囊中贮存过的胆汁)则因浓缩而颜色变深。肝胆汁呈弱碱性(pH为7.4),胆囊胆汁则因碳酸氢盐在胆囊中被吸收而呈弱酸性(Ph6.8)。
胆汁的成分很复杂,除水分和钠、钾、钙、碳酸氢盐等无机成分外,其有机成分有胆盐、胆色素、脂肪酸、胆固醇、卵磷脂和粘蛋白等。胆汁中没有消化酶。
胆盐是肝细胞分泌的胆汁酸与甘氨酸或牛磺酸结合形成的钠盐或钾盐,它是胆汁参与消化和吸收的主要成分。胆汁中的胆色素是血红蛋白的分解产物,包括胆红素复写纸的氧化物——胆绿质。胆色素的种类和浓度决定了胆汁的颜色,肝能合成胆固醇,其中约一半转化成胆汁酸。其余的一半则随胆汁进入胆囊或排入小肠。
在正常情况下,胆汁中的胆盐(或胆汁酸)、胆固醇和卵磷脂的适当比例是维持胆固醇成溶解状态的必要条件。当胆固醇分泌过多,或胆盐、卵磷脂合成减少时,胆固醇就容易测沉积下来,这是形成胆石的一种原因。
(二)胆汁的作用
胆汁对于脂肪的消化和吸收具有重要意义:
1.胆汁中的胆盐、胆固醇和卵磷脂等都可作为乳化剂,减低脂肪的表面张张,使脂肪乳化成微滴,分散在肠腔内,这样便增加了胰脂肪酶的作用面积,使其分解脂肪的作用加速。
2.胆盐因其分子结构的特点,当达到一定浓度后,可聚合而形成微胶粒。肠腔中脂肪的分解产物,如脂肪酸、甘油一酯等均可掺入到微胶中,形成水溶性复合物(混合微胶粒)。因此,胆盐便成了不溶于水的脂肪水解产物到达肠粘膜表面所必需的运载工具,对于脂肪消化产物的吸收具有重要意义。
3.胆汁通过促进脂肪分解产物的吸收,对脂溶性维生素(维生素A、D、E、K)的吸收也有促进作用。
此外,胆汁在十二指肠中还可以中和一部分胃酸;胆盐在小肠内吸收后还是促进胆汁自身分泌的一个体液因素。
(三)胆汁分泌和排出的调节
肝细胞是不断分泌胆汁的,但在非消化期间,肝胆汁都流入胆囊内贮存。胆囊可以吸收胆汁中的水分一无机盐,使肝胆汁浓缩4-10倍,从而增加了贮存的效能。在消化期,胆汁可直接由肝以及由胆囊中大量排出至十二指肠。因此,食物在消化道内是引起胆汁分泌和排出的自然刺激物。高蛋白食物(蛋黄、肉、肝)引起胆汁流出最多,高脂肪或混合食物的作用次之,而糖类食物的作用最小。在胆汁排出过程中,胆囊和Oddi括约肌的活动通常表现出协调的关系,即胆囊收缩时,Oddi括约肌舒张;相反,胆囊舒张时,Oddi括约肌则收缩。
1.神经因素的作用 神经对胆汁分泌和胆囊 收缩的作用均较弱。进食动作或食物对胃、小肠的刺激可通过神经反射引起肝胆汁分泌的少量增加,胆囊收缩也轻度加强。反射的传出途径是迷走神经,切断两侧迷走神经,或应用胆碱能受体阻断剂,均可阻断这种反应。
迷走神经除了直接作用于肝细胞和胆囊外,它还可通过引起胃泌素释放而间接引起肝胆汁的分泌和胆囊收缩。
2.体液因素的作用 有多种体注因素参与调节胆汁的分泌和排出。
(1)胃泌素:胃泌至少对肝胆的分泌及胆囊平滑肌的收缩均有一定的刺激作用,它可通过血液循环作用于肝细胞和胆囊;也可先引起胃酸分泌,后者再作用于十二指肠粘膜,引起促胰液素释放而促进肝胆汁分泌。
(2)促胰液素: 促胰液素主要的作用是刺激胰液分泌,但它还有一定的刺激肝胆汁分泌的作用。促胰液素主要作用于胆管系统而非作用于肝细胞,它引起的胆汁分泌主要是量和HCO3含量的增加,胆盐的分泌并不增加。
(3)胆囊收缩素:在蛋白质分解产物、盐酸和脂肪等物质作用下,小肠上部粘膜内的I细胞可释放胆囊收缩素,它通过血液循环兴奋胆囊平滑肌,引起胆囊的强烈收缩。胆囊收缩素对Oddi括约肌则有降低其紧张性的作用,因此可促使胆囊汁的大量排放。
胆囊收缩素也能刺激胆管上皮细胞,使胆汁流量和HCO3的分泌增加,但其作用较弱。
(4)胆盐:胆汁中的胆盐或胆汁酸当排至小肠后,绝大部分(约90%以上)仍可由小肠(主要为回肠末端)粘膜吸收入血,通过门静静脉回到肝,再组成胆汁而又分泌入肠,这一过程称为胆盐的肠肝循环(图6-21)。胆盐每循环一次约损失5%,每次进餐后约6-8g胆盐排出。每次进餐后可进行2-3次肠肝循环。返回到肝的胆盐有刺激肝胆汁分泌的作用,实验证明,当胆盐通过胆瘘流失至体外后,胆汁的分泌将比正常时减少数倍。
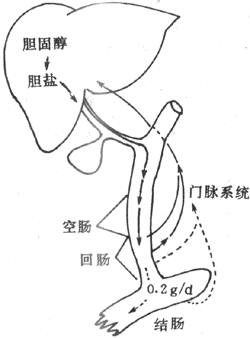
图 6-21 胆盐的肠-肝循环
进入门脉的实线代表来自肝的胆盐,虚线代表由细菌作用产生的胆盐胆盐对胆囊的运动并无影响。
总之,由进食开始,到食物进入小肠内,在神经和体液因素调节下,都可引起胆汁的分泌和排出活动,尤以食物进入小肠后的作用最为明显。在这一时期中,不仅肝胆汁的分泌明显增加,而且由于胆囊的强烈收缩,使贮存在胆囊中的胆汁也大量排出。
三、小肠液的分泌
小肠内有两种腺体:十二指肠和肠腺。十二指肠又称勃氏腺(Brunner’sgland),分布在十二指肠的粘膜下层中,分泌碱性液体,内含粘蛋白,因而粘稠度很高。这种分泌物的主要机能是保持十二指肠的上皮,不被胃酸侵蚀。肠腺又称李氏腺(Lieberkühn crypt),分布于全部小肠的粘膜层内,其分泌液构成了小肠液的主要部分。
(一)小肠液的性质、成分和作用
小肠液是一种弱碱性液体,pH约为7.6,渗透压与血浆相等。小肠液的分泌量变化范围很大,成年人每日分泌量约1-3L。大量的小肠液可以稀释消化产物,使其渗透压下降,有利于吸收。小肠分泌后又很快地被绒毛重吸收,这种液体的交流为小肠内营养物质的吸收提供了媒介。
在各种不同条件下,小肠液的性状变化也很大,有时是较稀的液体,而有时则由于含有大量粘蛋白而很粘稠。小肠注保还常混有脱落的肠上皮细胞、白细胞,以及由肠上皮细胞分泌的免疫球蛋白。
近年来认为,真正由小肠腺分泌的酶只有肠致活酶一种,它能激活胰液中的胰蛋白酶原,使之变有活性的胰蛋白酶,从而有利于蛋白质的消化。小肠本身对食物的消化是以一种特殊的方式进行的,即在小肠上皮细胞的纹状缘和上皮细胞内进行的。在肠上皮细胞内含有多种消化酶,如分解多肽的肽酶、分解双糖的蔗糖酶和麦芽糖酶等。这些存在于肠上皮细胞内的酶可随脱落的肠上皮细胞进入肠腔内,但它们对小肠内消化并不起作用。
(二)小肠液分泌的调节
小肠液的分泌是经常性的,但在不同条件下,分泌量的变化可以很大。食糜对粘膜的局部机械刺激和化学刺激都可引起小肠液的分泌。小肠粘膜对扩张刺激最为敏感,小肠内食糜的量越多,分泌也越多。一般认为,这些刺激是通过肠壁内神经丛的局部反射而引起肠腺分泌的。刺激迷走神经可引起十二指肠蝗分泌,但对其它部位的肠腺作用并不明显,有人认为,只有切断内脏大神经(取消了抑制性影响)后,刺激迷走神经才能引起小肠液的分泌。
在胃肠激素中,胃泌素、促胰液素、胆囊收缩素和血管活性肠肽都有刺激小肠分泌的作用。
四、小肠的运动
小肠的运动功能是靠肠壁的两层平滑肌完成的。肠壁的外层是纵行肌,内层是环行肌。
(一)小肠的运动形式
小肠的运动形式包括紧张性收缩、分节运动和蠕动三种。
1.紧张性收缩 小肠平滑肌紧张性是其它运动形式有效进行的基础。当小肠紧张性降低时,肠腔易于扩张,肠内容物的混合和转运减慢;相反,当小肠紧张性升高时,食糜在小肠内的混合和运转过程就加快。
2.分节运动 这是一种以环行肌为主的节律性收缩和舒张运动。在食糜所在的一段肠管上,环行肌在许多点同时收缩,把食糜分割成许多节段;随后,原来收缩处舒张,而原来舒张处收缩,使原来的节段分为两半,而相邻的两半则合拢来形成一个新的节段;如此反复进行,食糜得以不断地分开,又不断地混合(图6-22)。分节运动的推进作用很小,它的作用在于使食糜与消化液充分混合,便于进行化学性消化,它还使食糜与肠壁紧密接触,为吸收创造了良好的条件。分节运动还能挤压肠壁,有助于血液和淋巴的回流。
分节运动在空腹时几乎不存在,进食后才逐渐变强起来。小肠各段分节运动的频率不同,小肠上部频率较高,下部较低。在人,十二指肠分节运动的频率约为每分钟11次,回肠末端为每分钟8次。这种活动梯度对于食糜从小肠的上部向下部推进具有一定意义。
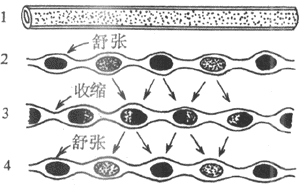
图6-22 小肠的分节运动模式图
1.肠管表面观 2、3、4:肠管切面观,示不同阶段的食糜节段分割和合拢情况
电生理研究指出,小肠分节运动的梯度现象与其平滑肌的基本电节律有关。小肠平滑肌的基本电节律的起步点位于十二指肠近胆管入口处的纵行细胞上,其频率在人约为每分钟11次。从十二指肠到回肠末端,基本电节律的频率逐渐下降,但在完整的小肠内,上部具有较高频率的肠段可控制其下部频率较低的一段肠段。因此。实际上在小肠全长中,其内在节律形成了数个频率平台。
3.蠕动 小肠的蠕动可发生在小肠的任何部位,其速率约为0.5-2.0cm/s,近端小肠的蠕动速度大于远端。小肠蠕动波很弱,通常只进行一段短距离(约数厘米)后即消失。蠕动的意义在于使经过分节运动作用的食糜向前推进一步,到达一个新肠段,再开始分节运动。食糜在小肠内实际的推进速度只有1cm/min,也就是说,食糜从幽癯部到回盲瓣,大约需要历时3—5小时。
在小肠还常可见到一种进行速度很快(2-25cm/s)、传播较远的蠕动,称为蠕动冲。蠕动冲可把食糜从小肠始端一直推送到大肠。蠕动冲可能是由于进食时吞咽动作或食糜进入十二指肠而引起的。
消化间期小肠的波动 动物或人在消化间期或禁食期,小肠的运动形式与消化期不同,呈周期性变化,称为移行性运动综合波(migrating motlity complex,MMC)。MMC以一定的间隔在胃或小肠上部发生,沿着肠管向肛门方向移行。在传播途中,其移行速度逐渐减慢。当一个波群到达回盲肠时,另一波群又在十二指肠发生,其间隔通常为90-120min 。
综合波的每一周期一般包括四个时相:I相(静止时相),此时只能记录到慢波电位,不出现胃肠收缩,持续约30-60min;Ⅱ相出现不规律的锋电位,其频率和振幅逐渐增加,持续15-40min;Ⅲ相时每个慢波电位上都叠加有成簇的锋电位,并引起相应部位发生强烈的收缩,持续4-8min;Ⅳ相与下一个周期之间为一个持续约5min的过渡葙,即N相,此进锋电位突然消失(图6-23)。
MMC的生理意义尚不完全清楚。一般认为,在Ⅱ相和Ⅲ相(特别Ⅲ相)出现的强力收缩掠过小肠时,可将肠内容物,包括上次进餐后遗留的残渣、脱落的细胞碎片和细菌等清除干净,因而有消化间期“管家人”之称。此外,通过这种周期性运动,可使小肠的肌肉在长期禁食期内保持良好的功能状态。消化间期肠运动不良的患者常伴有肠内细菌的过度繁殖。
图6-23狗小肠消化间期的移行性运动综合波及其与血浆胃动素浓度的密切关系
MMC的发生和移行受神经和激素的调节。迷走神经兴奋使周期缩短;禁食期间由肠粘膜中释放的胃动素(motlin),其血浆中浓度的峰值与MMC的Ⅲ相开始相符合,且外源性注射胃动素可诱发禁食动物出现额外的周期。因此,胃动素被认为是诱发MMC的激素。
(二)小肠运动的调节
1.内在神经丛的作用位于纵行肌和环行肌之间的肌间神经丛对小肠运动起主要调节作用。当机械和化学刺激作用于肠壁感受器时,通过局部反射可引起平滑肌的蠕动运动。切断小肠的外来神经,小肠的蠕动仍可进行。
2.外来神经的作用一般业说,副交感神经的兴奋能加强肠运动,而交感神经兴奋则产生抑制作用。但上述效果还依肠肌当时的状态而定。如肠肌的紧张性高,则无论副交感或交感神经兴奋,都使之抑制;相反,如肠肌的紧张性低,则这两种神经兴奋都有增强其活动的作用。
3.体液因素的作用小肠壁内的神经丛和平滑肌对各种化学物质具有广泛的敏感性。除两种重要的神经递乙酰胆碱和去甲肾上腺素外,还有一些肽类激素和胺,如P物质、脑啡肽和5-羧色胺,都有兴奋肠运动的作用。
(三)回盲括约肌的功能
回肠末端与盲肠交界外的环行肌显着加厚,起着括约肌的作用,称为回盲括约肌。回盲括约肌在平时保持轻度收缩状态,其内压力约比结肠内压力高2.67kPa(20mmHg)。
对盲肠粘膜的机械刺激或充胀刺激,可通过肠肌局部反射,引起括约肌收缩,从而阻止回肠内容物向盲肠排放。进食时,当食物进入胃时,可通过胃-回肠反射引起回肠蠕动,在蠕动波到达回肠末端最后数厘米时,括约肌便舒张,这样,当蠕动波到达时,大约有4ml食糜由回肠被驱入结肠。此外,胃幽门部中释放的胃泌素也能引起括约肌内的压力下降。
总之,回盲括约肌的主要功能是防止回肠内容物过快地进入大肠,延长食糜在小肠内停留的时间,因此有利于小肠内容物的完全消化和吸收。据统计,正常情况下每天约有450-500ml食糜进入大肠。此外,回盲括约肌不定还具有活瓣样作用,它可阻大肠内容物向回肠倒流。
小肠内容物向大肠的排放,除与回盲括约肌的活动有关外,还与食糜的流动性和回肠与结肠内的压力差有关:食糜越稀,通过回盲瓣也越容易;小肠腔内压力升高,也可迫使食糜通过括约肌。
第五节 大肠内消化
人类的大肠内没有重要的消化活动。大肠的主要功能在于吸收水分,大肠还为消化后的残余物质提供暂时贮存所。
一、大肠液的分泌
大肠液是由在肠粘膜表面的柱状上皮细胞及杯状细胞分泌的。大肠的分泌富含粘液和硕酸氢盐,其pH为8.3-8.4。大肠液中可能含有少量二肽酶和淀粉酶,便它们对物质的分解作用不大。大肠液的主要作用在于其中的粘液蛋白,它能保护扬粘膜和润滑粪便。
大肠液的分泌主要是由食物残渣对肠壁的机械性刺激引起的。刺激副交感神经可使分泌增加,而刺激交感神经则可使正在进行着的分泌减少。尚未发现重要的体液调节。
二、大肠的运动和排便
大肠的运动少而慢,对刺激的反应也较迟缓,这些特点对于大肠作为粪便的暂时贮存所是适合的。
(一)大肠运动的形式
1.袋状往返运动这是在空腹时最多见的一种运动形式,由环行肌无规律地收缩所引起,它使结肠袋中的内容物向两个方向作短距离的位移,但并不向前推进。
2.分节或多袋推进运动这是一个结肠袋或一段结肠收缩,其内容物被推移到下一段的运动。进食后或结肠受到拟副并感药物刺激时,这种运动增多。
3.蠕动大肠的蠕动是由一些稳定向前的收缩波所组成。收缩波前方的肌肉舒张,往往充有气体;收缩波的后面则保持在收缩状态,使这段肠管闭合并排空。
在大肠还一种进行很快,且前进很远的蠕动,称为集团蠕动。它通常开始于横结肠,可将一部分大肠物推送至降结肠或乙状结肠。集团蠕动常见于进食后,最常发生在早餐后60min之内,可能是胃内食物进入十二指肠,由十二指肠-结肠反射所引起。这一反射主要是通过内在神经丛的传递实现的。
(二)排便
食物残渣在大肠内停留的时间较长,一般在十余小时以上,在这一过程中,食物残渣中的一部分水分被大肠粘膜吸收。同时,经过大肠同细菌的发酵和腐败作用,形成了粪便。粪便中除食物残渣外,还包括脱落的肠上皮细胞和大量的细菌。此外,机体代谢后的废物,包括由肝排出的胆色素衍生物,以及由血液通过肠壁排至肠腔中的某些金属,如钙、镁、汞等的盐类,也随粪便排至体外。
正常的直肠通常是空的,没有粪便在内。当肠的蠕动将粪便推入直肠时,刺激了直肠壁内的感受器,冲动经盆神经和腹下神经传至脊髓腰骶段的初级排便中枢,同时上传到大脑皮层,引起便意和排便反射。这时,通过盆神经的传出冲动,使降结肠、乙状结肠直肠收缩,肛门内括约肌舒。与此同时,阴部神经的冲动减少,肛门外括约肌舒张,使粪便排排出体外。此外,由于支配腹肌和膈肌的神经兴奋,腹肌和膈肌也发生收缩,腹内压增加,促进粪便的排出。正常人的直肠对粪便的压力刺激具有一定的阈值,当达到此阈值时即可引起便意。
排便运动受大脑皮层的影响是显而易见的,意识可以加强或抑制排便。人们对对便意经常予以制止,就使直肠渐渐地对粪便压力刺激失去正常的敏感性,加之粪便在大肠内认定留过久,水分吸收过多而变得干硬,引起排便困难,这是产生便秘的最常见的原因之一。
(三)大肠内细菌的活动
大肠内有许多细菌。细菌主要来自食物和空气,它们由口腔入胃,最后到达大肠。大肠内的酸碱度和温度对一般细菌的繁殖极为适宜,细菌便在这里大量繁殖。细菌中含有能分解食物残渣的酶。糖及脂肪的分解称为发酵,其产物有乳酸、醋酸、二氧化碳、沼气、脂肪酸、甘油、胆碱等。蛋白质的细菌分解称为腐败,其产物有胨、氨基酸、氨、硫化氢、组胺、吲哚等,其中有的成分由肠壁吸收后到肝中解毒。
大肠内的细菌能利用肠内较为简单的物质合成维生素B复合物和维生素K,它们由肠内吸收后,对人体有营养作用。
据估计,粪便中死的和活的细菌约占粪便固体重量的20-30%。
(四)食物中纤维素对肠功能的影响
近年来,对于食物中纤维素对肠功能和肠疾病发生的影响,引起了医学界极大的重视。事实证明,适当增加纤维素的摄取有增进健康,预防便秘、痔疮、结肠癌等疾病的作用。食物中纤维素对胃肠功能的影响主要有以下方面:①大部分多糖纤维能与水结合而形成凝胶,从而限制了水的吸收,并使肠内容物容积膨胀加大;②纤维素多能刺激肠运动,缩短粪便在肠内停留时间和增加粪便容积;③纤维素可降低食物中热量的比率,减少含能物质的摄取,从而有助于纠正不正常的肥胖。
第六节 吸 收
消化管内的吸收是指食物的成分或其消化后的产物,通过上皮细胞进入血液和淋巴的过程。消化过程是吸收的重要前提。由于吸收为多细胞机体提供了营养,因而具有很大的生理意义。
消化管不同部位的吸收能力有吸收速度是不同的,这主要取决于各部分消化管的组织结构,以及食物在各部位被消化的程度和停留的时间。在口腔和食管内,食物实际上是不被吸收的。在胃内,食物的吸收也很少,胃可吸收酒精和少量水分。小肠是吸收的主要部位,一般认为,糖类、蛋白质和脂肪的消化产物大部分是在十二指肠和空肠吸收的,回肠有其独特的功能,即主动吸收胆盐和维生素B12(图6-24)。对于大部分营养成分,当它到达回肠时,通常已吸收完毕,因此回肠主要是吸收功能的贮备。小肠内容物进入大肠时已经不含多少可被吸收的物质了。大肠主要吸收水分和盐类,一般认为,结肠可吸收进入其内的80%的水和90%的Na+和CI-。
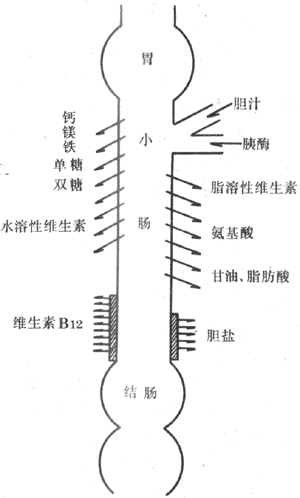
图6-24 各种主要营养物质在小肠的吸收部位
人的小肠长约4m,它的粘膜具有环形皱褶,并拥有大量的绒毛,绒毛是小肠粘膜的微小突出构造,其长度约0.5-1.5mm。每一条绒毛的外面是一层柱状上皮细胞。在显微镜下观察,可见柱状上皮细胞顶端有明显有纵纹,电了显微镜下的观察进一步表明,纵纹乃是柱状细胞顶端细胞膜的突出,被称为微绒毛。人的肠绒毛上,每一柱状上皮细胞的顶端约有1700条微绒毛。由于环状皱褶、绒毛和微绒毛的存在,最终使小肠的吸收面积比同样长短的简单圆筒的面积增加约600倍,达到200m2左右(图6-25)。小肠除了具有巨大的吸收面积外,食物在小肠内停留的时间较长(3-8h),以及食物在小肠内已被消化到适于吸收的小分子物质,这些都是小肠在吸收中发挥作用的有利条件。
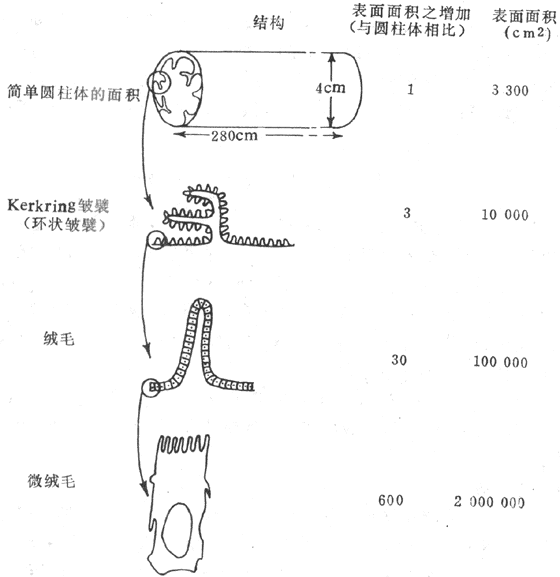
图 6-25 增加小肠表面面积的三种机制
人小肠的长度据Hirsch等报道,半径据Gray报道。由于Kerkring皱壁所致的表面面积的增
大是作用的估计。Verzar估计,在大鼠和鸽子,由于绒毛所致的面积之增大为8。
由于微绒毛所致的面积之增大的小鼠为14(Zetterqvixt的估计),
在大鼠为24(Palay 和Karlin的估计)。
小肠绒毛内部有毛细血管、毛细淋巴管、平滑肌纤维和神经纤维网等结构。动物在空腹时,绒毛不活动。进食则可引起绒毛产生节律性的伸缩和摆动。这些运动可加速绒毛内血液和淋巴的流动,有助于吸收。绒毛运动神经控制,刺激内脏神经可加强绒毛运动。绒毛运动还受小肠粘膜中释放的一种胃肠激素——绒毛收缩素(villkinin)的刺激。
营养物质和水可以两条途径进入血液或淋巴:一为跨细胞途径,即通过绒毛柱状上皮细胞的腔面膜进入细胞,再通过细胞底-侧面膜进入血液或淋巴;另一为旁细胞途径,即物质或水通过细胞间的紧密连接,进入细胞间隙,然后再转入血液或淋巴(图6-26)。营养物质通过膜的机制包括扩散、易化扩散、主动转运及胞饮等,其过程可参看第二章第一节。
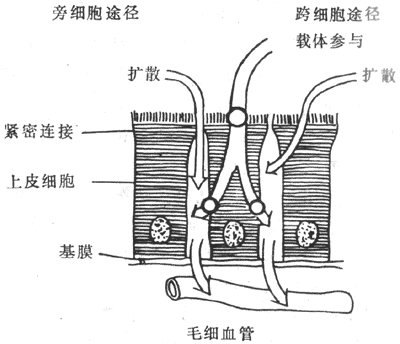
图6-26 小肠粘膜吸收水和小的溶质的两条途径
二、小肠内主要营养物质的吸收
在小肠中被吸收的物质不仅是由口腔摄入的物质,由各种消化腺分泌入消化管内的水分、无机盐和某些有机成分,大部分将在小肠中被重吸收。例如,人每日分泌入消化管内的各种消化液总量可达6-7L之多,每日还从口腔摄入1L多的水分,而每日由粪便中丢失的水分只有150ml左右。因此,重吸收回体内的液体量每日可过8L。这样大量的水分如果不被重吸收,势必严重影响内环境的相对稳定而危及生命,急性呕吐和腹泻时,在短时间内损失大量液体的严重性就在于此。
在正常情况下,小肠每天还吸收几百克糖,100g或更多的脂肪,50-100g氨基酸,50-100g离子等。实际上,小肠吸收的能力远远超过这个数字,因此,小肠的吸收具有巨大的贮备力。
(一)水分的吸收
前已述,人每日由胃肠吸收回体内的液体量约有8L之多。水分的吸收都是被动的,各种溶持,特别是NaCI的主动吸收所产生的渗透压梯度是水分吸收的主要动力。细胞膜和细胞间的紧密连接对水的通透性都很大,因此,驱使水吸收的渗透压一般只有3-5mOs/L。
在十二指肠和空肠上部,水分由肠腔进入血液的量和水分由血液进入肠腔的量都很大,因此肠腔内液体的量减少得并不多。在回肠,离开肠腔的液体比进入的多,从而使肠内容大为减少。
(二)无机盐的吸收
一般说,单价碱性盐类如钠、钾、铵盐的吸收很快,多价碱性盐类则吸收很慢。凡能与钙结合而形成沉淀的盐,如硫酸盐、磷酸盐、草酸盐等,则不能被吸收。
1.钠的吸收 成人每日摄入约250-300mmol的钠,消化腺大致分泌相同数量的钠,但从粪便中排出的钠不到4mmol,说明肠内容中95%-99%的钠都被吸收了。
由于细胞内的电位较粘膜面负40V,同时细胞内钠的浓度较周围液体为低,因此,钠可顺电化学梯度通过扩散作用进入细胞内。但细胞内的钠能通过低-侧膜进入血液,这是通过膜上钠泵的活动逆电化学进行的主动过程(图6-27)。钠泵是一种Na+-K+依赖性ATP酶,它可使ATP分解产生能量,以维持钠和钾逆浓度的转运。钠的泵出和钾的泵入是耦联的
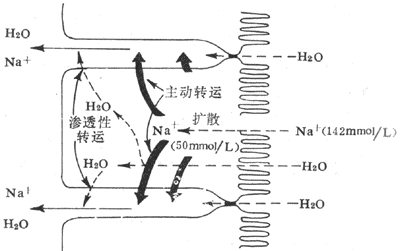
图6-27 小肠粘膜对钠和水的吸收
2.铁的吸收 人每日吸收的铁约为1mg,仅为每日膳食中含铁量的1/10。铁的吸收与机体对铁的需要有关,当服用相同剂量的铁后,缺铁的患者可比正常人的铁吸收量大1-4倍。食物中的铁绝大部分是三价的高铁形式,但有机铁和高铁都不易被吸收,故须还原为亚铁后,方被吸收。亚铁吸收的速度比相同量的高铁要快2-5倍。维生素C能将高铁还原为亚铁而促进铁的吸收。铁在酸性环境中易溶解而便于被吸收,故胃液中的盐酸有促进铁吸收的作用,胃大部切除的病人,常常会伴以缺铁性贫血。
铁主要在小肠上部被吸收。肠粘膜吸收铁的能力决定于粘膜细胞内的含铁量。由肠腔吸收入粘膜细胞内的无机铁,大部分被氧化为三价铁,并和细胞内丰硕睥去铁铁蛋白结合,形成铁蛋白,暂时贮存在细胞内,慢慢地向血液中释放。一小分部被吸收入粘膜细胞而尚未与去铁铁蛋白结合的亚铁,则可以主动吸收的方式转移到血浆中。当粘膜细胞刚刚吸收铁而尚未能转移至血浆中时,则暂时失去其由肠腔再吸收铁的能力。这样,存积在粘膜细胞内的铁量,就成为再吸收铁的抑制因素。
3.钙的吸收 食物中的钙仅有一小部分被吸收,大部分随粪便排出。主要影响钙吸收的因素是维生素D和机体对钙的需要。维生素DC有促进小肠对钙吸收的作用。儿童和乳母对钙的吸收增加。此外,钙盐只有在水溶液状态(如氯化钙、葡萄糖酸钙溶液),而且在不被肠腔中任何其他物质沉淀的情况下,才能被吸收。肠内容的酸度对钙的吸收有重要影响,在pH约为3时,钙呈离子化状态,吸收最好。肠内容中磷酸过多,会形成不溶解的磷酸钙,使钙不能被吸收。此外,脂肪食物对钙的吸收有促进作用,脂肪分解释放的脂肪酸,可与钙结合形成钙皂,后者可和胆汁酸结合,形成水溶性复合物而被吸收。
钙的吸收主要是通过主动转动完成的。肠粘膜细胞的微绒毛上有一种与钙有高度亲和性的钙结合蛋白,它参与钙的转运而促进钙的吸收。
4.负离子的吸收 在小肠内吸收的负离子主要是CI-t HCO3。由钠泵产生的电位差可促进肠腔负离子向细胞内移动。但也有证据认为,负离子也可以独立地移动。
(三)糖的吸收
糖类只有分解为单糖时才能被小肠上皮细胞所吸收。各种单糖的吸收速率有很大差别,已糖的吸收很快,而戊糖则很慢。在已糖中,又以半乳糖和葡萄糖的吸收为最快,果糖次之,甘露糖最慢。
单糖的吸收是消耗能量的主动过程,它可逆着浓度差进行,能量来自钠泵,属于继发性主动转运(参见第二章)。在肠粘膜上皮细胞的纹状缘上存在着一种转运体蛋白,它能选择性地把葡萄糖和半乳糖从纹状的肠腔面运入细胞内,然后再扩散入血。各种单糖与转运体蛋白的亲和力不同,从而导致吸收的速率也不同。
转运体蛋白在转运单糖的同时,需要钠的存在。一般认为,一个转运体蛋白可与两个Na+和一个葡萄糖分子结合。由此可见,钠对单糖的主动转运是必需的。用抑制钠泵的哇巴因,或用能与Na+竞争转运体蛋白的K+,均能抑制糖的主动转运。
(四)蛋白质的吸收
无论是食入的蛋白质(100g/d)或内源性蛋白质(25-35g/d),经消化分解为氨基酸后,几乎全部被小肠吸收。经煮过的蛋白质因变性而易于消化,在十二指肠和近端空肠就被迅速吸收,未经煮过的蛋白质和内源性蛋白质较难消化,需进入回肠后才基本被吸收。
氨基酸的吸收是主动性的。目前在小肠壁上已确定出3种主要的转运氨基酸的特殊运载系统,它们分别转动中性、酸性或碱性氨基酸。一般来讲,中性氨基酸的转运比酸性或碱性氨基酸速度快。与单糖的吸收相似,氨基酸的吸收也是通过与钠吸收耦联的,钠泵的活动被阻断后,氨基酸的转运便不能进行。氨基酸吸收的路径几乎完全是经血液的,当小肠吸收蛋白质后,门静脉血液中的氨基酸含量即行增加。
曾经认为,蛋白质只有水解成氨在酸后才能被吸收。但近年来的实验指出,小肠的纹状缘上还存在有二肽和三肽的转运系统,因此,许多二肽和三肽也可完整地被小肠上皮细胞吸收,而且肽的转运系统吸收效率可能比氨基酸更高。进入细胞内的二肽和三肽,可被细胞内的二肽酶和三肽酶进一步分解为氨基酸,再进入血液循环。
完整的蛋白质是还可被人的小肠上皮细胞吸收?许多实验证明,小量的食物蛋白可完整地进入血液,由于吸收的量很少,从营养的角度来看是无意义的;相反,它们常可作为抗原而引起过敏反应或中毒反应,对人体不利。
(五)脂肪的吸收
在小肠内,脂类的消化产物脂肪酸、甘油一酯、胆固醇等很快与胆汁中的胆盐形成混合微胶粒。由于胆盐有亲水性,它能携带脂肪消化产物通过覆盖在小肠绒毛表面的非流动水层到达微绒毛上。在这里,甘油一酯、脂肪酸和胆固醇等又逐渐地从混合胶粒中释出,它们透过微绒毛的脂蛋白膜而进入粘膜细胞(胆盐被遗留于肠腔内)。
长链脂肪酸及甘油酯被吸收后,在肠上皮细胞的内质网中大部分重新合成为甘油三酯,并与细胞中生成的载脂蛋白合成乳糜微粒(chylomicron)。乳糜微粒一旦形成即进入高尔基复合体中,乳糜微粒被包裹在一个囊泡内。囊泡移行到细胞底-侧膜时,便与细胞膜融合,释出乳糜微粒进入细胞间隙,再扩散入淋巴。(图6-28)
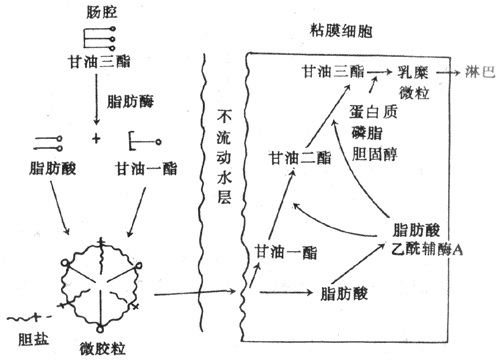
图6-28 脂肪在小肠内消化和吸收的主要方式
中、短链甘油三酯水解产生的脂肪酸和甘油一酯,在小肠上皮细胞中不再变化,它们是水溶性的,可以直接进入门脉而不入淋巴。由于膳食的动、植物油中含有15个以上碳原子的长链脂肪酸很多,所以脂肪的吸收途径乃以淋巴为主。
(六)胆固醇的吸收
进入肠道的胆固醇主要有两下来源:一是食物中来的,一是肝分泌的胆汁中来的。由胆汁来的胆固醇是游离的,而食物中的胆固醇部分是酯化的。酯化的胆固醇必须在肠腔中经消化液中的胆固醇酯酶的作用,水解为游离胆固醇后才能被吸收。游离的胆固醇通过形成混合微胶粒,在小肠上部被吸收。被吸收的胆固醇大部分在小肠粘膜中又重新酯化,生成胆固醇酯,最后与载脂蛋白一起组成乳糜微粒经由淋巴系统进入血循环。
胆固醇的吸收受很多因素的影响。 食物中胆固醇含量越高,其吸收也越多,但两者不呈直线关系。食物中的脂肪和脂肪酸有提高胆固醇吸收的作用,而各种植物固醇(如豆固醇、β-谷固醇)则抑制其吸收。胆盐可与胆固醇形成混合微胶粒而助于胆固醇的吸收,食物中不能被利用的纤维素、果胶、琼脂等容易和胆盐结合形成复合物,妨碍微胶粒的形成,从而能降低胆固醇的吸收;最后,抑制肠粘膜由细胞载脂蛋白合成的物质,可因妨碍乳糜微粒的形成,减少胆固醇的吸收。
参考资料
1.徐丰彦,张镜如主编.人体生理学.北京:人民卫生出版社,1989
2.王志均.梅懋华,朱文玉.胃肠激素.北京:科学出版社,1985
3.梅懋华主编,消化道生理学与临床.北京:人民卫生出版社,1990
4.周吕主编.胃肠生理学:基础与临床.北京:科学出版社,1991
5.Johnson LR.GastrointestinalPhysiology 4thed,CV Mosby Co,St Louis,1991
6.Davenport HW.Physiology ofthedigestive tract 5thed,Year Book MedicalPublishers,inc,chicago,1982
7.wood jd ed,handbook ofphysiology,section 6:the gastrointestinal system,american physiologysociety,bethesda,1989
8.sell ah,wash jh.regulation ofgastric acid secretion.ann rev physiol 1979;41:35-53
9.sarna s.cause and effectrdlationship betweem motilin and migrating myoelectric complex,am j physiol1983;245:277-294
10.hootmed sr.willamsja.stmulus-secretion coupling in the pancreatic acinus. In physioloy of thegastrointestinal tract,lr johmson ed 2nded,raven press,new york,1987
第七章 能量代谢和体温
第一节 能量代谢
新陈代谢是机体生命活动的基本特征,新陈代谢包括物质代谢与相传伴的能量代谢,简称代谢。
糖、脂肪、蛋白质三种营养物质,经消化转变成为可吸收的小分子营养物质而被吸收入血。在细胞中,这些营养物质经过同化作用(合成代谢),构筑机体的组成成分或更新衰老的组织;同时经过异化作用(分解代谢)分解为代谢产物。合成代谢和分解代谢是物质代谢过程中互相联系的、不可分割的两个侧面。
在分解代谢过程中,营养物质蕴藏的化学能便释放出来。这些化学能经过转化,便成了机体各种生命活动的能源,所以说分解是代谢的放能反应。而在合成代谢过程中,需要供给能量,因此是吸能反应。可见,在物质代谢过程中,物质的变化与能量的代谢是紧密联系着的。生物体内物质代谢过程中所伴随的能量释放、转移和利用等,称为能量代谢(energy metabolism)。
机体所需的能量来源于食物中的糖、脂肪和蛋白质。这些能源物质分子结构中的碳氢键蕴藏着化学能,在氧化过程中碳氢键断裂,生成CO2和 H2O,同时释放出蕴藏的能。这些能量的50%以上迅速转化为热能,用于维持体温,并向体外散发。其余不足50%则以高能磷酸键的形式贮存于体内,供机体利用。体内最主要的高能磷酸键化学物是三磷酸腺苷(ATP)。此外,还可有高能硫酯键等。机体利用ATP去合成各种细胞组成分子、各种生物活性物质和其他一些物质;细胞利用ATP去进行各种离子和其它一些物质的主动转运,维持细胞两侧离子浓度差所形成的势能;肌肉还可利用ATP所载荷的自由能进行收缩和舒张,完成多种机械功。总的看来,除骨骼肌运动时所完成的机械功(外功)以外,其余的能量最后都转变为热能。例如心肌收缩所产生的势能(动脉血压)与动能(血液流速),均于血液在血管内流动过程中,因克服血流内、外所产生的阻力而转化为热能。在人体内,热能是最“低级”形式的能,热能不能转化为其它形式的能,不能用来作功。
本节主要叙述整个机体的能量代谢测定的原理与方法,基础代谢以及机体在某些状态下的代谢等问题,不涉及能量代谢的各个方面。
通用的法定计量的热量单位为焦耳(Joules.J)。过去热量单位是卡或千卡,1卡=4.187焦耳,1千卡=4.187千焦耳(kJ)。1焦耳/s为1瓦特。
一、能量代谢测定的原理和方法
热力学第一定律指出:能量由一种形式转化为另一种形式的过程中,既不能增加,也不减少。这是所有形式的能量(动能、热能、电能入化学能)互相转化的一般规律,也就是能量守恒定律。机体的能量代谢也遵循这一规律,即在整个能量转化过程中,机体所利用的蕴藏于食物中的化学能与最终转化成的热能和所作的外功,按能量来折算是完全相等的。因此,测定在一定时间内机体所消耗的食物,或者测定机体所产生的热量与所做的外功,都可测算出整个机体的能量代谢率(单位时间内所消耗的能量)。
测定整个机体单位时间内发散的总热量,通常有两类方法:直接测热法和间接测热法。
(一)直接测热法
直接测热法(direct calormetry)是测定整个机体在单位时间内向外界环境发散的总热量。此总热量就是能量代谢率。如果在测定时间内做一定的外功,应将外功(机械功)折算为热量一并计入。图7-1是本世纪初Arwater-Benedict所设计的呼吸热量计的结构模式图。它是由隔热密封的房间,其中设一个铜制的受试者居室。用调节温度的装置控制隔热壁与居室之间空气的温度,使之与居室内的温度相等,以防居室内的热量因传导而丧失。这样,受试者机体所散发的大部分热量便被居室内管道中流动的水所吸收。根据流过管道的水量和温度差,将水的比热考虑在内,就可测出水所吸收的热量。当然,受试者发散的热量有一部分包含在不感蒸发(参看第二节)量中,这在计算时也要加进去。受试者呼吸的空气由进出居室的气泵管道系统来供给。此系统中装有硫酸和钠石灰,用业吸收水蒸气和CO2。管道系统中空气中的O2则由氧气筒定时补给。
直接测热法的设备复杂,操作繁锁,使用不便,因而极少应用。一般都采用间接测热法。
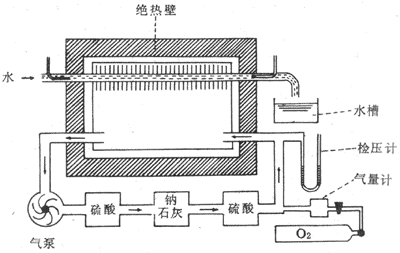
图7-1 直接测热装置示意图
(二)间接测热法
在一般化学反应中,反应物的量与产物量之间呈一定的比例关系,这就是定比定律。例如,氧化1mol葡萄糖,需要6mol氧,同时产生6molCO2和6molH2O,并释放一定量的能。下列反应式表明了这种关系:
C6H12O6+602→6CO2+6H20+△H
同一种化学反应,不论经过什么样的中间步骤,也不论反应条件差异多大,这种定比关系仍然不变。例如,在人本内氧化1mol葡萄糖,同在体外氧化燃烧1mol葡萄糖一样,都要消耗6molCO2和6molH20,而且产生的热量也相等。一般化学反应的这种基本规律也见于人体内营养物质氧化供能的反应(蛋白质的情况下有些出入,参看下文),所以它成了能量代谢间接测热法的重要依据。
间接测热法(indirectcalorimetry)的基本原理就是利用这种定比关系,查出一定时间内整个人体中氧化分解的糖、脂肪、蛋白质各有多少,然后据此计算出该段时间内整个机体所释放出来的热量。因此,必须解决两个问题:一是每种营养物质氧化分解时产生的能量有多少(即食物的热价);二要分清三种营养物质各氧化了多少。
食物的热价应用弹式热量计,在体外测定了一定量的的糖、脂肪和蛋白质燃烧时所释放的热量,并同这三类物质在动物体内氧化到最终产物C02和水时所产生的热量相比较,证明了糖和脂肪在体外燃烧与在体内氧化分解所产生的热量是相等的。于是将1g食物氧化(或在体外燃烧)时所释放出来的能量称为食物的热价(thermalequivalent of food)。食物的热价分为物理热价和生物热价。前者指食物在体外燃烧时释放的热量,后者系食物经过生物氧化所产生的热量。糖(或脂肪)的物理热价和生物热价是相等的,而蛋白质的生物热价则小于它的物理热价。因为蛋白质在体内不能被彻底氧化分解,它有一部分主要以尿素的形式从尿中排泄的缘故。三种营养物质在物理热价和生物热价见表演7-1。
呼吸商 机体依靠呼吸功能从外界摄取氧,以供各种营养物质氧化分解的需要,同时也将代谢终生物CO2呼出体外,一定时间内机体的CO2产量与耗氧量的比值称为呼吸商(respiratory quotient, RQ)。各种营养物质在细胞内氧化供能属于细胞呼吸过程,因而双将各种营养物质氧化时的CO2产量与耗氧量的比值称为某物质的呼吸商。严格说来,应该以CO2和O2的克分子(mol)比值来表示呼吸商。但是,因为在同一温度和气压条件下,容积相等的不同气体,其分子数都是相等的,所以通常都用容积数(ml或L)来计算CO2与O2的比值,即:
![]()
糖、脂肪和蛋白质氧化时,它们的CO2产量与耗氧量各不相同,三者的呼吸商也不一样。
因为各种营养物质无论在体内或体外氧化,它们的耗氧量与CO2产量都取决于各该物质的化学组成,所以,在理论上任何一种营养物质的呼吸商都可以根据它的氧化成终产物(CO2和H20)化学反应式计算出来的。
糖的一般分子式为(CH20)n,氧化时消耗的02和产生的C02分子数相等,呼吸商应该等于1。如上述葡萄糖氧化的反应式所示,C02产量与耗氧量均为6mol故:
![]()
脂肪氧化时需要消耗更多的氧。在脂肪本身的分子结构中,氧的含量远较碳和氢少。因此,另外提供的氧不仅要用氧化脂肪分子中的碳,还要用来氧化其中的氢。所以脂肪的呼吸商将小于1。现以甘油三酸酯(triolein)为例:
![]()
RQ=57molC02/80mol02=0.71
蛋白质的呼吸商较难测算,因为蛋白质在体内不能完全氧化,而且它氧化分解途径的细节,有些还不够清楚,所以只能通过蛋白质分子中的碳和氢被氧化时势需氧量和C02产量,间接算出蛋白质的呼吸商,其计算值为0.80。
在人的日常生活中,营养物质不是单纯的,而是糖、脂肪和蛋白质混合而成的(混合膳食)。所以,呼吸商常变动于0.71-1.00之间。人体在特定时间内的呼吸产要看哪种营养物质是当时的主要能量来源而定。若能源主要是糖类,则呼吸商接近于1.00;若主要是脂肪,则呼吸商接近于0.71。在长期病理性饥饿情况下,能源主要来自机体本身的蛋白质和脂肪,则呼吸商接近于0.80。一般情况下,摄取混合食物时,呼吸商常在0.85左右。
现将糖、脂肪和蛋白质三者的热价、氧热窬及呼吸商等数据列于表7-1,以供理解和测算能量代谢率之用。
表7-1 三种营养物质氧化时的几种数据
| 营养物质 | 产热量(kJ/g) | 耗氧量 | CO2产量 | 氧热价 | 呼吸商 | ||
| 物理热价(用弹式热量计测得) | 生物热价(体内生物氧化什) | 营养学热价* | |||||
| (L/g) | (L/g) | (kJ/J) | (RQ) | ||||
| 糖 | 17 | 17 | 16.7 | 0.83 | 0.83 | 21 | 1.00 |
| 蛋白质 | 23.5 | 18 | 16.7 | 0.95 | 0.76 | 18.8 | 0.80 |
| 脂肪 | 39.8 | 39.8 | 37.7 | 2.03 | 1.43 | 19.7 | 0.71 |
*营养学通常采用概数来计算食物的热价
影响呼吸商的其它因素:
机体的组织、细胞 不仅能同时氧化分解各种营养物质,而且也使一种营养物质转变为另一种营养物质。糖的转化为脂肪时,呼吸商可能变大,甚至超过1.00。这是由于当一部分糖转化为脂肪时,原来糖分子中的氧即有剩余,这些氧可能参加机体代谢过程中氧化反应,相应地减少了从外界摄取的氧量,因而呼吸商变大。反过来,如果脂肪转化为糖,呼吸商也可能低于0.71。这是由于脂肪分子中含氧比例小,当转化为糖时,需要更多的氧进入分子结构,因而机体摄取并消耗外界氧的量增多,结果呼吸商变小。另外,还有其它一些代谢反应也能影响呼吸商。例如,肌肉剧烈运动时,由于氧供不应求,糖酵解增多,将有大量乳酸进入血液。乳酸和碳酸盐作用的结果,会有大量由肺肺排出,此时呼吸商将变大。又如,肺过度通气、酸中毒等情况下,机体中与生物氧化无关的CO2大量排出,也可现呼吸大于1.00的情况。相反,肺通气不足、碱中毒等情况下,呼吸商将降低。
前已述,应该测出在一定时间内机体中糖、脂肪和蛋白质三者氧化分解的比例。为此。首先必须查清氧化了多少蛋白质,并且将氧化这些蛋白质所消耗的氧量和所产生的CO2从机体在该时间内的总耗氧量和总CO2产量中减去,算出糖和脂肪氧化(非蛋白质代谢)的CO2产量和耗氧量的比值,即非蛋白呼吸商(non-proteinrespiratory quotient,NPRQ),然后才有可能进一步查清糖和脂肪各氧化了多少克。
尿氮测定 尿中的氮物质主要是蛋白质的分解产物。因此可以通过尿氮来估算体内被氧化的蛋白质的数量。蛋白质的平均重量组成是:C50%,O223%,N16%,S1%。蛋白质中16%的N是完全随尿排出的。所以,1g 尿氮相当于氧分解6.25g蛋白质,测得的尿氮重量(g)乘以6.25,便相当于体内氧分解的蛋白质量。
非蛋白呼吸商 它是估算非蛋白代谢中糖和脂肪氧化的相对数量的依据。研究工作者早已按从0.707到1.00范围内的非蛋白呼吸产,算出糖和脂肪两者氧化的各自百分比以及氧热价(表7-2)。
表7-2 非蛋白呼吸商和氧热价
| 非蛋白呼吸商 | 氧 化 的 % | 氧热价(kJ/L) | |
| 糖 | 脂肪 | ||
| 0.707 | 0.00 | 100.0 | 19.62 |
| 0.71 | 1.10 | 98.9 | 19.64 |
| 0.72 | 4.75 | 95.2 | 19.69 |
| 0.73 | 8.40 | 91.6 | 19.74 |
| 0.74 | 12.0 | 88.0 | 19.79 |
| 0.75 | 15.6 | 84.4 | 19.84 |
| 0.76 | 19.2 | 80.8 | 19.89 |
| 0.77 | 22.8 | 77.2 | 19.95 |
| 0.78 | 26.3 | 73.7 | 19.99 |
| 0.79 | 29.0 | 70.1 | 20.05 |
| 0.80 | 33.4 | 66.6 | 20.10 |
| 0.81 | 36.9 | 63.1 | 20.15 |
| 0.82 | 40.3 | 59.7 | 20.20 |
| 0.83 | 43.8 | 56.2 | 20.26 |
| 0.84 | 47,2 | 52.8 | 20.31 |
| 0.85 | 50.7 | 49.3 | 20.36 |
| 0.86 | 54.1 | 45.9 | 20.41 |
| 0.87 | 57.5 | 42.5 | 20.46 |
| 0.88 | 60.8 | 39.2 | 20.51 |
| 0.89 | 64.2 | 35.8 | 20.56 |
| 0.90 | 67.5 | 32.5 | 20.61 |
| 0.91 | 70.8 | 29.2 | 20.67 |
| 0.92 | 74.1 | 25.9 | 20.71 |
| 0.93 | 77.4 | 22.6 | 20.77 |
| 0.94 | 80.7 | 19.3 | 20.82 |
| 0.95 | 84.0 | 16.0 | 20.87 |
| 0.96 | 87.2 | 12.8 | 20.93 |
| 0.97 | 90.4 | 9.58 | 20.98 |
| 0.98 | 93.6 | 6.37 | 21.03 |
| 0.99 | 96.8 | 3.18 | 21.08 |
| 1.00 | 100.0 | 0.0 | 21.13 |
Lusk修订
间接测热法计算原则 实验测得的机体24小时内的耗氧量和CO2产量以及尿氮量,根据表7-1和7-2中相应的一些数据计算。首先,由尿氮量算出被氧分解的蛋白质量。由被氧化的蛋白质量从表7-1中算出其产热量、耗氧量和CO2产量;其次从总耗氧量和总CO2产量中减去蛋白质耗氧量和CO2产量,计算出非蛋白呼吸商。根据非蛋白呼吸商查表7-2的相应的非蛋白呼吸商的氧热价,计算出非蛋白代谢的产热量;最后,24小时产热量为蛋白质代谢的产热量与非蛋白代谢的产热量之和。此外,从非蛋白呼吸还可推算出参加代谢的糖和脂肪的比例。
间接测热法的计算方法举例
首先测定受试者一定时间内的耗氧量和CO2产量,假定受试者24小时的耗氧量为400L,CO2产量为340L(已换算成标准状态的气体容积)。另经测定尿氮排出量为12g。根据这些数据和查表7-1、7-2,计算24小时产热量,其步骤如下:(1)蛋白质氧化量=12×6.25=7g
产热量=18×75=1350kJ
耗氧量=0.95×75=71.25L
CO2产量=0.76×75=57L
(2)非蛋白呼吸商
非蛋白代谢耗氧量=400-71.25=328.75L
非蛋白代谢CO2产量=340-57=283L
非蛋白呼吸商=283/328.75=0.86
(3)根据非蛋白呼吸商的氧热价计算非蛋白代谢的热量
查表7-2,非蛋白呼吸商为0.86时,氧热价为20.41。所以,非蛋白代谢产热量=328.75×20.41=6709.8kj 。
(4)计算24小时产热量
24小时产热量=1350+6709.8=8059.8kJ
(蛋白质代(非蛋白代谢产热量)谢产热量)
计算的最后数值8059.8kJ就是该受试者24小时内的能量代谢率
耗氧量与CO2产量的测定方法及临床应用测定耗氧量和CO2产量的方法有两种:闭合式测定法和开放式测定法。
(1)闭合式测定法:在动物实验中,将受试动物置于一个密闭的能吸热的装置中。通过气泵,不断将定量的氧气送入装置。动物不断地摄取氧,可根据装置中氧量的减少计算出该动物在单位时间内的耗氧量。动物呼出的CO2则由装在气体回路中的CO2吸收剂吸收。然后根据实验前后CO2吸收剂的重量差,算出单位时间内的CO2产量。由耗氧量和CO2产量算出呼吸商。
临床上为了简便,通常只使用肺量计(图7-2)来测量耗氧量。该装置的气体中容器中装置氧气,受试者通过呼吸口瓣将氧气吸入呼吸器官。此时气体容器的上盖随吸气而下降,并由连于上盖的描笔记录在记录纸上。根据记录纸上的方格还可读出潮气量值。受试者的呼出气则通过吸收容器(呼出气中的CO2和水可除除掉)进入气体容器中,于是气体容器的上盖又复升高,描笔也了随之升高。由于受试者摄取了一定量的氧气,呼出气中CO2又被除掉,气体容器中的氧气量因而逐渐减少。描笔则记录出曲线逐渐下降的过程。在一定时间内(通常为6min),描笔的总下降高度,就是该时间内的耗氧量。
图7-2 肺量计结构模式图
(2)开放式测定法(气体分析法):它是在机体呼吸空气的条件下测定耗氧量和CO2产量的方法,所以称为开放法。其原理是,采集受试者一定时间内的呼出气,测定呼出气量并分析呼出气中氧和CO2的容积百分比。由于吸入气就是空气,所以其中氧和CO2的容积百分比不必另测。根据吸入气和呼出气中氧和CO2的容积百分比的差数,可算出该时间内的耗氧量和CO2排出量。
气体分析方法很多,最简便而又广泛应用的方法,是将受试者在一定时间内呼出气采集于气袋中,通过气量计测定呼气量,然后用气体分析器分析呼出气的组成成分,进而计算耗氧量和CO2产量,并算出呼吸商。
现举出一个气体分析实验例。
现某健康成人安静状态下的呼出气作气体分析,结果为:O2=16.26%;CO2=4.14%。呼出气量为1分钟5.2L(通常将呼出气量换算为不含水蒸气的标准状态值;也有换算为被水蒸气饱和的1个大气压、体温状态下的值)。空气的组成是:O2=20.96%;N=79.00%;CO2=0.04%。则:
受试者从每100ml通过肺的气体中吸收的氧气为:
20.96ml-16.26ml=4.7ml(或每升气体中的47ml)
(空气) (呼出气)
因为呼出气量为5.2L,则1分钟的耗氧量为:
47ml×5.2=244.4ml
100ml呼气中的CO2为:
4.14ml-0.04ml=4.1ml(或每升气体中的41ml)
(呼出气) (空气)
1分钟CO2排出量为:
41ml×5.2=213.2ml
RQ=213.1/244=0.85
所得呼吸商为受试者在安静状态下混合膳食代谢的呼吸商。
临床上和劳动卫生常采用简略法,即用气体分析法测得一定时间内的耗氧量和CO2产量,并求出呼吸商,并且不考虑蛋白质代谢部分,就根据非蛋白呼吸商表7-2查出呼吸商的氧热价,然后将氧热价乘以耗氧量,便得出该时间内的产热量。仍以上述间接测热法计算方法列举的例子为数据(见前),按此简略法来计算,则结果如下:
呼吸商=340/400=0.85
查表7-2,呼吸商0.85时的氧热价为20.36kJ,所以24小时的产热量=20.36×400=8144kJ。
这个数值与按完整的间接法计算得出的数值8059.8kJ是很近似的,误差都在1%-2%以下。而且在非蛋白呼吸商从0.70到1.00的范围内,氧热价也过变动于19.6-21.1之间。此法在实际工作中是可用的。
另一种更简便的简略法只利用肺量计测出受试者一定时间内(通常为6min)的耗氧量。受试者一般都吃混合膳食,所以通常将非蛋白呼吸商定为0.82,氧热价为20.20kJ。因此,测出一定时间内的耗氧量后,使可依下式来计算:
产热量=20.20×耗氧量(kJ)
二、影响能量代谢的因素
影响能量代谢的因素有肌肉活动、精神活动、食物的特殊动力作用和环境温度等。
(一)肌肉活动
肌肉活动对能量代谢的影响最为显着。机体任何轻微的活动都可提高代谢率。人在运动或劳动时耗量显着增加,因为肌肉活动需要补给能量,而能量则来自大量营养物质的氧化,导致机体耗氧量的增加。机体耗氧量的增加与肌肉活动的强度呈正比关系,耗氧量最多右达安静时的10-20倍。肌肉活动的强度称为肌肉工作的强度,也就是劳动强度。劳动强度通常用单位时间内机体的产热量来表示,也就是说,可以把能量代谢率作为评估劳动强度的指标。从表7-3可以看出劳动强度或运动时的能量代谢率的增长情况。
表7-3 运动或劳动时的能量代谢率
| 机体的状态 | 产 热 量 |
| 平均(kJ/m2·min) | |
| 躺卧 | 2.73 |
| 开会 | 3.40 |
| 擦窗子 | 8.30 |
| 洗衣 | 9.98 |
■[此处缺少一些内容]■
由于随之出现的无意识的肌紧张以及刺激代谢的激素释放增多等原因,产热量可以显著增加。因此,在测定基础代谢率时,受试者必须摒除精神紧张的影响。
(三)食物的特殊动力作用
在安静状态下摄入食物后,人体释放的热量比摄入的食物本身氧化后所产生的热量要多。例如摄入能产100kJ热量的蛋白质后,人体实际产热量为130kJ,额外多产生了30kJ热量,表明进食蛋白质后,机体产热量超过了蛋白质氧化后产热量的30%。食物能使机体产生“额外”热量的现象称为食物的特殊动力作用(specific dynamic action)。糖类或脂肪的食物特殊动力作用为其产热量的4%-6%,即进食能产100kJ热量的糖类或脂肪后,机体产热量为104-106kJ。而混合食物可使产热量增加10%左右。这种额外增加的热量不能被利用来作功,只能用于维持体温。因此,为了补充体内额外的热量消耗,机体必须多进食一些食物补充这份多消耗的能量。
食物特殊动力作用的机制尚未完全了解。这种现象在进食后1h左右开始,并延续到7-8h。有人将氨基酸注入静脉内,可出现与经口给予相同的代谢率增值现象,这些事实使人们推想,食后的“额外”热量可能来源于肝处理蛋白质分解产物时“额外”消耗的能量。因此,有人认为肝在接脱氨基反应中消耗了能量可能是“额外”热量产生的原因。
(四)环境温度
人(裸体或只着薄衣)安静时的能量代谢,在20-30℃的环境中最为稳定。实验证明,当环境温度低于20℃时,代谢率开始有所增加,在10℃以下,代谢率便显着增加。环境温度低时代谢率增加,主要是由于寒冷刺激反射地引起寒战以及肌肉紧张增强所致。在20-30℃时代谢稳定,主要是由于肌肉松驰的结果。当环境温度为30-45℃时,代谢率又会逐渐增加。这可能是因为体内化学过程的反应速度有所增加的缘故,这时还有发汗功能旺盛及呼吸、循环功能增强等因素的作用。
三、基础代谢
基础代谢(basal metabolism)是指基础状态下的能量代谢。基础代谢率(basal metabolic rate,BMR)是指单位时间内的基础代谢,即在基础状态下,单位时间内的能量代谢。所谓基础状态是指人体处在清醒而又非常安静、不受肌肉活动、环境温度、食物及精神紧张等因素的影响时的状态。测定基础代谢率,要在清晨末进餐以前(即食后12-14h)进行。前一日晚餐最好是清淡菜肴,而且不要吃得太饱,这样,过了12-14h ,胃肠的消化和吸收活动已基本完毕,也排除了食物的特殊动力作用的影响。测定之前不应做剧烈的活动,而且必须静卧半小时以上。测定时平卧,全身肌肉要松驰,尽量排除肌肉活动的影响。这时还应要求受试者排除精神紧张的影响,如摒除焦虑、烦恼、恐惧等心理活动。室温要保持在20-25℃之间,以排除环境温度的影响。基本条件下的代谢率,比一般安静时的代谢率可低些(比清醒安静时低8%-10%)。基础代谢率以每小时,每平方米体表面积的产热量为单位,通常以kJ/m2·h来表示。要用每平方米体表面积而不用每公斤体重的产热量来表示,是因为基础代谢率的高低与体重并不成比例关系,而与体表面积基本上成正比。
表7-4 人体产热量和体表面积的关系
| 测定人数 | 平均体重(kg) | 体重范围 | kJ/m2·24h |
| 6 | 48.7 | 40-50 | 3860 |
| 41 | 53.4 | 50-60 | 3827 |
| 164 | 64.5 | 60-70 | 3881 |
| 24 | 74.7 | 70-80 | 3868 |
| 8 | 83.7 | 80-90 | 3868 |
| 平均 | 3860 |
若以每公斤体重的产热量进行比较,则小动物每公斤体重的产热量要比大动物高得多。若以每平方米体表面积的产热量进行比较,则不论机体的大小,各种动物每平方米每24小时的产热量很相近。在人体上测定的结果也表明了这一事实(表7-4)。因此,用每平方米体表面积标准来衡量能量代谢是比较合适的。
受试者体表面积的测定繁琐而不易进行,鉴于体表面积与身高、体重之间有一定的相关关系,因此,有人对一定的人群作过测定后,从身高、体重推算出体表面计算的经验公式。最基本的是Meeh的算式:
S=KW2/3
式中,S为体表面积,W为体重(kg),K为不同种属动物的常数。
计算人的体表面积在DuBois的身长体重算式:
S=W0.425×H0.725×K
式中,S为体表面积(m2),W为体重(kg),H为身长。K为不同人种的常数。我国人的体表面积可根据下列Stevenson算式来计算:
体表面积(m2)=0.0061×身长(cm)+0.0128×体重(kg)-0.1529
另外,体表面积还可根据图7-3直接求出。其用法是,将受试者的身高和体重在相应两条列线的两点连成一直线,引直线与中间的体表面积列线的交点就是该人的体表面积。有意义的事实是:肺活量、心输出量、主动脉和气管的横截面、肾小球滤过率等都与体表面积有一定的比例关系。]
通常采用前述简略法来测定和计算基础代谢率。下面举一实例供参考。
某受试者,男性,20岁。在基础状态下,1h的耗氧量为15L。非蛋白呼吸商定为0.82,氧热价为20.20,所以1h的产热量为:
20.20×15=303kJ
此人的体表面积为1.5m2,从而1h内每平方米体表面积拉热量,即基本代谢率为:303÷1.5=201.8kJ/m2·h
20岁男子的正常基础代谢率为157.8kJ/m2·h,所以此人超出正常值的数字为(201.8-157.8)=44.0,超出正常值的百分数为:
44.0×100/157.8%=27.8%
即+27.8%

图7-3 体表面积测算用图
实际测定结果表明,基础代谢率随性别、年龄等不同而有生理变化。当甚情况相同时,男子的基础代谢率平均比女子的高;幼年人比成年人的高;年龄越大,代谢率越低,但是,同一个体的基础代谢率,只在测定时的条件完全符合前述的要求,则有不同时日重复测定的结果基本上无差异。这就反映了正常人的基础代谢率是相当稳定的。
关于我国正常人基础代谢率的水平,男女各年龄组的平均值如表7-5所示。
一般来说,基础代谢率的实际数值现上述正常的平均值比较,相差±10%-15%之内,无论较高或较低,都不属病态。当相差之数超过20%时,才有可能是病理变化。在各种疾病中,甲状腺功能的改变总是伴有基础代谢率异常变化。甲状腺功能低下时,基础代谢率将比正常值低20%-40%;甲状腺功能亢进时的基础代谢率将比正常值高出25%-80%(图7-4)。因此,基础代谢率的测量是临床诊断甲状腺疾病的重要辅助方法。其它如肾上腺皮质和垂体的功能低下时,基础代谢率也要降低。
表7-5 我国人正常的基础代谢率平均值(kJ/m2·h)
年龄 11-15 16-17 18-19 20-30 31-40 41-50 51 以上
男性 195.5 193.4 166.2 157.8 158.7 154.1 149.1
女性 172.5 181.7 154.1 146.4 142.4 142.4 138.6
当人体发热时,基础代谢率将升高。一般说来,体温每升高1℃,基础代谢率可升高13%。其它如糖尿病、红细胞增多症、白血病以及伴有呼吸困难的心脏病等,也伴有基础代谢率升高。当机体处于病理性饥饿时,基础代谢率将降低。其他如阿狄森病、肾病综合症以及垂体肥胖症也常伴有基础代谢率降低。
图7-4 甲状腺疾病患者的基础代谢与正常人基础代谢的比较
纵坐标表示受检人数;横坐标的0代表正常平均值,
-10表示比正常人平均值低10%,10表示高10%
第二节 体温及其调节
一、体温
人和高等动物机体都具有一定的温度,这就是体温。体温是机体进行新陈代谢和正常生命活动的必要条件。
(一)表层体温和深部体温
人体的外周组织即表层,包括皮肤、皮下组织和肌肉等的温度称为表层温度(shelltemperature)。表层温度不稳定,各部位之间的差异也不大。在环境温度为23℃时,人体表层最外层的皮肤温,如足皮肤温为27℃,手皮肤温为30℃。躯干为32℃,额部为33-34℃。四肢末稍皮肤温最低,越近躯干、头部,皮肤温越高。气温达32℃以上时,皮肤温的部位差将变小,在寒冷环境中,随着气温下降,手、足的皮肤温降低最显着,但头部皮肤温度变动相对较小。
皮肤与局部血流量有密切关系。凡是能影响皮肤血管舒缩的因素(如环境温度变化或精神紧张等)都能改变皮肤的温度。在寒冷环境中,由于皮肤血管收缩,皮肤血流量减少,皮肤温随之降低,体热散失因此减少。相反,在炎热环境中,皮肤血管舒张,皮肤血流量增加,皮肤温因而上升,同时起到了增强发散体热的作用。人情绪激动时,由于血管紧张度增加,皮肤温、特别是手的皮肤温便显著降低。例如手指的皮肤温可从30℃骤降到24℃。当然情绪激动的原因解除后,皮肤温会逐渐恢复。此外,当发汗时由于蒸发散热,皮肤温也会出现波动。
机体深部(心、肺、脑和腹腔内脏等处)的温度称为深部温度(coretemperature)。深部温度比表层温度高,且比较稳定,各部位之间的差异也较小。这里所说的表层与深部,不是指严格的解剖学结构,而是生理功能上所作的体温分布区域。在不同环境中,深部温度和表层温度的分布会发生相对改变。在较寒冷的环境中,深部温度分布区域较缩小,主要集中在头部与胸腹内脏,而且表层与深部之间存在明显的温度梯度。在炎热环境中,深部温度可扩展到四肢(图7-5)。
图7-5 在不同环境温度下人体体温分布图
A: 环境温度20℃ B:环境温度35℃
体温是指机体深部的平均温度。由于体内各器官的代谢水平不同,它们的温度略有差别,但不超过1℃。在安静时,肝代谢最活跃,温度最高;其次,是心脏和消化腺。在运动时则骨骼肌的温度最高。循环血液是体内传递热量的重要途径。由于血液不断循环,深部各个器官的温度会经常趋于一致。因此,血液的温度可以代表重要器官温度的平均值。
临床上通常用口腔温度、直肠温度和腋窝温度来代表体温。直肠温度的正常值为36.9-37.9℃,但易受下肢温度影响。当下肢冰冷时,由于下肢血液回流至髂静脉时的血液温度较低,会降低直肠温度;口腔温度(舌下部)平均比口腔温度低0.3℃,但它易受经口呼吸、进食和喝水等影响;腋窝温度平均比口腔温度低0.4℃.但由于腋窝不是密闭体腔,易受环境温度、出汗和测量姿势的影响,不易正确测定。
此外,食管温度比直肠温度约低 0.3℃。食管中央部分的温度与右心的温度大致相等,而且体温调节反应的时间过程与食管温度变化过程一致。所以,在实验研究中,食管温度可以作为深部温度的一个指标。鼓膜温度的变动大致与下丘脑温度的变化成正比,所以在体温调节生理实验中常常用鼓膜温度作为脑组织温度的指标。
(二)体温的正常变动
在一昼夜之中,人体体温呈周期性波动。清晨2-6时体温最低,午后1-6时最高。波动的幅值一般不超过1℃。体温的这种昼夜周期性波动称为昼夜节律或日周期(circadian rhythm)。
女子的基础体温随月经周期而发生变动。在排卵后体温升高,这咱体温升高一直持续至下次月经开始(图7-6)。这种现象很可能同性激素的分泌有关。实验证明,这种变动性同血中孕激素及其代谢产物的变化相吻合。
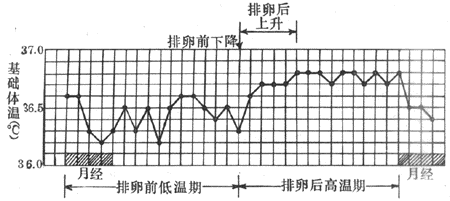
图7-6 女子的基础体温曲线
体温也与年龄有关。一般说来,儿童的体温较高,新生儿和老年人的体温较低。新生儿,特别是早产儿,由于体温调节机制发育还不完善,调节体温的能力差,所以他们的体温容易受环境温度的影响而变动。因此对新生儿应加强护理。
肌肉活动时代谢加强,产热量因而增加,结果可导致体温升高。所以,临床上应让病人安静一段时间以后再测体温。测定小儿体温时应防止哭闹。
此外,情绪激动、精神紧张、进食等情况对体温都会有影响;环境温度的变化对体温也有影响;在测定体温时,就考虑到这些情况。
二、体热平衡
如第一节所述,机体内营养物质代谢释放出来的化学能,其中50%以上以热能的形式用于维持体温,其余不足50%的化学能则载荷于ATP,经过能量转化与利用,最终也变成热能,并与维持体温的热量一起,由循环血液传导到机体表层并散发于体外。因此,机体在体温调节机制的调控下,使产热过程和散热过程处于平衡,即体热平衡,维持正常的体温。如果机体的产热量大于散热量,体温就会升高;散热量大于产热量则体温就会下降,直到产热量与散热量重新取得平衡时才会使体温稳定在新的水平。
(一)产热过程
机体的总产热量主要包括基础代谢,食物特殊动力作用和肌肉活动所产生的热量。基础代谢是机体产热的基础。基础代谢高产热量多;基础代谢低,产热量少。正常成年男子的基础代谢率约为170kJ/m2·h。成年女子约155kJ/m2·h在安静状态下,机体产热量一般比基础代谢率增高25%,这是由于维持姿势时肌肉收缩所造成的。食物特殊动力作用可使机体进食后额外产生热量。骨骼肌的产热量则变化很大,在安静时产热量很小。运动时则产热量很大;轻度运动如平行时,其产热量可比安静时增加3-5倍,剧烈运动时,可增加10-20倍。
人在寒冷环境中主要依靠寒战来增加产热量。寒战是骨骼肌发生不随意的节律性收缩的表现,其节律为9-11次/分。发生寒战的肌肉在肌电图上表现出一簇一簇的高波幅群放电,这是不同肌纤维的动作电位同步化的结果。寒战的特点是屈肌和伸肌同时收缩,所以基本上不做功,但产热量很高,发生寒战时,代谢率可增加4-5倍。机体受寒冷刺激时,通常在发生寒战之前,首先出现温度刺激性肌紧张(thermalmuscle tone)或称寒战前肌紧张(pre-shivering tone),此时代谢率就有所增加。以后由于寒冷刺激的持续作用,便在温度刺激性肌紧张的基础上出现肌肉寒战,产热量大大增加,这样就维持了在寒冷环境中的体热平衡。内分泌激素也可影响产热,肾上腺素和去甲肾上腺素可使产热量迅速增加,但维持时间短;甲状腺激素则使产热缓慢增加,但维持时间长。机体在寒冷环境中度过几周后,甲状腺激素分泌可增加2倍能上能下,代谢率可增加20-30%。
(二)散热过程
人体的主要散热部位是皮肤。当环境温度低于体温时,大部分的体热通过皮肤的辐射、传导和对流散热。一部分热量通过皮肤汗液蒸发来散发,呼吸、排尿和排粪也可散失一小部分热量(表7-6)。
表7-6 在环境温度为21℃时人体散热方式及其所占比例
| 散热方式 | 百 分 数 (%) |
| 辐射、传导、对流 | 70 |
| 皮肤水分蒸发 | 27 |
| 呼吸 | 2 |
| 尿、粪 | 1 |
1.辐射、传导和对流散热
辐射(radiation)散热:这是机体以热射线的形式将热量传给外界较冷物质一种散热形式。以此种方式散发的热量,在机体安静状态下所占比例较大(约占部散热量的60%左右)。辐射散热量同皮肤与环境间的温度差以及机体有效辐射面积等因素有关。皮肤温稍有变动,辐射散热量就会有很大变化。四肢表面积比较大,因此在辐射散热中有重要作用。气温与皮肤的温差越大,或是机体有效辐射面积越大,辐射的散热量就越多。
传导(conduction)和对流(convection)散热:传导散热是机体的热量直接传给同它接触的较冷物体的一种散热方式。机体深部的热量以传导方式传到机体表面的皮肤,再由后者直接传给同它相接触的物体,如床或衣服等。但由于此等物质是热的不良导体,所以体热因传导而散失的量不大。另外,人体脂肪的导热度也低,肥胖者皮下脂肪较多,女子一般皮下脂肪也较多,所以,他们由深部向表层传导的散热量要少些。皮肤涂油脂类物质,也可以起减少散热的作用。水的导热度较大,根据这个道理可利用冰囊、冰帽给高热病人降温。
对流散热是指通过气体或液体或交换热量的一种方式。人体周围总是绕有一薄层同皮肤接触的空气,人体的热量传给这一层空气,由于空气不断流动(对流),便将体热发散到空间。对流是传导散热的一种特殊形式。通过对流所散失的热量的多少,受风速影响极大。风速越大,对流散热量也越多,相反,风速越小,对流散热量也越少。
辐射、传导和对流散失的热量取决于皮肤和环境之间的温度差,温度差越大,散热量越多,温度差越小,散热量越少。皮肤温度为皮肤血流量所控制。皮肤血液循环的特点是,分布到皮肤的动脉穿透隔热组织(脂肪组织等),在乳头下层形成动脉网;皮下的毛细血管异常弯曲,进而形成丰富的静脉丛;皮下还有大量的动-静脉吻合支,这些结构特点决定了皮肤的血流量可以在很大范围内变动。机体的体温调节机制通过交感神经系统控制着皮肤血管的口径。增减皮肤血流量以改变皮肤温度,从而使散热量符合于当时条件下体热平衡的要求。
在炎热环境中,交感神经紧张度降低,皮肤小动脉命张,动-静脉吻合支开放,皮肤血流量因而大大增加(据测算,全部皮肤血流量最多可达到心输出量的12%)。于是较多的体热从机体深部被带到体表层,提高了皮肤温,增强了散热作用。
在寒冷环境中,交感神经紧张度增强,皮肤血管收缩,皮肤血流量剧减,散热量也因而大大减少。此时机体表层宛如一个隔热器,起到了防止体热散失的作用。此外,四肢深部的静脉是和动脉相伴走行的。这样的解剖结构相当于一个热量逆流交换系统。深部静脉呈网状围绕着动脉。静脉血温较低,而动脉血温度较高。两者之间由于温度差而进行热量交换。逆流交换的结果,动脉血带到末稍的热量,有一部分已被静脉血带回机体深部。这样就减少了热量的散失。如果机体处于炎热环境中,从皮肤返回心脏的血液主要由皮肤表层静脉来输送,此时逆流交换机制将不再起作用(图7-7)。
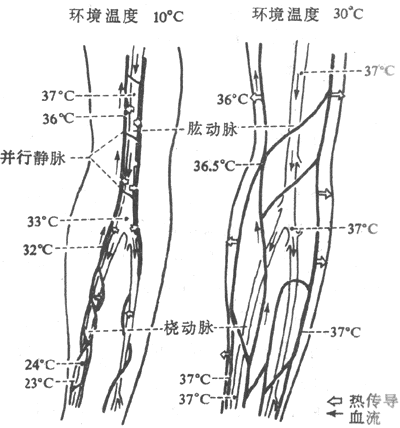
图7-7 上肢的逆流热量交换
环境温度降低时,热量由肱动脉传向它周围的静脉,
动脉血温度因此下降到24℃。环境温度升高时,热量由表层静脉发散
衣服覆盖的皮肤表层,不易实现对流,棉毛纤维间的空气不易流动,这类情况都有利于保温。增加衣着以御寒,就是这个道理。
2.蒸发散热 在人的体温条件下,蒸发(evaporation)1g水分可使机体散失2.4kJ热量。当环境温度为21℃时,大部分的体热(70%)靠辐射、传导和对流的方式散热,少部分的体热(29%)则由蒸发散热;当环境温度升高时,皮肤和环境之间的温度差变小,辐射、传导和对流的散热量减小,而蒸发的散热作用则增强;当环境温度等于或高于皮肤温度时,辐射、传导和对流的散热方式就不起作用,此时蒸发就成为机体唯一的散热方式。
人体蒸发有二种形式:即不感蒸发(insesibleperspiration)和发汗(sweating)。人体即使处在低温中,没有汁液分泌时,皮肤和呼吸道都不断有水分渗也而被蒸发掉,这种水分蒸发称为不感蒸发,其中皮肤的水分蒸发又称为不显汗,即这种水分蒸发不为人们所觉察,并与汁腺的活动开关。在室温30℃以下时,不感蒸发的水分相当恒定,有12-15g/h·m2水分被蒸发掉,其中一半是呼吸道蒸发的水分;另一半的水分是由皮肤的组织间隙直接渗出而蒸发的。人体24h的不感蒸发量为400-600ml。婴幼儿的不感蒸发的速率比成从大,因此,在缺水时婴幼儿更容易造成严重脱水。不感蒸发是一种很有成效的散热途径,有些动物如狗,虽有汁腺结构,但在高温环境下也不能分泌汁液,此时,它必须通过热喘呼吸(panting)由呼吸道来增强蒸发散热。
发汗汗腺分泌汁液的活动称为发汗。发汗是可以意识到的有明显的汗液分泌,因此,汁液的蒸发又称为可感蒸发。
人在安静状态下,当环境温度达30℃左右时便开始发汗。如果空气湿度大,而且着衣较多时,气温达25℃便可引起人体发汗。人进行劳动或运动时,气温虽在20℃以下,亦可出现发汗,而且汗量往往较多的。
汗液中水分占99%,而固体成分则不到1%,在固体成分中,大部分为氯化钠,也有少量氯化钾、尿素等。同血浆相比,汗液的特点是:氯化钠的浓度一般低于血浆;在高温作业等大量出汗的人,汗液中可丧失较多的氯化钠,因此应注意补充氯化钠。汗液中葡萄糖的浓度几乎是零;乳酸浓度主于血浆;蛋白质的浓度为零。实验测得在汗腺分泌时,分泌管腔内的压力高达37.3kPa(250mmHg)以上。这表明汗液不是简单的血浆滤出液,而是由汗腺细胞主动分泌的。大量的乳酸是腺细胞进入分泌活动的产物。刚刚从汗腺细胞分泌出来的汗液,与血浆是等渗的,但在流经汗腺管腔时,由于钠和氯被重吸收,所以,最后排出的汗液是低渗的。汗液中排出的钠量也受醛固醇的调节。下因为汗液是低渗的,所以当机体因大量发汗而造成脱水时,可导致高渗性脱水。
发汗是反射活动。人体汗腺接受交感胆碱能纤维支配,所以乙酰胆碱对小汗腺有促进分泌作用。发汗中枢分布在从脊髓到大脑皮层的中枢神经系统中。在正常情况下,起主要作用是是下丘脑的发汗中枢,它很可能位于体温调节中枢之中或其附近。
在温热环境下引起全身各部位的小汗腺分泌汗液称为温热性发汗。始动温热性发汗的主要因素有:①温热环境刺激皮肤中的瘟觉感受器,冲动传入至发汗中枢,反射性引起发汗;②温热环境使皮肤血液被加温,被加温的血液流至下丘脑发汗中枢的热敏神经元,可引起发汗。温热性发汗的生理意义在于散热。若每小时蒸发1.7L汗液,就可使体热散发约4200kJ的热量。但是,如果汗水从身上滚落或被擦掉而未被蒸发,则无蒸发散热作用。
发汗速度受环境温度和湿度影响。环境温度越高,发汗速度越快。如果在高温环境中时间太长,发汗速度会因汗腺疲劳而明显减慢。湿度大,汗液不易被蒸发,体热因而不易蒸发,体热因而不易散失。此外,风速大时,汗液易蒸发,汗液蒸发快,容易散热而使发汗速度变小。
劳动强度也影响发汗速度。劳动强度大,产热量越多,发汗量越多。
精神紧张或情绪激动而引起地发汗称为精神性发汗。主要见于掌心、脚底和腋窝。精神性发汗的中枢神经可能在大脑皮层运动区。精神性发汗在体温调节中的作用不大。
三、体温调节
恒温动物包括人,有完善的体温调节机制。在外界环境温度改变时,通过调节产热过程和散热过程,维持体温相对稳定。例如,在寒冷环境下,机体增中产热和减少散热;在炎热环境下,机体减少产热和增加散热,从而使体温保持相对稳定。这是复杂的调节过程,涉及感受温度变化的温度感觉器,通过有关传导通路把温度信息传达到体温调节中枢,经过中枢整合后,通过自主神经系统调节皮肤血流量、竖毛肌和汗腺活动等;通过躯体神经调节骨骼肌的活动,如寒战等;通过内分泌系统,改变机体的代谢率。
体温调节是生物自动控制系统的实例。如图7-8所示,下丘脑体温调节中枢,包括调定点(set point)神经元在内,属于控制系统。它的传出信息控制着产热器官如肝、骨骼肌以及散热器官如皮肤血管、汗腺等受控系统的活动,使受控对象——机体深部温度维持一个稳定水平。而输出变量体温总是会受到内、旬环境因素干扰的(譬如机体的运动或外环境气候因素的变化,如气温、湿度、风速等)。此时则通过温度检测器——皮肤及深部温度感受器(包括中枢温度感受器)将干扰信息反馈于调定点,经过体温调节中枢的整合,再调整受控系统的活动,仍可建立起当时条件下的体热平衡,收到稳定体温的效果。
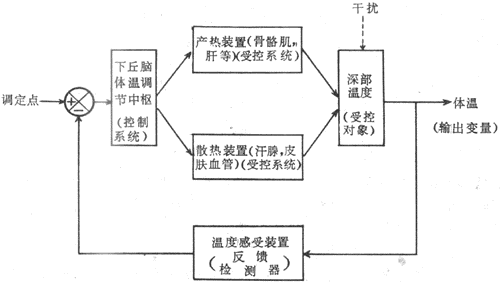
图7-8 体温调节自动控制示意图
(一)温度感受器
对温度敏感的感受器称为温度感受器,温度感受器分为外周温度感受器和中枢温度感受器。
外周温度感受器在人体皮肤、粘膜和内脏中,温度感受器分为冷感受器和温觉感受器,它们都是游离神经末稍的。当皮肤温度升高时,温觉感受器兴奋,而当皮肤温度下降时,则冷感受器兴奋。从记录温度感受器发放冲动可看到,温觉感受器和冷觉感受器在28℃时发放冲动频率最高,而温觉感受器则在43℃时发放冲动频率最高。当皮肤温度偏离这两个温度时,两种感受器发放冲动的频率都逐渐下降(图7-9)。此外,温度感受器对皮肤温度变化速率更敏感。
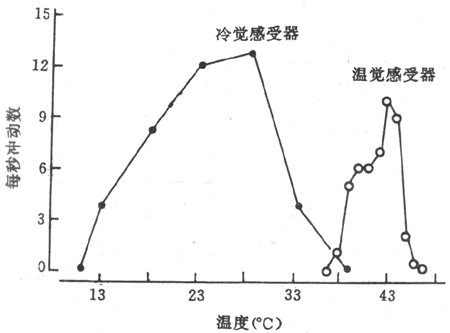
图7-9 大鼠阴囊皮肤冷觉和温觉感受器
内脏器官也有温度感受器。有人将电热器埋藏在差点羊腹腔内 并加温至43-44℃,观察到羊的呼吸频率和蒸发散热迅速增加,加热3-5分钟后,动物开始喘息,使下丘脑温度下降。说明内脏温度升高可引起明显的散热反应。
中枢温度感受器在脊髓、延髓、脑干网状结构及下丘脑中有温度感受器。
用改变脑组织温度的装置(变温管),对不麻醉或麻醉的兔、猫或狗等的下丘脑前部进行加温或冷却,发现在视前区-下丘脑前部(preoptic anterior hypothalamus,PO/AH)加温,可引起动物出现喘息和出汗等散热反应,而局部冷却则引起产热量增加,说明PO/AH本身就可调节散热和产热这两种相反的过程。用电生理方法记录PO/AH中存在着热敏神经元(warm-sensitive neuron)和冷敏神经元(cold-sensitiveneuron)。前者的放电频率随局部温度的升高而增加,而后者的放电频率则随着脑组织的降温而增加(图7-10,7-11)。实验证明,局部脑组织温度变动0.1C,这两种温度敏感神经元的放电频率就会反映出来,而且不出现适应现象。
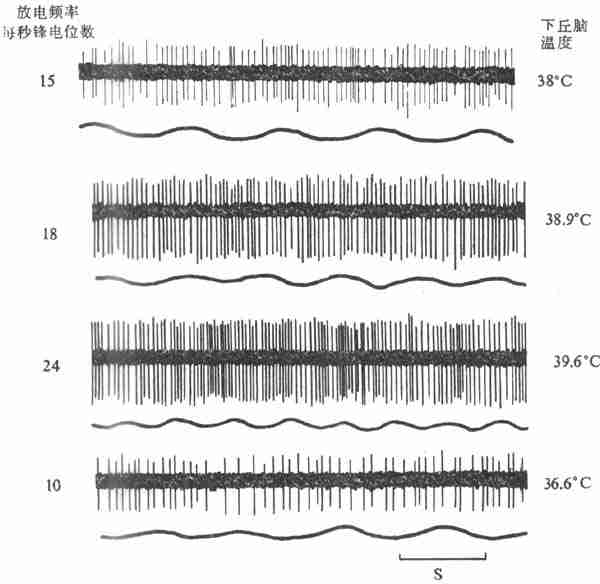
图7-10下丘脑局部加温时热敏神经元放电的记录(上)和呼吸曲线(下)(猫)
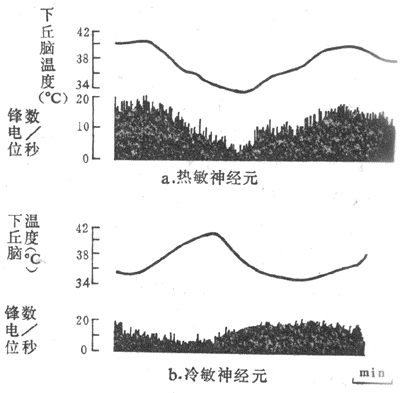
图7-11由下丘脑视前区导出的温度敏感神经元的放电活动
脊髓中也有温度敏感神经元。冷却轻度麻醉狗的颈、胸髓或胸腰髓,则动物出现皮肤血管收缩和寒战等体温调节反应。这时,切断被冷却部位的后根或高位切断脊髓,血管反应和寒战也不消失。加温脊髓,则引起皮肤血管舒张和热喘呼吸,寒战受到抑制。另外,据谓脊髓中传导温度信息的上行性神经元的纤维前侧侧索中走行,它将信息发送给PO/AH。
延髓中也存在着温度敏感神经元。皮肤、脊髓及中脑的传入温度信息都会聚于延髓温度敏感神经元;而延髓也接受来自PO/AH的信息,并且向PO/AH输送信息。
脑干网状结构也有对局部温度变化发生反应的神经元,它接受发生皮肤、脊髓的温度信息,并且向PO/AH输送温度信息。
(二)体温调节中枢
根据多种恒温动物脑的分段切除实验看到,切除大脑皮层及部分皮层下结构后,只要保持下丘脑及其以下的神经结构完整,动物虽然在行为方面可能出现一些欠缺,但仍具有维持恒定体温的能力。如进一步破坏下丘脑,则动物不再具有维持体温相对恒定的能力。这些事实说明,调节体温的基本中枢在下丘脑。下丘脑局部破坏或电刺激等实验观察到,PO/AH破坏,则散热反应消失,体温升高;刺激之,则引起散热反应,而且寒战受到抑制;而破坏下丘脑后部,体温下降,产热反庆受抑制;刺激之,则引起寒战。据此得出结论,下丘脑前部是散热中枢,而下丘脑后部是产热中枢,但是,这两种实验方法比较粗糙,因此得出来的结论也较精细的实验方法观察到的结果不相符。
前已述及, PO/AH就有热敏神经元和冷敏神经元,分别调节散热和产热反应。下丘脑以外的脑细胞也有类似的两种神经元存在。看来没有明确定位的产热中枢和散热中枢。体温调节是涉及多方输入温度信息和多系统的传出反应,因此是一种高级的中枢整合作用。视前区-下丘脑前部应是体温调节的基本部位。下丘脑前部的热敏神经元和冷敏神经元既能感受它们所在部位的温度变化,又能过传入的温度信息进行整合。因此,当外界环境温度改变时,可通过①皮肤的温、冷觉感受器的刺激,将温度变化的信息沿躯体传入神经经脊髓到达下丘脑的体温调节中枢;②外界温度改变可通过血液引起深部温度改变,并直接作用于下丘脑前部;③脊髓和下丘脑以外的中枢温度感受器也将温度信息传给下丘脑前部。通过下丘脑前部和中枢其它部位的整合作用,由下述三条途径发出指令调节体温:①通过交感神经系统调节皮肤血管舒缩反应和汗腺分泌;②通过躯体神经改变骨骼肌的活动,如在寒冷环境时的寒战等;③通过甲状腺和肾上腺髓质的激素分泌活动的改变来调节机体的代谢率。有人认为,皮肤温度感受器兴奋主要调节皮肤血管舒活动和血流量;而深部温度改变则主要调节发汗和骨骼肌的活动。通过上述的复杂调节过程,使机体在外界温度改变时能维持体温相对稳定。
调定点学说 此学说认为,体温的调节类似于恒温器的调节,PO/AH中有个调定点,即规定数值(如37℃)。如果偏离此规定数值,则由反馈系统将偏离信息输送到控制系统,然后经过对受控系统的调整来维持体温的恒定。通常认为,PO/AH中的温度敏感神经元可能在体温调节中起着调定点的作用。例如,此学说认为,由细菌所致的发热是由于热敏神经元的阈值因受到热原(pyrogen)的作用而升高,调定点上移(如39℃)的结果。因此,发热反应开始先出现恶寒战栗等产热反应,直到体温升高到39℃以上时才出现散热反应。只要致热因素不消除,产热与散热两个过程就继续在此新的体温水平上保持着平衡。应该指出的是,发热时体温调节功能并无阻碍,而只是由于调定点上移,体温才被调节到发热水平。
单胺物质对体温调节的作用在哺乳动物下丘脑的与体温调节有关的神经末稍中含有丰富的单胺物质。60年代初,用狗、猫、猴做的实验证明,用5-羧色胺灌注动物的脑室或微量注入于下丘脑,动物的体温上升,同时伴有血管收缩反应和寒战;而去甲肾上肾素则使动物的体温降低0.5-2℃,同时伴有外周血管舒张。根据这类实验,提出了体温调节的单胺学说,此学说认为,5-羧色胺和去甲肾上腺素这两种物质在量上的动态平衡可保持体温的恒定。但目前认为,这两种物质对体温调节中枢的活动只能起到调整的作用,而对于体温的恒定水平没有决定作用。
参考资料
1.徐丰彦,张镜如主编.人体生理学.北京;人民卫生出版社,1989
2.Patton HD,FuchsAE,Hille B,Scher RS,Textbook of Physiology 21th ed,WB SaundersCO,Philadelphia,1989
3.Ganong WF.Review of MedicalPhysiology 13thed,Lange Medical Publications,LosAltos,Calfornia,1991
4.Emslie-Smith D,PatersonCR,Scratcherd T,Read NW.Textbook of Physiology,Scratchill Livingstone EdinburghLondon Melbourne and New York,1988
5.Mountcastle VB,MedicalPhysiology 14thed,CV Mosby Co,St Louis 1980
第八章 尿的生成和排出
肾是维持机体内环境相对稳定的最重要的器官之一。通过尿的生成和排出,①排除机体的大部分代谢终产物以及进入体内的异物;②调节细胞外液量和渗透压;③保留体液中的重要电解质如钠、钾、碳酸氢盐以及氯离子等,排出氢离子,维持酸碱平衡。
尿的生成包括肾小球的滤过,肾小管和集合管的重吸收以及它们的分泌三个基本过程。
本章主要阐述肾的尿生成过程及其调节机制,以及输尿管和膀胱的排尿活动。
第一节 肾的功能解剖和肾血流量
一、肾的功能解剖
(一)肾单位和集合管
肾单位是肾的基本功能单位,它与集合管共同完成泌尿功能。人的两侧肾约有170-240万个肾单位,每个肾单位包括肾小体和肾小管部分(图8-1)。肾小体包括肾小球和肾小囊两部分。肾小球是一团毛细血管网,其峡谷端分别与入球小动脉和出球小动脉相连(图8-2)。肾小球的包囊称为肾小囊。它有两层上皮细胞,内层(脏层)紧贴在毛细血管壁上,外层(壁层)与肾小管壁相连;两层上皮之间的腔隙称为囊腔,与肾小管管腔相通。血浆中某些成分通过肾小球毛细血管网向囊腔滤出;滤出时必须通过肾小球毛细血管内皮细胞、基膜和肾小囊脏层上皮细胞,这三者构成滤过膜(图8-3)。
图8-1 肾单位示意图
图8-2 肾小球、肾小囊穿刺和近球小体示意图方框示近球小体
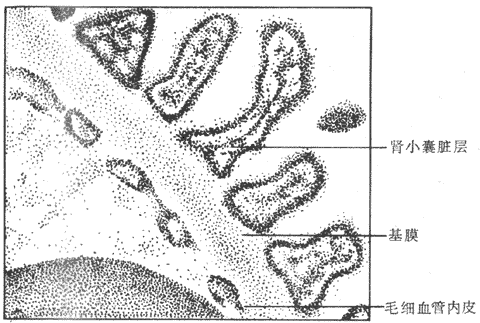
图8-3 滤过膜示意图
肾小管由近球小管、髓袢和远球小管三部分组成。近球小管包括近由小管和髓袢降支粗段。髓袢由髓袢降支和髓袢升支组成;前者包括髓袢降支粗段(也是近球小管的组成部分)和降支细段;后者是指髓袢升支细段和升支粗段(也是远球小管的一部分)。远球小管包括髓袢升支粗段和远曲小管。远曲小管末端怀集合管相连。
集合管不包括在肾单位内,但在功能上和远球小管密切相关,它在尿生成过程中,特别是在尿液浓缩过程中起着重要作用,每一集合管接受多条远内小管运来的液体。许多集合管又汇入乳头管,最后形成的尿液经肾盏、肾盂、输尿管而进入膀胱,由膀胱排出体外。
(二)皮质肾单位和近髓肾单位
肾单位按其所在部位不同,可分为皮质肾单位和近髓肾单位(髓旁肾单位)两类(图8-4)。
皮质肾单位主要分布于外皮质层和中皮质层。人肾的皮质肾单位约占肾单位总数的85%-90%。这类肾单位的肾小球体积较小;入球小动脉的口径比出球小动脉的粗,两者口径之比约为2:1。出球小动脉进一步再分为毛细血管后,几乎全部分布于皮质部分的肾小管周围。这类肾单位的髓袢甚短,只达外髓质层,有的甚至不到髓质。
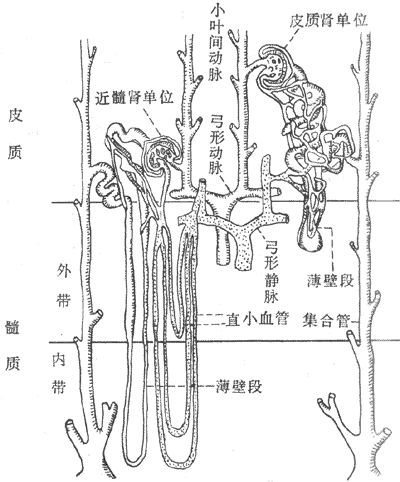
图8-4 肾单位和肾血管的示意图
处于肾皮质不同部位的肾单位和肾血管的结构显著不同
近髓肾单位分布于靠近髓质的内皮质层,在人肾约占肾单位中的10%-15%。这类肾单位的肾小球体积较大;其髓袢甚长,可深入到内髓质层,有的甚至到达乳头部。出球小动脉不仅形成缠绕邻近的近曲小管或远曲小管的网状毛细血管,而且还形成细而长的U字形直小血管。直小血管可深入到髓质,并形成毛细血管网包绕髓袢升支和集合管。近髓肾单位和直小血管的这些解剖特点,决定了它们在尿的浓缩与稀释过程中起着重要作用。
(三)近球小体
近球小体(juxtaglomerular apparatus)由颗粒细胞、系膜(间质)细胞和致密斑三者组成(图8-2)。颗粒细胞是位于入球小动脉的中膜内的肌上皮样细胞,内含分泌颗粒,分泌颗粒内含肾素。系膜细胞是指入球小动脉和出球小动脉之间的一群细胞,具有吞噬功能。致密斑位于远曲小管的起始部分,此处的上皮细胞变为主柱状细胞,局部呈现斑纹隆起,称为致密斑,致密斑与入球小动脉和出球小动脉相接触。致密斑可感受小管液中NaCI含量的变化,并将信息传递至颗粒细胞,调节肾素的释放。
近球小体主要分布在皮质肾单位,因而皮质肾单位含肾素较多,而近髓肾单位则几乎不含肾素。肾素分布的这种差异,也提示两种肾单位在功能上有所不同。
(四)肾的神经支配
肾交感神经主要从胸12至腰12脊髓发出,其纤维经腹腔神经丛支配肾动脉(尤其是入球小动脉和出球小动脉的不滑肌)、肾小管和释放肾素的颗粒细胞。肾交感神经末稍释放去甲肾上腺素,调节肾血流量、肾小球滤过率、肾小管的重吸收和肾素释放。有人认为,未发现肾有副并感神经支配,肾的各种感受器可经肾神经传入脊髓,并从脊髓投射到中枢的不同部位。
(五)肾的血液供应
肾动脉由腹主动脉垂直分出,其分支经叶间动脉→弓形动脉→小叶间动脉→入球小动脉。每支入球小动脉进入肾小体后,又分支成肾小球毛细血管网,后者汇集成出球小动脉而离开肾小体。出球小动脉再次分成毛细血管网,缠绕于肾小管和集合管的周围。所以,肾血液供应要经过两次毛细血管网,然后才汇合成静脉,由小叶间静脉→弓形静脉→叶间静脉→肾静脉(图8-4)。
肾小球毛细血管网介于入球小动脉和出球小动脉之间,而且皮质肾单位入球小动脉的口径比出球小动脉的粗1倍。因此,肾小球毛细血管内血压较高,有利于肾小球的滤过作用;肾小管周围的毛细血管网的血压较低,可促进肾小管的重吸收。
二、肾血液循环的特征
肾的血液供应很丰富。正常成人安静时每分钟有1200ml血液流过两侧肾,相当于心输出量的1/5-1/4左右。其中约94%的血液分布在肾皮质层,5%-6%分布在外髓,其余不到1%供应内髓,通常所说的肾备注量主要指肾皮质血流量。
肾血流量的调节包括肾血流量的自身调节和神经体液调节。
(一)肾血流量的自身调节
肾血流量的自身调节表现为动脉血压在一定范围内变动时,肾血流量仍然保持相对恒定(图8-5)。离体肾实验观察到,当肾动脉的灌注压(相当于体内的平均动脉压)由2.7kPa(20mmHg)提高到10.7kPa(80mmHg)的过程中,肾血流量将随肾灌注压的升高而成比例地增加;而当灌注压在10.7-247kPa(80-180mmHg)范围内变动时,肾血流量又将随灌注压的长高而增加。这种不依赖肾外神经支配使肾血流量在一定的血压变动范围内能保持不变的现象,表明它具有自身调节。一般认为,自身调节只涉及肾皮质的血流量。
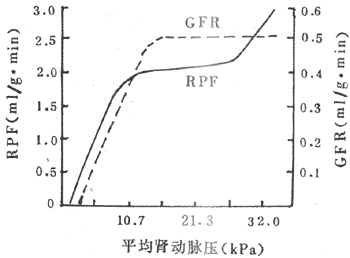
图8-5 肾血流量和肾小球滤过率的自身调节
RPF:肾血浆流量 GFR:肾小球滤过率
关于自身调节的机制,有人提出肌源学说来解释。此学说认为,当肾灌注压增高时,血管平滑肌因灌注压增加而受到牵张刺激,这使得不滑肌的紧张性加强,血管口径相应地缩小,血流的阻力便相应地增大,保持肾血流量稳定;而当灌注压减小时则发生相反的变化。由于在灌注压低于10.77kPa(80mmHg)时,平滑肌已达到舒张的极限;而灌注压高于247kPa(180mmHg)时,平滑肌又达到收缩的极限。因此,在10.7kPa(80mmHg)以下和24kPa(180mmHg)以上时,肾血流量的自身调节便不能维持,肾血流量将随血压的变化而变化。只有在10.7-24kPa(80-180mmHg)的血压变化范围内,入球小动脉平滑肌才能发挥自身调节作用,保持肾血流量的相对恒定。如果用罂粟碱、水合氯醛或氰化钠等药物抑制血管平滑肌的活动,自身调节便告消失。此外,还不能完全排除其它因素如肾内组织液压力、代谢产物等因素在肾血流量自身调节中的作用。通过肾血流量自身调节,使肾小球滤守率不会因血压波动而改变,维持肾小球滤过率相对恒定(图8-5)。
(二)肾血流量的神经和体液调节
肾血流量的神经、体液调节使肾血流量与全身的血液循环调节相配合。肾交感神经活动加强时,引起肾血管收缩,肾血流量减少。影响肾交感神经活动的因素已在心血管反射中作了详述(见第四章)。
肾上腺素与去甲肾上腺素都能使肾血管收缩,肾血流量减少。血管升压素和血管紧张素等也能使肾血管收缩;前列腺素可使肾血管扩张。
总之,在通常情况下,在一般的血压变化范围内,肾主要依靠自身调节来保持血流量的相对稳定,以维持正常的泌尿功能。在紧急情况下,全身血液将重新分配,通过交感神经及肾上腺素的作用来减少肾血流量,使血液分配到脑、心脏等重要器官,这对维持脑和心脏的血液供应有重要意义。
第二节 肾小球的滤过功能
循环血液经过肾小球毛细血管时,血浆中的水和小分子溶质,包括少量分子量较小的血浆蛋白,可以滤入肾小囊的囊腔而形成滤过液。用微穿刺法实验证明,肾小球的滤过液就是血浆中的超滤液。
微穿刺法是利用显微操纵仪将外径6-10μm的微细玻璃插入肾小体的囊腔中。在与囊腔相接部位的近球小管内,注入石蜡油防止起滤液进入肾小管。用微细玻璃管直接抽到囊腔中的液体进行微量化学分析(图8-2)。分析表明,除了蛋白质含量甚少之外,各种晶体物质如葡萄糖、氯化物、无机磷酸盐、尿素、尿酸和肌酐等的浓度都与血浆中的非常接近,而且渗透压及酸碱度也与血浆的相似,由此证明囊内液确是血浆的超滤液。
单位时间内(每分钟)两肾生成的超滤液量称为肾小球滤过率(glomerularfiltration rate,GFR)。据测定,体表面积为1.73m2的个体,其肾小球滤过率为125ml/min左右。照此计算,两侧肾每一昼夜从肾小球滤出的血浆总量将高达180L。此值约为体重的3倍。肾小球滤过率和肾血浆流量的比例称为滤过分数(filtration fraction)。经测算,肾血浆流量为66ml/min,所以滤过分数为:125/660×100=19%。滤过分数表明,流经肾的血浆约有1/5幔有小球小茁到囊腔中。肾小球滤过率大小决定于滤过系数(Kf)(即滤过膜的面积及其通透性的状态)和有效滤过压。肾小球滤过率=Kf×PUF,PUF表示有效滤过压。
一、滤过膜及其通透性
人体两侧肾全部肾小球毛细血管总面积估计在1.5m2以上,这样大的滤过面积有利于血浆的滤过。在正常情况下,人两肾的全部肾小球滤过面积可以保持稳定。但是在急性肾小球肾炎时,由于肾小球毛细血管管腔变窄或完全阻塞,以致有滤过功能的肾小球数量减少,有效滤过面积也因而减少,导致肾小球滤过率降低,结果出现少尿(每昼夜尿量在100-500ml之间)以致无尿(每昼夜尿量不到100ml)。
表8-1 物质的有效半径和肾小球滤过能力的关系
| 物质 | 分子量 | 有效半径(nm) | 滤过能力 |
| 水 | 18 | 0.10 | 1.0 |
| 钠 | 23 | 0.14 | 1.0 |
| 尿素 | 60 | 0.16 | 1.0 |
| 葡萄糖 | 180 | 0.36 | 1.0 |
| 蔗糖 | 342 | 0.44 | 1.0 |
| 菊粉 | 5500 | 1.48 | 0.98 |
| 肌球蛋白 | 17000 | 1.95 | 0.75 |
| 卵白蛋白 | 43000 | 2.85 | 0.22 |
| 血红蛋白 | 68000 | 3.25 | 0.03 |
| 血浆白蛋白 | 69000 | 3.55 | <0.01 |
滤过能力(filterability)值为1.0表示该物质可自由滤过,0则表示不能滤过
不同物质通过肾小球滤过膜的能力决定于被滤过物质的分子大小及其所带的电荷。表8-1表示被滤过物质的分子量和有效半径对滤过的影响。一般来说,有效半径小于1.8nm的物质,如葡萄糖(分子量180)的有效半径为0.36nm,它可以被完全滤过。有效半径大于3.6nm的大分子物质,如血浆白蛋白(分子量约69000)则几乎完全不能滤过。有效半径介于葡萄糖和白蛋白之间的各种物质,随着有效半径的增加,它们被滤过的量逐渐降低,以上事实提示,滤过膜上存在着大小不同的孔道,小分子物质很容易通过各种大小的孔道,而有效半径较大的物质只能通过较大的孔道,用不同有效半径的中性右旋糖酐分子进行实验,也清楚地说明了被滤过物质的大小对滤过的影响。有效半径小于1.8nm的中性右旋糖酐能自由通过滤过膜,有效半径大于3.6nm的右旋糖酐就完全不能通过。有效半径在1.8-3.6nm的右旋糖酐,其滤过量与有效半径成反比,即随着有效半径增大,滤过量就不断减少(图8-6)。
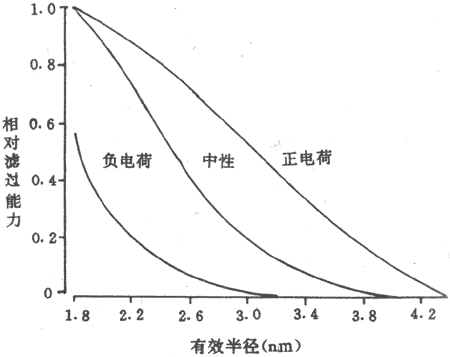
图8-6 不同的有效半径和带不同电荷对右旋糖酐滤过能力的作用
滤过能力的值为1.0,表示自由滤过 0则不能滤过
滤过膜的通性还决定于被滤过物质所带的电荷。用带不同电荷的右旋糖酐进行实验观察到,即使有效半径相同,带正电荷的右旋糖酐较易被滤过,而带负电荷的右旋糖酐则较难通过(图8-6)。血浆白蛋白虽然其有效半径为3.5nm,由于其带负电荷,因此就难于通过滤过膜。
滤过膜的上述特性可由滤过膜的超微结构的特点来说明。滤过膜由三层结构组成(图8-3);①内层是毛细血管的内皮细胞。内皮细胞有上许多直径50-100nm的小孔,称为窗孔(fenestration),它可防止血细胞通过,但对血浆蛋白的滤过可能不起阻留作用。②中间层是非细胞性的基膜,是滤过膜的主要滤过屏障。基膜是由水合凝胶(hydrated gel)构成的微纤维网结构,水和部分溶质可以通过微纤维网的网孔。有人把分离的基膜经特殊染色证明有4-8nm的多角形网孔。微纤维网孔的大小可能决定着分子大小不同的溶质何者可以滤过。③外层是肾小囊的上皮细胞。上皮细胞具有足突,相互交错的足突之间形成裂隙。裂隙上有一层滤过裂隙膜(filtration slit membrane),膜上有直径4-14nm的孔它是滤过的最后一道屏障。通过内、中两层的物质最后将经裂隙膜滤出,裂隙膜在超滤作用中也很重要。
滤过膜各层含有许多带负电荷的物质,主要为糖蛋白。这些带负电荷的物质排斥带带负电荷的血浆蛋白,限制它们的滤过。肾在病理情况下,滤过膜上带负电荷的糖蛋白减少或消失,就会导致带负电荷的血浆蛋白滤过量比正常时明显增加,从而出现蛋白尿。
二、有效滤过压
肾小球滤过作用的动力是有效滤过压。像其它器官组织液生成的机制那样,肾小球有效滤过压=(肾小球毛细血管压+囊内液胶体渗透压)-(血浆胶体渗透压+肾小囊内压)(图8-7)。由于肾小囊内的滤过液中蛋白质浓度较低,其胶体渗透压可忽力略不计。因此,肾小球毛细血管血压是滤出的唯一动力,而血浆胶渗透压和囊内压则是滤出的阻力。有效滤过压=肾小球毛细血管压-(血浆胶体渗透压+肾小囊内压)。皮质肾单位的入球小动脉粗而短,血流阻力较小;出球小动脉细而长,血流阻力较大。因此,肾小球德细血管血压较其它器官的毛细血管血压高。用微穿刺法没得肾小球毛细血管平均值为6.0kPa(45mmHg)(为主动脉平均压的40%左右);用微穿法还发现,由肾小球毛血管的入球端到出球端,血压下降不多,两端的血压几乎相等。肾小囊内压与近曲小管内压力相近。囊内压为1.3kPa(10mmHg)。据测定,在大鼠的肾小球毛细血管入球端的血浆胶体渗透压约为3.3kPa(25mmHg)左右。
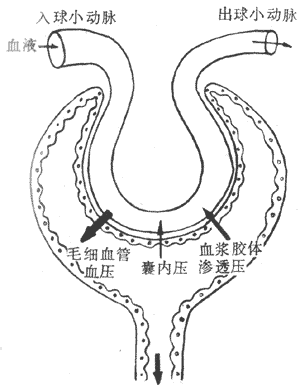
图8-7 有效滤过示意图
在入球端,有效滤过压=6.0-(3.3+1.3)=1.4kPa。但肾小球毛细血管内的血浆胶体渗透压不是固定不变的。在血液流经肾小球毛细血管时,由于不断生成滤过液,血液中血浆蛋白浓度就会逐渐增加,血浆胶体渗透压也随之升高。因此,有效滤过压也逐渐下降。当有效滤过压下降到零时,就达到滤过平衡(filtration equilibrium),滤过便停止了(图8-8)。由此可见,不是肾小球毛细血管全段都有滤过作用,只有从入球小动脉端到滤过平衡这一段才有滤过作用。滤过平衡越靠近入球小动脉端,有效滤过的毛细血管长度就越短,有效滤过压和面积就越小,肾小球滤过率就低。相反,滤过平衡越靠近出球小动脉端,有效滤过的毛细血管长度越长,有效滤过压和滤过面积就越大,肾小球滤过率就越高。如果达不到滤过平衡,全段毛细血管都有滤过作用(图8-8)。
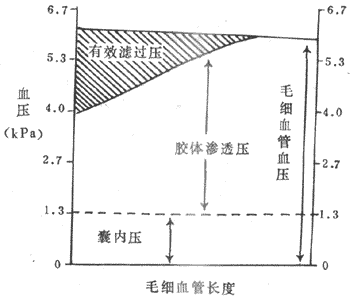
图8-8肾小球毛细血管血压,胶体渗透压和囊内压对肾小球滤过率的作用
三、影响肾小球滤过的因素
滤过膜的通透性和滤过面积的改变对肾小球滤过功能的影响前已述。下面进一步分析肾小球毛细血管血压、血浆胶体渗透压、囊内压和肾血浆流量变化对肾小球滤过功能的影响。
(一)肾小球毛细血管血压
全身动脉血压如有改变,理应影响肾小球毛细血管的血压。由于肾血流量具有自身调节机制,动脉血压变动于10.7?/FONT>24kPa(80-45mmHg)范围内时,肾小球毛细血管血压维持稳定,人而使肾小球滤过率基本保持不变(图8-5)。但当动脉血压降到10.7kPa(80mmHg)以下时,肾小球毛细血管将相应下降,于是有效滤过压降低,肾小球滤过率也减少。当动脉血压降到5.3-6.7kPa(40-50mmHg)以下时,肾小球滤过率将降低到零,因而无尿。在高血压病晚期,入球小动脉由于硬化而缩小,肾小球毛细血管血压可明显降低,于是肾小球滤过率减少而导致少尿。
(二)囊内压
在正常情况下,肾小囊内压是比较稳定的。肾盂或输尿管结石、肿瘤压迫或其他原因引起的输尿管阻塞,都可使肾盂内压显着升高。此时囊内压也将升主,致使有效滤过压降低,肾小球滤过率因此而减少。有些药物如果浓度太高,可在肾小管液的酸性班干部析出结晶;某些疾病时溶血过多,血红蛋白过可堵塞肾小管,这些情况也会导致囊内压升高而影响肾小球滤过。
(三)血浆胶体渗透压
人体血浆胶渗透坟在正常情况下不会有很大变动。但若全身血浆蛋白的浓度明显降低时,血浆胶体渗透压也将降低。此时有效滤过压将升高,肾小球滤过率也随之增加。例如由静脉快速注入生理盐水时,肾小球滤过率将增加,其原因之一可能是血浆胶体渗透压的降低。
(四)肾血浆流量
肾血浆流量对肾小球滤过率有很大影响,主要影响滤过平衡的位置。如果肾轿浆流量加大,肾小球毛细血管内血浆胶体渗透压的上升速度减慢,滤过平衡就靠近出球小动脉端,有效滤过压和滤过面积就增加,肾小球滤过率将随之增加。如果肾血流量进一步增加,血浆胶体渗透压上升速度就进一步减慢,肾小球毛细血管的全长都达不到滤过平衡,全长都有滤过,肾小球滤过率就进一步增加。相反,肾血浆流量减少时,血浆胶体渗透压的上升速度加快,滤过平衡就靠近入球小动脉端,有效滤过压和滤过面积就减少,肾小球滤过率将减少(图8-8)。在严重缺氧、中毒性休克等病理情况下,由于交感神经兴奋,肾血流量和肾血浆流量将显著减少,肾小球滤过率也因而显著减少。
第三节 肾小管与集合管的转运功能
人两肾每天生成的肾小球滤过液达180L,而终尿仅为1.5L。这表明滤过液中约99%的水被肾小管和集合管重吸收,只有约1%被排出体外。不仅如此,滤过液中的葡萄糖已全部被肾小管重吸收回血;钠、尿素告示不同程度地重吸收;肌酐、尿酸和K+等还被肾小管分泌入管腔中。
一、肾小管与集合管的转运方式
肾小管和集合管的转运包括重吸收和分泌。重吸收是指物质从肾小管液中转运至血液中,而分泌是指上皮细胞本身产生的物质或血液中的物质转运至肾小管腔内。肾不球滤过液进入肾小管后称为小管液。
物质通过细胞的转运包括被动转运和主动转运。被动转运是指溶质顺电化学梯度通过肾小管上皮细胞的过程。水的渗透夺之差是水的转运动力。水从渗透压低一侧通过细胞膜进入渗透压高一侧。
主动转运是指溶质逆电化学梯度通过肾小管上皮细胞的过程。主动转运需要消耗能量,根据主动转运过程中能量来源的不同,分为原发性主动转运和继发性主动转运。原发性主动转运(简称为主动转运)所需要消耗的能量由ATP水解直接提供。例如Na+和K+的主动转运是靠细胞膜上的Na+泵水解ATP直接提供能量的。继发性主动转运所需的能量不是直接来自Na+泵。而是来自其他溶质顺电化学梯度转运时释放的。例如一些物质的继发性主动转运的动力直接来自Na+顺电化学梯度转运时释放的能量。释放的之些能量归根到底也是来自Na+泵。由于上皮细胞基侧膜上存在Na+,将细胞内的Na+泵至细胞外,造成细胞内的Na+浓度明显低于细胞外,细胞外K+被泵回细胞内,造成细胞内K+浓度明显高于细胞外,并维持细胞内的负电位。这样,小管液中的Na+便顺电化学梯度通过管腔膜进入细胞,并释放能量提供其他物质的转运。许多物质的转运都与Na+的主动转运相耦联,例如小管液中的葡萄糖、氨基酸、有机酸和CI-等物质的重吸收都与Na+同向转运(cotransport)有关。同向转运是指两种物质与细胞膜上的同向转运体(cotransporter,symporter)特殊蛋白质结合,以相同方向通过细胞膜的转运;又如肾小管细胞分泌H+是与Na+的逆向转运相耦联。逆向转运(antiport)是指两种物质与细胞膜上的逆向转运体(antiport)又称交换体(exchanger)结合,以相反方向通过细胞膜的转运。可见,Na+的主动转运在肾小管上皮细胞的转运中起着关键作用(图8-9)。一个带下电荷和另一个带负电荷的两种物质的同向转动,或电荷相同的两种物质的逆向转运都不会造成小管内外电位改变,这种转运称为电中性转动。如果一个物质是离子,另一个是电中性物质,这种转运就会使小管内外出现电位差,称为生电性转运。如在近球小管,Na+与葡萄糖的同向转运,因葡萄糖是电中性物质,Na+和葡萄糖被重吸收就会造成小管内较小管外带负电位。又如在近球小管的后半段,小管液CI-浓度比管外高,CI-顺浓度差被动重吸收造成管内带正电位。
图8-9 Na+转运与其他溶质转运之间的伴联关系
二、各段肾小管和集合管的转运功能
(一)近球小管
肾小球滤过流经近球小管后,滤过液中67%Na+、CI-、K+和水被重吸收,85%的HCO3也被重吸收,葡萄糖、氨基酸全部被重吸收;H+则分泌到肾小管中。近球小管重吸收的关键动力是基侧膜上的Na+泵;许多溶质,包括水的重吸收都与Na+泵的活动有关。
1.Na+、CI-和水的重吸收在近球小管前半段,大部分Na+与葡萄糖,氨基酸同向转运、与H+逆向转运而被主动重吸收;面近球小管后半段,Na+和CI-主要通过细胞旁路而被被动重吸收。水随NaCI等溶质重吸收而被重吸收,因此,该段小管注与血浆渗透压相同,是等渗重吸收。
在近球小管前半段,由于Na+泵的作用,Na+被泵至细胞间隙,使细胞内Na+浓度低,细胞内带负电位。因此,小管液中的Na+和葡萄糖与管腔膜上的同向转运体结合后,Na+顺电化学梯度通过管腔膜的同时,释放的能量将葡萄糖同向转运入细胞内。进入细胞内的Na+即被细胞基侧膜上的Na+泵泵出至细胞间隙,这样,一方面使细胞内Na+的浓度降低,小管液中的Na+-葡萄糖便可不断转运进入细胞内,细胞内的葡萄糖由易化扩散通过细胞基侧膜离开细胞回到血液中;另一方面,使细胞间隙中的Na+浓度升高,渗透压也升高,通过渗透作用,水随之进入细胞间隙。由于细胞间隙在管腔膜侧的紧密连接相对是密闭的,Na+和水进入后就使其中的静水压升高,这一压力可促使Na+和水通过基膜进入相邻的毛细血管而被重吸收,但也可能使部分Na+和水通过紧密连接回漏(back-leak)至小管腔内(图8-10A)。
另一部分的Na+-H+交换而主动重吸收。小管液中的Na+和细胞内的H+与管腔膜上的交换体结合进行逆向转运,使小管液中的Na顺浓度梯度通过管腔膜进入细胞的同时,将细胞内的H+分泌到小管液中;进入细胞内的Na+随即被基侧膜上的Na+泵泵至细胞间隙而主动重吸收。分泌到小管液中的H+将有利于小管液中的HCO3的重吸收。
图8-10 近球小管重吸收NaCI的示意图
A:近球小管的前半段 X代表葡萄糖、氨基酸、磷酸盐CI-
B:近球小管的后半段F-代表甲酸盐 HF,甲酸
在近球小管后半段,NaCI是通过细胞旁路和跨上皮细胞两条途径而被重吸收的。小管液进入近球小管后半段时,绝大多数的葡萄糖、氨基酸已被重吸收。由于HCO3重吸收速率明显大于CI-重吸收,CI-留在小管液中,造成近球小管后半段的CI-浓度比管周组织间液高20%-40%。因此,CI-顺浓度梯度经细胞旁路(即通过紧密连接进入细胞间隙)而重吸收回血。由于CI-被动重吸收是生电性的,使小管液中正离子相对较多,造成管内外电位差,管腔内带正电,管外带负电,在这种电位差作用下,Na+顺电位差通过细胞旁路而被动重吸收。CI-通过细胞旁路重吸收是顺浓度梯度进行的,而Na+通过细胞旁路重吸收是顺电位梯度进行的,因此,NaCI是重吸收都是被动的(图8-10B)。
NaCI跨上皮细胞重吸收与H+和甲酸盐(formate,F-)再循环有关,要通过管腔膜上相互耦联的Na+-H+交换和CI—甲酸根交换。在正常肾小管液中含有低浓度甲酸根,通过Na+、H+交换,Na+进入细胞,H+分泌到小管液中,并与小管液中的甲酸根结合形成甲酸(formic acid,HF)。甲酸是脂溶性的,可迅速通过管腔膜进入细胞,在细胞内分解为H+和甲酸根。甲酸根和小管液中的CI-进行逆向转运,结果,CI-进入细胞内并通过基侧膜而被重吸收,而甲酸根则人细胞内进入小管液。细胞内的H+则与小管液中的Na+进行逆向交换,Na+进入细胞,并被Na+泵泵至细胞间隙,然后进入管周毛细血管而被重吸收;H+分泌至小管液,再与小管液中的甲酸根据结合,形成甲酸再进入细胞。因此,H+和甲酸根可再循环使用,CI-和Na+则被重吸收回血。
水的重吸收是被动的,是靠渗透作用而进行的。水重吸收的渗透梯度存在于上皮细胞和细胞间隙之间。这是由于Na+顺电化学梯度通过管腔膜进入细胞后,细胞内的Na+被基侧膜上的Na+泵泵至细胞间隙,使细胞间隙渗透压升高。在渗透作用下,水便不断从小管液进入上皮细胞,并从细胞不断进入细胞间隙,造成细胞间隙静水压升高;加上管周毛细血管内静水压较低,胶体渗透压较高,水便通过周围组织间隙进入毛细血管而被重吸收。
2.HCO3重吸收与H+的分泌 HCO3的重吸收与小管上皮细胞管腔膜上的Na+-H+交换有密切关系。HCO3在血浆中钠盐(NaHCO3)的形式存在,滤过中的NaHCO3滤入囊腔进入肾小管后可解离成Na+和HCO3。通过Na+-H+交换,H+由细胞内分泌到小管液中,Na+进入细胞内,并与细胞内的HCO3一起被转运回血(图8-11)。由于小管液中的HCO3不易通过管腔膜,它与分泌的H+结合生成H2CO2,在碳酸酐酶作用下,H2CO2迅速分解为CO2和水。CO2是高度脂溶性物质,能迅速通过管腔膜进入细胞内,在碳酸酐酶作用下,进入细胞内的CO2与H2O结合生成H2CO3。H2CO3又解离成H+和HCO3。H+通过Na+-H+交换从细胞分泌到小管液中,HCO3则与Na+一起转运回血。因此,肾小管重吸收HCO3是以CO2的形式,而不是直接以HCO3的形式进行的。如果滤过的HCO3超过了分泌的H+,HCO3就不能全部(以CO2形式)被重吸收。由于它不易透过管腔膜,所以余下的便随尿排出体外。可见肾小管上皮细胞分泌1H+就可使1HCO3和1Na+重吸收回血,这在体内的酸碱平衡调节中起到重要作用。乙酰唑胺可抑制碳酸酐酶的活性,因此,用乙酰唑受后,Na+-H+交换就会减少,因而NaHCO3、NaCI和水的排出增加,可引起利尿。由于近球小管的Na+-H+交换,小管液中的HCO3与H+结合并生成CO2,CO2透过管腔膜的速度明显高于CI-的速度。因此,HCO3的重吸收率明显大于CI-的重吸收率。
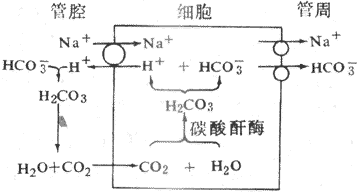
图8-11 肾小管上皮细胞生成和分泌H+示意图
3.K+的重吸收微穿刺实验表明,肾小球滤过的K+,67%左右在近球小管重吸收回血,而尿中的K+主要是由远曲小管和集合管分泌的。有人认为,近球小管对K+的重吸收是一个主动转运过程。小管液中钾浓度为4mmol/L,大大低于细胞内K+浓度(150mmol/L)。因此在管腔膜处K+重吸收是逆浓度梯度进行的。管腔膜K+主动重吸收的机制尚不清楚。
4.葡萄糖重吸收肾小球滤过液中的葡萄糖浓度与血糖浓度相同,但尿中几乎不含葡萄糖,这说明葡萄糖全部被重吸收因血。微穿刺实验表明,重吸收葡萄糖的部位仅限于近球小管,尤其是在近球小管前半段,其他各段肾小管都没有重吸收葡萄糖的能力。因此,如果在近球小管以后的小管液中仍含有葡萄糖,则尿中将出现葡萄糖。
葡萄糖是不带电荷的物质,它的逆浓度梯度重吸收的,是由Na+继发性主动同向转运而被重吸收的。在兔肾近球小管微灌流实验中观察到,如果灌流液中去掉葡萄糖等有机溶质,则Na+的重吸收率降低;如果灌流液中的Na+全部去掉,则葡萄糖有机溶质的重吸收将完全停止,说明葡萄糖的重吸收与Na+同向转运密切相关。葡萄糖和Na+分别与管腔膜上的同向转运体蛋白的结合位点相结合而进行同向转运(见前述Na+重吸收)。
近球小管对葡萄糖的重吸收有一定限度。当血液中葡萄糖浓度超过160-180mg/100ml时,有一部分肾小管对葡萄糖的吸收已达到极限,尿中开始出现葡萄糖,此时的血糖浓度称为肾糖阈。血糖浓度再继续升高,尿中葡萄糖含量也将随之不断增加;当血糖浓度超过300mg/100ml后,全部肾小管对葡萄糖的吸收均已达到极限,此值即为葡萄糖吸收极限量。此时,尿葡萄糖排出率则随血糖浓度升高而平行增加。人肾的葡萄糖吸收极限量,在体表面积为1.73m2的个体,男性为375mg/min,女性为300mg/min。肾之所以有葡萄糖吸收极限量,可能是由于同向转运体的数目有限的缘故,当所有同向转运体的结合位点都被结合而达饱和时,葡萄糖转运量就无从再增加了。
5.其他物质的重吸收和分泌小管注保的氨基酸的重吸收与葡萄糖的重吸收机制相同,也与Na+同向转运(图8-9)。但是,转运葡萄糖的和转运氨基酸的同向转运体可能不同,也就是说同向转运体具有特异性。此外,HPO4-2、SO4-2的重吸收也也Na+同向转运而进行。正常时进入滤液中的微量蛋白质则通过肾小管上皮细胞吞饮作用而被重吸收。
体内代谢产物和进入体仙的某些物质如青霉素、酚红,大部分的利尿药等,由于与血浆中蛋白结合而不能通过肾小球滤过,它们均在近球小管被主动分泌到小管液中而排出体外。
(二)髓袢
近球小管液流经髓袢过程中,约20%的Na+、CI-、和K+等物质被进一步重吸收。髓袢升支粗段的NaCI重吸收在尿液稀释和浓缩机制中具有重要意义。髓袢升支粗段CI-是逆电化学梯度被上皮细胞重吸收的。微穿刺实验证明,兔髓袢升支粗段管腔内为正电位(+10mV)。在微灌流实验中,如果灌流液中不含K+,则管内的正电位基本消失,CI-重吸收率很低,这说明管腔内正电位与CI-的重吸收和小管液中的K+有密切关系。如果在髓袢升支粗段管周的浸溶液中加入选择性Na+泵抑制剂哇巴因(ouabain)抑制Na+泵后,CI-的转运也受阻,说明Na+泵是CI-重吸收的重要因素。据上述实验,有人提出Na+:2CI-:K+同向转运模式来解释升支NaCI的继发性主动重吸收。该模式认为:①髓袢升支粗段上皮细胞基侧膜上的Na+泵,将Na+由细胞内示向组织间液,使细胞内的Na+下降,造成管腔内与细胞内Na+有明显的浓度梯度;②Na+与管腔膜上同向转运体结合,形成Na+:2CI-:K+同向转运体复合物,Na+顺电化学梯度将2CI-和K+一起同向转运至细胞;③进入细胞内的Na+、2CI-和K+的去向各不相同:Na+由Na+泵泵至组织间液,2CI-由于浓度梯度经管周膜上CI-通道进入组织间液,而K+则顺浓度梯度经管腔膜而返回管腔内,再与同向转运体结合,继续参与Na+:2CI-K+的同向转运,循环使用;④由于2CI-进入组织间液,K+返回管腔内,这就导致管腔内出现正电位;⑤由于管腔内正电位,使管腔液中的Na+等正离子顺电位差从细胞旁路进入组织间液,这是不耗能的Na+被动重吸收。从这个模式说明,通过Na+泵的活动,继发性主动重吸收了2CI-,同时伴有2Na+的重吸收,其中1Na+是主动重吸收,另1Na+通过细胞旁路而被动重吸收,这样为Na+的重吸收节约了50%能量消耗(图8-12)。髓袢升支粗段对水的通透性很低,水不被重吸收而留在小管内。由于NaCI被上皮细胞重吸收至组织间液,因此造成小管液低渗,组织间液高渗。这种水和盐重吸收的分离,有利于尿液的浓缩和稀释。Na+:2CI-:K+同向转运对速尿,利尿酸等利尿剂很敏感。它们与同向转运体结合后,可抑制其转运功能,管腔内正电位消失,NaCI的重吸收受抑制,从而干扰尿的浓缩机制,导致利尿。
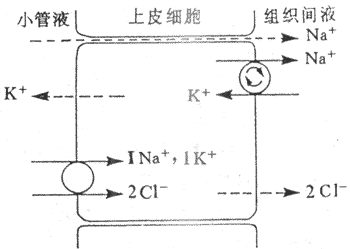
图8-12 髓袢升支粗段继发性主动吸收CI-的示意图
(三)远球小管和集合管
在远曲小管和集合管,重吸收大约12%滤过的Na+和CI-,分泌不同量的K+和H+,重吸收不同量的水。水、NaCI的重吸收以及K+和H+的分泌可根据机体贴的水、盐平衡状况来进行调节。如相机缺水或缺盐时,远曲小管和集合管可增加水、盐的重吸收;当机体水、盐过剩时,则水、盐重吸收明显减少,水和盐从尿排出增加。因此,远曲小管和集合管对水和盐的转运是可被调节的。水的重吸收主要受抗利尿激素调节,而Na+和K+的转运主要受醛固酮调节。
远曲小管和集合管上皮细胞间隙的紧密连接对小离子如Na+、K+和CI-等的通透性低,这些离子不易通过紧密连接回漏至小管腔内,因此,所能建立起来的管内外离子浓度梯度和电位梯度大。在远曲小管初段,对水的通透性很低,但仍主动重吸收NaCI,继续产生低渗小管液。Na+在远曲小和集合管的重吸收是逆较大的电化学梯度进行的,是主动重吸收过程。这可能与远曲小管的Na+泵在肾单位中活性最高有关。有人认为在远曲小管初段的小管液中,Na+是通过Na+-CI-同向转运进入细胞的,然后由Na+泵将Na+泵出细胞而主动重吸收回血(图8-13A)。NaCI同向转运体可被噻嗪类利尿药所抑制。
远曲小管后段和集合管含有两类细胞,即主细胞和闰细胞。主细胞重吸收Na+和水,分泌K+,闰细胞则主要分泌H+,主细胞重吸收Na+主要通过管腔膜上的Na+通道。管腔内的Na+顺电化学梯度通过管膜上的Na+通道进入细胞,然后,由Na+泵泵至细胞间液而被重吸收(图8-13B)。
K+的分泌 尿中K+的排泄量视K+的摄入量而定,高钾饮食可排出大量的钾,低钾饮食则尿中排钾量少,使机体的钾摄入量与排出量保持平衡,维持机体K+浓度的相对恒定。
K+分泌的动力包括:①在远曲小管和集合管的小管液中,Na+通过主细胞的管腔膜上的Na+通道进入细胞,然后,由基侧膜上的Na+泵将细胞内的Na+泵至细胞间隙而被重吸收,因而是生电性的,使管腔内带负电位(-10?40mV)。这种电位梯度也成为K+从细胞分泌至管腔的动力;②在远曲小管后段和集合管的主细胞内的K+浓度明显高于小管液中的K+浓度,K+便顺浓度梯度从细胞内通过管腔膜上的K+通道进入小管液;③Na+进入主细胞后,可刺激基侧膜上的Na+,使更多的K+从细胞外液中泵入细胞内,提高细胞内K+浓度,增加细胞内和小管液之间的K+浓度梯度,从而促进K+分泌,因此,K+的分泌与Na+的重吸收有密切关系。(图8-13B)。
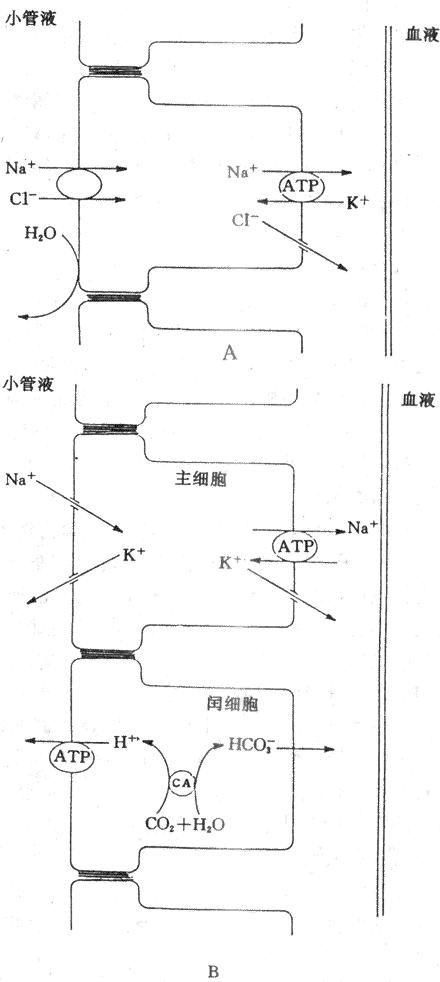
图8-13 远球小管和集合管重吸收NaCI、分泌K+和H+的示意图
A:远曲小管初段 B:远曲小管后段和集合管
H+的分泌除了近球小管细胞通过Na+-H+交换分泌H+,促进NaHCO3重吸收外,远曲小管和集合管的闰细胞也可分泌H+。H+的分泌是一个逆电化学梯度进行的主动转运过程。有人认为管腔膜上有H+泵,能将细胞内的H+历史意义入小管腔内。细胞内的CO2和H2O在碳酸酐酶催化作用下生成的H+和HCO3,H+由H+泵泵至小管液,HCO3则通过基侧膜回到血液中,因而H+分泌和HCO3的重吸收与酸碱平衡的调节有关(图8-13B)。闰细胞分泌的H+与HPO4-2结合形成H2PO4,这是可滴定酸;分泌的H+可与上皮细胞分泌的NH3结合,形成NH4+。可滴定酸和NH4+都不易透过管腔膜进入细胞而留在小管液中。因此,它们是尿液酸碱度的决定因素。
NH3的分泌远曲小管和集合管的上皮细胞在代谢过程中不断地生成NH3,这些NH3主要由谷氨酰胺脱氨而来。NH3具有脂溶性,能通过细胞膜向小管液周围组织间液和小管液自由扩散,扩散量取决于两种液体的pH值。小管液的pH较低(H+浓度较高),所以NH3较易向小管液中扩散。分泌的NH3能与小管液中的H+结合并生成NH4-,小管液中NH3浓度因而下降,于是管腔膜两侧形成了NH3浓度梯度,此浓度梯度又加速了NH3向小管液中扩散。由此可见,NH3的分泌与H+的分泌密切相关;H+分泌增加促使NH3分泌增多。NH3与H+结合并生成NH4-后,可进一步与小管液中的强酸盐(如NaCI等)的负离子结合,生成酸性铵盐(NH4CI等)并随尿排出。强酸盐的正离子(如Na+)则与H+交换而进入肾小管细胞,然后和细胞内HCO3-一起被转运回血。所以,肾小管细胞分泌NH3,不仅由于铵盐形成而促进了排H+,而且也促进了NaHCO3的重吸收。
第四节 尿液的浓缩和稀释
尿的渗透浓度可由于体内缺水或水过剩等不同情况而出现大幅度的变动。当体内缺水时,机体将排出渗透浓度明显高于血浆渗透浓度的主渗尿,即尿被浓缩。而体内水过剩时,将排出渗透浓度低于血浆渗透浓度的低渗尿。正常人尿液的渗透浓度可在约50-1200mOsm/kgH2O之间波动。所以,根据尿的渗透浓度可以了解肾的渗透浓度和稀释能力。肾的浓缩和稀释能力,以维持体液平衡和渗透压恒定中有极为重要的作用。
一、尿液的稀释
尿液的稀释是由于小管液的溶质被重吸收而水不易被重吸收造成的。这种情况主要发生在髓袢升支粗段。前已述及,髓袢升支粗段能主动重吸收Na+和CI-(图8-12),而对水通透,故水不被重吸收,造成髓袢升支粗段小管液为低渗。在体内水过剩而抗利尿激素释放被抑制时,集合管对水的通透性非常低。因此,髓袢升支的小管液流经远曲小管和集合管时,NaCI继续重吸收,使小管液渗透浓度进一步下降。可降低至50mOsm/kgH2O,形成低渗尿,造成尿液的稀释。如果抗利尿激素完全缺乏时,如严重尿崩症患者,每天可排出高达20L的低渗尿,相当于肾小球滤过率的10%。
二、尿液的浓缩
尿液的浓缩是由于小管液中的水被重吸收而溶质仍留在小管液中造成的。水重吸收的动力来自肾髓质渗透梯度的建立,即髓质渗透浓度从髓质外层向乳头部深入而不断升高。用冰点降低法测定鼠肾的渗透浓度观察到肾皮质部的组织间液(包括细胞内液和细胞外液)的渗透浓度与血液渗透浓度之比为1.0,说明皮质部组织间液与血浆是等渗的。而髓质部组织间液与血浆的渗透浓度之比,随着由髓质外层向乳头部深入而逐渐升主,分别为2.0、3.0、4.0(图8-14)。这表明肾髓质的渗透浓度由外向内逐步升高,具有明确的渗透梯度。在抗利尿激素存在时,远曲小管和集合管对水通透性增加,小管液从外髓集合管向内髓集合管流动时,由于渗透作用,水便不断进入高渗的组织间液,使小管液不断被浓缩而变成高渗液,最后尿液的渗透浓度可高达120mOsm/kgH2O,形成浓缩尿。可见髓质的渗透梯度是建立就成为浓缩尿的必要条件。髓袢是形成髓质渗透梯度的重要结构,只有具有髓袢的肾才能形成浓缩尿,髓袢愈长,浓缩能力就愈强。例如沙鼠的肾髓质内层特别厚,它的肾能产生20倍于血浆渗透浓度的高渗尿。猪的髓袢较短,只能产生1.5倍于血浆渗透浓度的尿液。人的髓袢具有中等长度,最多能产生4-5倍于血浆渗透浓度的高渗尿。
髓质渗透梯度是如何形成的?有人用肾小管各段对水和溶质通透性不同(表8-2)和逆流倍增现象来解释。
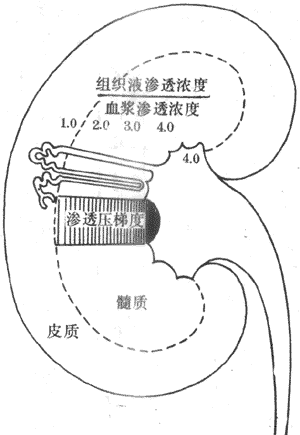
8-14图 肾髓质渗透压梯度示意图
线条越密,表示渗透压越高
表8-2 兔肾小管不同部分的通透性
| 肾小管部分 | 水 | Na+ | 尿素 |
| 髓袢升支粗段 | 不易通透 | Na+主动重吸收 | 不易通透 |
| CI-继发性主动重吸收 | |||
| 髓袢升支细段 | 不易通透 | 易通透 | 中等通透 |
| 髓袢降支细段 | 易通透 | 不易通透 | 不易通透 |
| 远曲小管 | 有ADH时水易通透 | Na+主动重吸收 | 不易通透 |
| 集合管 | 有ADH时水易通透 | Na+主动重吸收 | 皮质稳步髓部不易通透 |
ADH为抗利尿激素*不同动物,通透性不一样
物理学中逆流的涵意是指两个并列的管道,其中液体流动的方向相反,如图8-15所示,甲管中液体向下流,乙管中液体向上流。如果甲乙两管下端是连通的,而且两管间的隔膜容许液体中的溶质或热能在两管间交换,便构成了逆流系统。在逆流系统中,由于管壁通透性和管道周围环境的作用,就会产生逆流倍增现象。
图8-15 逆流系统示意图
甲管内液体向下流 乙管内液体向上流
逆流倍增现象可要根据图8-16的模型来理解。模型中含有钠盐的液体从甲管流进,通过管下端的弯曲部分又折返流入乙管,然后从乙管反向流出,构成逆流系统。溶液流动时,由于M1膜能主动将Na+由乙管泵入甲管,而M1膜对水的通透性又很低,因此,甲管中深夜在向下流动过程中将不断接受由乙管泵入的Na+,于是Na+的浓度不断增加(倍增)。结果甲管中溶液自上而下的渗透浓度会越来越高,到甲管下端的弯曲部分时Na+深夜逐渐下降,渗透浓度也相应下降。这样,不论是甲管还是乙管,从上而下来比较,溶液的渗透浓度均逐渐升高,即出现了逆流倍增现象,形成了渗透梯度。如果有渗透浓度较低的溶液从丙管向下流动,而且M2膜对水不能通透,对溶质不通透,水将因渗透作用而进入乙管。这样丙管内溶质的深夜将逐渐增加;从丙管下端流出的液体成了高渗溶液。

图8-16 逆流倍增作用模型
甲管内液体向下流 乙田径赛内液体向上流丙管内液体向下流 M1膜能将液体
中Na+由乙管泵入甲管,且对水不易通透M2膜对水易通透
髓袢、集合管的结构排列与上述的逆流倍增的模型很相似。这对理解尿的浓缩机制是有帮助的。
在外髓部,由于髓袢升支粗段能主动重吸收Na+和CI-(图8-17),而对水不通透,故升支粗段内小管液向皮质方向流动时,管内NaCI浓度逐渐降低,小管液渗透浓度逐渐下降;而升支粗段外围组织产液则变成高渗。髓袢升支粗段位于外髓部,故外髓部的渗透梯度主要是由升支粗段NaCI的重吸收所形成。愈靠近皮质部,渗透浓度越低;愈靠近内髓部,渗透浓度越高。
在内髓部,渗透梯度的形成与尿素的丙循环和NaCI重吸收有密切关系。①远曲小管及皮质部和外髓部的集合管对尿素不易通透,但小管液流经远曲小管及皮质部和外髓部的集合管时,在抗利尿激素作用下,对水通透性增加,由于外髓部高渗,水被重吸收,所以小管液中尿素的浓度逐渐升高。②当小管液进入内髓部集合管时,由于管壁对尿素的通透性增大,小管液中尿素就顺浓度梯度通过管壁向内髓部组织间液扩散,造成了内髓部组织间液中尿素浓度的增高,渗透浓度因之而长高。③髓袢降支细段对尿素不易通透,而对水则易通透,所以在渗透压的作用下,水被“抽吸”出来,从降支细段进入内髓部组织间液。由于降支细段对Na+不易通透,小管液将被浓缩,于是其中的NaCI浓度愈来愈高,渗透浓度不断升高。④当小管液绕过髓袢顶端折返流入升支细段时,它同组织间注NaCI浓度梯度明显地建立起来。由于升支细段对Na+易通透,Na+将顺浓度梯度而被动扩散至内髓部组织间液,从而进一步提高了内髓部组织间液的渗透浓度。由此看来,内髓部组织间液的渗透浓度,是由内髓部集合管扩散出来的尿素以及髓袢升支细段扩散出来的NaCI两个因素造成的。⑤小管液在升支细段流动过程中,由于NaCI扩散到组织间液,而且该管壁又对水不易通透,所以造成了管内NaCI浓度逐渐降低,渗透浓度也逐渐降低,这样,降支细段与升支细段就构成了一个逆流倍增系统,使内髓部组织间液形成了渗透梯度。⑥尿素是可以再循环的。因为升支细胞对尿素具有中等的通透性,所以从内髓部集合管扩散到组织间液的尿素可以进入升支细段,而后流过升支粗段、远曲小管、皮质部和外髓部集合管,又回到内髓部集合管外再扩散到内髓部组织间液,这样就形成了尿素的丙循环(图8-17)。
图8-17 尿浓缩机制示意图
粗箭头表示升支粗段主动重吸收Na+和CI-。粗线表示髓袢升支粗段和远曲小管前段对水不通透。字体大小表示溶质深夜。XS表示未被重吸收的溶质
从髓质渗透梯度形成全过程来看,髓袢升支粗段对Na+和CI-的主动重吸收是髓质渗透梯度建立的主要动力。而尿素和NaCI是建立髓质渗透梯度的主要溶质。
三、直小血管在保持肾髓质高渗中的作用
直小血管的功能可用逆流交换现象来理解。图8-18是逆流交换的示意图。在图A中,U形管的升、降支之间不能进行热量交换,所以降支中的冷水在流到热源以前得不到加温,升支中的水温在离开热源以后也不能降低。这样,冷水流过U形管的升、降支之间能够交换热量,所以降支中的冷水在进入热源以前就被升支管壁透过来的热量所加温,而升支中的水则因热量不断透入降支而降温。这样,冷水流过U形管时,从热源带走的热量就很有限,所在热源损失掉的热量也很少。
图8-18 逆流交换作用的简单物理模型示意图
下如前述,通过肾小管上述的逆流作用,不断有溶质(NaCI和尿素)进入髓制裁组织间液形成渗透梯度,也不断有水被肾小管和集合管重吸收至组织间液。因此,必须把组织间液中多余的溶质和水被除去才能保持髓质渗透梯度。通过直小血管的逆流交换作用就能保持髓质渗透梯度。直小血管的降支和长支是并行的细血管,这咱结构就是逆流系统。在直小血管降支进入髓质的入口处,其血浆渗透浓度约为300mOsm/kgH2O。由于直小血管对溶质和水的通透性高,当它在向髓质深部下行过程中,周围组织间液中的溶质就会顺浓度梯度不断扩散到直小血管降支中,而其中的水则渗出到组织间液,使血管中的血浆渗透浓度与组织间液达到平衡。因此,愈向内髓部深入,降支血管中的溶质浓度愈高。在折返处,其渗透浓度可高达120mOsmkgH2O。如果直小血管降支此时离开髓质,就会把从进入直小血管降支中的大量溶质流回循环系统,而从直小血管内出来的水保留在组织间液。这样,髓质渗透梯度就不能维持。由于直小血管是逆流系统。因此,当直小血管升支从髓质深部返回外髓部时,血管内的溶质浓度比同一水平组织间液的高,溶质又逐渐扩散回组织间液,并且可以再进入降支,这是一个逆流交换过程。因此当直小血管升支离开外髓部时,只把多余的溶质带回循环中。此外,通过渗透作用,组织间液中的水不断进入直小血管升支,又把组织间液中多余的水随血流返回循环。这样就维持了肾髓质的渗透梯度(图8-17)。
第五节 肾尿生成的调节
尿的生成有赖于肾小球的滤过作用和肾小管、集合管的重吸收和分泌作用。因此,机体对尿的生成的调节也就是通过对滤过作用和重吸收、分泌作用的调节来实现的。肾小球滤过作用的调节在前文已述,本节主要论述肾小管和集合管重吸收和分泌的调节。肾小管和集合管功能的调节包括肾内自身调节和神经、体液调节。
一、肾内自身调节
肾内自身调节包括小管液中溶质浓度的影响、球-管平衡和管-球反馈等。
(一)小管液中溶质的浓度
小管液中溶质所呈现的渗透压,是对抗肾小管重吸收水分的力量。如果小管液溶质浓度很高,渗透压很大,就会妨碍肾小管特别是近球小管对水的重吸收,小管液中的Na+被稀释而浓度下降,小管液中与细胞内的Na+浓度差变小,Na+重吸收减少,因此,不仅尿量增多,NaCI排出也增多。例如糖尿病患者的多尿,就是由于小管液中葡萄糖含量增多,肾小管不能将葡萄糖完全重吸收回血,小管液渗透压因而增高,结果妨碍了水和NaCI的重吸收所造成的。临床上有时给病人使用肾小球滤过而又不被肾小管重吸收的物质,如甘露醇等,利用它来提高小管液中溶质的浓度,借以达到利尿和消除水肿的目的。这种利尿方式称为渗透性利尿。
(二)球-管平衡
近球小管对溶质和水的重吸收量不是固定不变的,而是随肾小于滤过率的变动而发生变化。肾小球滤过率增大,滤液中的Na+和水的总含量增加。近球小管对Na+和水的重吸收率也提高;反之,肾小球滤过率减小,滤液中的Na+和水的总含量也减少,近球小管的Na+的水的重吸收率也相应地降低。实验说明,不论肾小于滤过率或增或减,近球小管是定比重吸收(constant fraction reabsorption)的,即近球小管的重吸收率始终占肾小球滤过率的65%-70%左右(即重吸收百分率为65%-70%)。这种现象称为于-管平衡(glomerulotubular balance)。球管平衡的生理意义在于使尿中排出的溶质和水不致因肾小管滤过率的增减而出现大幅度的变动。例如,在正常情况下,肾小球滤过率为125ml/min,近球小管的重吸收率为87.5ml/min(占70%)。流到肾小管远侧部分的量为37.5ml/min。如果滤过率增加到150ml/min,则近球小管的重吸收率变为105ml/min(仍占70%),而流到肾小管远侧部分的量为45ml/min。这几个数字表明,此时滤过率虽然增加了25ml/min,但流到肾小管远见侧部分的量仅增加7.5ml/min。而且在这种情况下,远侧部分的重吸收也有增加,因此尿量的变化是不大的。同样,滤过率减少到100ml/min,近球小管的重吸收率为70ml/min(仍占70%),流到肾小管远侧部分的量为30ml/min。此时的滤过率虽然减少了25ml/min,但流到肾小管远侧部分的量仅减少了7.5ml/min;而且在这种情况下远侧部分的重吸收也要减少,因此尿量的变化仍然不大。近球小管对Na+也是定比重吸收,即重吸收量为滤过量的65%-70%。如果近球小管对Na+重吸收的总量是固定不变的话,根据测算,肾小球滤过率仅增加2ml/min,Na+的排出量就会比原来的增加约2倍;肾小球滤过率减少2ml/min,尿中就不含Na+,可见球管平衡具有重要的生理意义。
定比重吸收的机制与管周毛细血管血压和胶体渗透压改变有关。比如,在肾血流量不变的前提下,当肾小球滤过率增加时,进入近球小管旁毛细血管的血液量就会减少,,血浆蛋白的浓度相对地增高,此时毛细血管内血压下降而胶体渗透压升高。在这种情况下,小管旁组织间液就加速进入毛细血管,组织间液内静水压因之下降,组织间液内静水压下降使得小管细胞间隙内的Na+和水加速通过基膜而进入小管旁的组织间隙;并且通过紧密连接回流至肾小管腔内的回漏量因此而减少,最后导致Na+和水重吸收量增加。这样,重吸收仍可达到肾小球滤过率的65%-70%。肾小球滤过率如果减少,便发生相反的变化,重吸收百分率仍能保持65%-70%。
球-管平衡在某些情况下可能被打乱。例如,渗透性利尿时,近球小管重吸收率减少,而肾小球滤过率不受影响,这时重吸收百分率就会小于65%-70%,尿量和尿中的NaCI排出量明显增多。
目前认为球-管平衡障碍与临床上见到的某些水肿的形成机制有关。例如在充血性心力衰竭时,肾灌注压和血流量可明显下降。但由于出球小动脉发生代偿性收缩,所以肾小球滤过率仍能保持水平。因此滤过分数将变大。此时近球小管旁毛细血管血压下降而血浆胶体渗透压增高。如上所述,这将导致Na+和水的重吸收增加,重吸收百分率将超过65%-70%。于是因体体内钠盐潴留和细胞外液量增多而发生水肿。
(三)管-球反馈
管-球反馈是肾血流量和肾小球滤过率自身调节的重要机制之一。当肾血流量和肾小球滤过率增加时,到达远曲小管致密斑的小管液的流量增加,致密班发生信息,使肾血流量和肾小球滤过率恢复至正常。相反,肾血流量和肾小球滤过率减少时,流经致密斑的小管液流量就下降,致密斑发生信息,使肾血流量和肾小球滤过率增加至正常水平。这种小管液流量变化影响肾血流量和肾小球滤过率的现象称为管-球反馈(tubuloglomerular feed back)。有人认为致密斑主要感受小管液中的NaCI含量改变而不是小管液的流量。一般来说,肾小管液流量与NaCI含量成正比。致密斑发在管-球环节中起重要的传感器(sensor)作用。致密斑与入球小动脉和出球小动脉相邻。致密斑发出的信息通过某种途径影响入球小动脉的口径,从而影响肾血流量和肾小球滤过率。当肾血流量增加时,肾小球滤过率也增加,流经远曲小管的小管液量也增加,致密斑部位NaCI含量升高,致密斑发出信息刺激颗粒细胞释放肾素,导致局部生成血管紧张素Ⅱ,血管紧张素Ⅱ引起入球小动脉收缩,口径缩小,阻力增加,从而使肾血流量和肾小球滤过率恢复至原来水平。相反,当肾血流量减少时,肾小球滤过率下降,流经远曲小管的小管液流量减少,颗粒细胞释放肾素减少,血管紧张至少Ⅱ生成减少,入球小动脉收缩变弱,口径变粗,阻力减少,肾血流量恢复至原有水平。此外,肾内产生的前列腺素、腺苷和儿茶酚胺等也参与管-球反馈。
二、神经和体液调节
(一)交感神经系统
肾交感神经兴奋通过下列作用影响尿生成:①入球小动脉和出球小动脉收缩,而前者血管收缩比后者更明显,因此,肾小球毛细血管的血浆流量减少和肾小球毛细血管的血压下降,肾小球的有效滤过压下降,肾小球滤过率减少;②刺激近球小体中的颗粒细胞释放肾素,导致循环中的血管紧张素Ⅱ和醛固酮含量增加,增加肾小管对NaCI和水的重吸收;③增加近球小管和髓袢皮皮细胞重吸收Na+、CI-和水。微穿刺表明,低频率低强度电刺激肾交感神经,在不改变肾小球滤过率的情况下,可增加近球小管和髓袢对Na+、CI-和水的重吸收。这种作用可被α1肾上腺素受体拮抗剂所阻断。这些结果表明,肾交感神经兴奋时其末稍释放去甲肾上腺素。作用于近球小管和髓袢细胞膜上的α1肾上腺素能受体,增加Na+、CI-和水的重吸收。抑制肾交感神经活动则有相反的作用。
(二)抗利尿激素
抗利尿激素(antidiuretic hormone,ADH)又称血管升压素(vasopressin,AVP),是由9个氨基酸残基组成的小肽,它是下丘脑的视上核和室旁核的神经元分泌的一种激素。它在细胞体中合成,经下丘脑-垂体束被运输到神经垂体然后释放出来。它的作用主要是提高远曲小管和集合管上皮细胞对水的通透性,从而增加水的重吸收,使尿液浓缩,尿量减少(抗利尿)。此外,抗利尿激素也能增加髓袢升支粗段对NaCI的主动重吸收和内髓部集合管对尿素的通透性,从而增加髓质组织间液的溶质浓度,提高髓质组织间液的渗透浓度,有利于尿注浓缩(见尿液浓缩和稀释)。
抗利尿激素与远曲小管和集合管上皮细胞管周膜上的V2受体结合后,激活膜内的腺甘酸化酶,使上皮细胞中cAMP的生成增加;cAMP生成增加激活上皮细胞中的蛋白激酶,蛋白激酶的激活,使位于管腔膜附近的含有水通道的小泡镶嵌在管腔膜上,增加管腔膜上的水通道,从而增加水的通透性。当抗利尿激素缺乏时,管腔膜上的水通道可在细胞膜的衣被凹陷处集中,后者形成吞饮小泡进入胞浆,称为内移(internalization)。因此,管腔膜上的水通道消失,对水就不通透。这咱含水通道的小泡镶嵌在管腔膜或从管腔膜进入细胞内,就可调节管腔内膜对水的通透性(图8-19)。基侧膜则对水可自由通过,因此,水通过管腔膜进入细胞后自由通过基侧膜进入毛细血管而被重吸收。
图8-19 抗利尿激素的作用机制示意图
调节抗利尿激素的主要因素是血浆晶体渗透压和循环血量、动脉血压。血浆晶体渗透压的改变可明显影响抗利尿激素的分泌。大量发汗。严重呕吐或腹泻等情况使机体失水时,血浆晶体渗透压升高,可引起抗利尿激素分泌增多,使肾对水的重吸收活动明显增强,导致尿液浓缩和尿量减少。相反,大量饮清水后,尿液被稀释,尿量增加,从而使机体内多余的水排出体外。例如,正常人一次饮用100ml清水后,约过半小时,尿量就开始增加,到第一小时末,尿量可达最高值;随后尿量减少,2-3小时后尿量恢复到原来水平。如果饮用的是等渗盐水(0.9NaCI溶液),则排尿量不出现饮清水后那样的变化(图8-20)。这种大量饮用清水后引起尿量增多的现象,称为水利尿,它是临床上用来检测肾稀释能力的一种常用的试验。循环血量的改变,能反射性地影响抗利尿激素的释放。血量过多时,左心房被扩张,刺激了容量感受器,传入冲动经迷走神经传入中枢,抑制了下丘脑-垂体后叶系统释放抗利尿激素,从而引起利尿,由于排出了过剩的水分,正常血量因而得到恢复。血量减少时,发生相反的变化。动脉血压升高,刺激颈动脉窦压力感受器,可反射性地抑制抗利尿激素的释放。此外,心房尿钠肽可抑制抗利尿激素分泌,血管紧张素Ⅱ则可刺激其分泌。
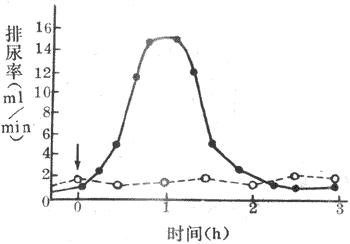
图8-20一次饮一升清水(实线)和饮一升等渗盐水
(0.9NaCI溶液)(虚线)后的利尿率箭头表示饮水时间
(三)肾素-血管紧张素-醛固酮系统
肾素主要是由近球小体中的颗粒细胞分泌的。它是一种蛋白水解酶,能催化血浆中的血管紧张素原使之生成血管紧张素壹I(十肽)。血液和组织中,特别是肺组织中有血管紧张素转换酶,转换酶可使血管紧张素I降解,生成血管紧张素Ⅱ(八肽)。血管紧张素Ⅱ可刺激肾上腺皮质球状带合成和分泌醛固酮。
肾素的分泌受多方面因素的调节。目前认为,肾内有两种感受器与肾素分泌的调节有关。一是入球小动脉处的牵张感受器,另一是致密斑感受器。当动脉血压下降,循环轿量减少时,肾内入球小动脉的压力也下降,血流量减少,于是对小动脉壁的牵张刺激减弱,这便激活了牵张感受器,肾素释放量因此而增加;同时,由于入球小动脉的压力降低和血流量减少,于是激活了致密斑感受器,肾素释放量也可增加。据推想,在近球小体的颗粒细胞和致密斑之间有一种特殊的联系。当两者接触增加时,肾素分泌便减少,而两者接触减少时,则肾素分泌增加。入球小动脉的压力下降,血流量减少时,血管口径缩小,于是颗粒细胞和致密班的接触减少,此时肾素分泌增加;当致密斑处Na+量和小管液量减少时,肾小管口径缩小,两者的接触减少,肾素分泌增加。但这种推想尚缺乏实验证据。此外,颗粒细胞受交感神经支配,肾交感神经兴奋时(如循环血量减少)能引致肾素的释放量增加。肾上腺素和去甲肾上腺素也可直接刺激颗粒细胞,促使肾素释放增加。
1.血管紧张素Ⅱ对尿生成的调节包括:①刺激醛固酮的合成和分泌;醛固酮可调节远曲小管和集合管上皮细胞的Na+和K+转运;②可直接刺激近球小管对NaCI的重吸收,使尿中排出的NaCI减少;③刺激垂体后叶释放抗利尿激素,因而增加远曲小管和集合管对水的重吸收,使尿量减少。
2. 醛固酮对尿生成的调节醛固酮是肾上腺皮质球状带分泌的一种激素。它对肾的作用是促进远曲小管和集合管的主细胞重吸收Na+,同时促进K+的排出,所以醛固酮有保Na+排K+作用。
醛固酮进入远曲小管和集合管的上皮细胞后,与胞浆受体结合,形成激素-受体复合物;后者通过核膜,与核中的DNA特异性结合位点相互作用,调节特异性mRNA转录,最后合成多种的醛固酮诱导蛋白(aldosterone-induced protein)。醛固酮诱导蛋白可能是:①管腔膜的Na+通道蛋白,从而增加管腔的Na+通道数量;②线粒体中合成的ATP的酶,增加ATP的生成,为上皮细胞活动(Na+泵)提供更多的能量;③基侧膜的Na+泵,增加Na+泵的活性,促进细胞内的Na+泵回血液和K+进入细胞,提高细胞内的K+浓度,有利于K+分泌(图8-21);由于Na+重吸收增加,造成了小管腔内的负电位,有利于K+的分泌和CI-的重吸收。结果,在醛固酮的作用下,远曲小管和集合管对Na+和集合管对Na+的重吸收增强的同时,CI-和水的重吸收增加,导致细胞外液量增多;K+的分泌量增加。
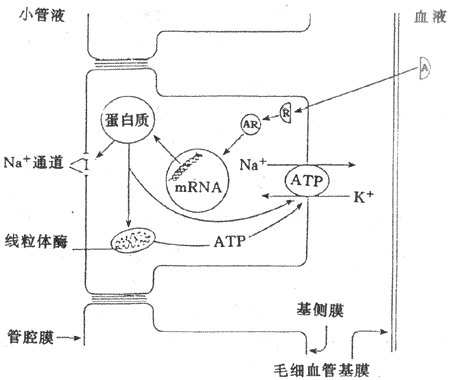
图8-21 醛固酮作用机制的示意图
醛固酮的分泌除了受血管紧张素调节外,血K+浓度升高和血Na+浓度降低,可直接刺激肾上腺皮质球状带增加醛固酮的分泌,导致保Na+排K+,从而维持了血K+和血Na+浓度的平衡;反之,血K+浓度降低,或血Na+浓度升高,则醛固酮分泌减少。醛固酮的分泌对血K+浓度升高十分敏感,血K+仅增加0.5-1.0mmol/L就能引起醛固酮分泌。而血Na+浓度必须降低很多才能引起同样的反应。
(四)心房利尿钠肽
心房利尿钠肽(atrial natriuretic pepitde,ANP)是心房肌合成的激素。循环中的心房利尿钠肽是由28个氨基酸残基组成的。它有明显的促进NaCI和水的排出作用。其作用机制可能包括:①抑制集合管对NaCI的重吸收。心房利尿钠肽与集合管上皮细胞基侧膜上的心房利尿钠肽受体结合,激活了鸟苷酸化酶,造成细胞内cGMP含量增加,后者使管腔膜上的Na+通道关闭,抑制Na+重吸收,增加NaCI的排出;②使出球小动脉、尤其是入球小动脉舒张,增加肾血浆流量和肾小球滤过率③抑制肾素的分泌;④抑制醛固酮的分泌;⑤抑制抗利尿激素的分泌。
第六节 清除率
一、清除率的概念和计算方法
清除率是一个抽象的概念,它把一肾在一定时间内排出的物质的量,同当时该物质在血浆中浓度联系起来,因而能更她地说明肾排出某物质的能力。为了讲清楚它的涵意,现举例说明。某甲每分钟尿量为1ml(V=1ml/min),尿中某物质的浓度(U)为100mg/100ml,血浆中该物质的浓度(P)为1mg/100ml。某乙每分钟尿量为0.8ml(V=0.8ml/min),尿中该物质的浓度(U)为50mg/100ml,血浆中该物质的浓度(P)为0.32mg/100ml。经计算,甲肾的该物质排泄量应为U×V=0.8ml/min×50mg/100ml=0.4mg/min。从每分钟该物质排泄量的多少来看,甲肾的功能似乎比乙肾好,其实则不然。因为甲血浆中该物质的浓度比乙肾血浆中的高三倍多,甲肾清除血浆中该物质比较容易,而乙肾则比较困难。所以,从清除血浆中该物质的能力业看,还应该将血浆中浓度(P)这一因素考虑过去。这样,甲肾将该物质从血浆中清除掉的能力为:
U×V/P=[(100mg/100ml)×1ml/min ]/(1mg/100ml)=100ml/min
乙肾为:U×V/P=[(50mg/100ml)×0.8ml/min ]/(0.32mg/100ml)=125ml/min
由此可见,从肾清除血浆中某物质的功能角度来说,乙肾的功能要比甲肾好。
清除率(clearance,C)是指肾在单位时间(一般用每分钟,min)内能将多少毫升血浆中所含的某些物质完全清除出去,这个被完全清除了某物质的血浆毫升数就长为该物质的清除率(ml/min)。其具体计算需要测量三个数值:U(尿中某物质的mg/100ml),V(每分钟尿量,ml/min),P(血浆中某物质的浓度,mg/100ml)。因为尿中该物质均来自血浆,所以,U×V=P×C 亦即 C=U×V/P
根据上式就可计算出各种物质的清除率。例如,Na+清除率的计算方法如下:测得尿量V为1ml/min,尿Na+浓度U为28mmmol/L,血浆Na+浓度P为140mmol/L,Na+清除率
C=[280mmol/L×1ml/min ]/(140mmol/L)=2ml/min
表示肾每分钟清除了2ml血浆中所含的所有Na+。各种物质的清除率并不一样。例如,葡萄糖的清除率为0,因为尿中不含葡萄糖(U=0mg/100ml);而尿素则为70ml/min,等等。因此,清除率能够反映肾对不同物质的清除能力。通过它也可了解肾对各种物质的排泄功能,所以它是一个较好的肾功能测量方法。
这里需要指出,所谓每分钟被完全清除了某物质的血浆毫升数,仅是一个推算的数值,实际上,肾并不一定把某1ml血浆中的某物质完全清除掉,而可能仅仅清除其中的一部分。但是,肾清除该物质的量可以相当于多少毫升血浆中所含的该物质的量。所以说,清除率所表示的血浆毫升数是一个相当量。
二、测定清除率的理论意义
测定清除率不仅可以了解肾的功能,还可能测定肾小球滤过率、肾血流量和推测肾小管转运功能。
(一)测定肾小球滤过率
肾小球滤守率可通过测定菊粉清除率和内生肌酐清除率等方法来测定。
1.菊粉清除率 肾每分钟排出某物质的量(U×V)应为涌小球滤过量与肾小管、集合管的重吸收量和分泌量的代数和。设肾小球滤过率为F;肾小囊囊腔超滤液中某物质(能自由滤过的物质)的浓度(应与血浆中的浓度一致)为P;重吸收量为R;分泌量为E。则U×V=F×P-R+E。如果某物质可以自由滤过,而且既不被重吸收(R=0)也不被分泌(E=0),则U×V=F×P,就可算出肾小球滤过率F。菊粉(inulin,也称菊糖)是符合这个条件的物质,所以它的清除率就是肾小球滤过率。
∵ U×V=F×P
∴ F=U×V/P=C
前文已提出,肾小球滤过率约为125ml/min。这个数值就是根据菊粉的清除率测得的。例如,静脉滴注一定量的菊粉以保持血浆浓度恒定,然后分别测得每分钟尿量(V)为1ml/min,尿中菊粉浓度(U)为125mg/100ml,血浆中菊粉浓度(P)为1mg/100ml,菊粉清除率可用下式计算:
C=U×V/P=[(1ml/min)×(125mg/100ml)]/(1mg/100ml)=125ml/min
所以,肾小球滤过率为125ml/min。
2. 内生肌酐清除率 由于菊粉清除率试验操作复杂,临床上改用较为简便的内生肌酐清除率试验,也以较准确地测得肾小球滤过率。所谓骨生肌酐,是指体内组织代谢所产生的肌酐。试验前二、三日,被试者禁食肉类,以免从食物中摄入过多的外来肌酐。其它饮食照常,但要避免强烈运动或体力劳动,而只从事一般工作。在这种情况下,受试者血浆中的肌酐浓度(平均在1mg/L左右)以及在一昼夜内肌酐的尿中排出总量都比较稳定。这样,在进行肌酐清除率试验时,就不必另给肌酐溶液,只需从第三天清晨起收集24小时的尿,合并起来计算其尿量,并测定混合尿中的肌酐浓度。抽到少量静脉血,测定血浆中的肌酐浓度,按下式可算出24小时的肌酐清除率。
肌酐清除率=[尿肌酐浓度(mg/L)×24小时尿量(L)/血浆肌酐浓度(mg/L)](L/24h)
肌酐能自由通过肾小球滤过,在肾小管中很少被重吸收,但有少量是由近曲小管分泌的。给正常人滴注肌酐,使血浆中浓度高达10-100mg/100ml时,近曲小管分泌肌酐的量增多,此时肌酐清除率可大于菊粉清除率,达175mg/ml。内生肌酐在血浆中的浓度相当低(仅0.1mg/100ml),近曲小管分泌的肌酐量可忽略不计,因此内生肌酐清除率与菊粉清除率相近,可以代表肾小球滤过率。然而,由于测定方法(用苦味酸显色)上的问题,实际测得的数据一般偏低。我国成人内生肌酐清除率平均为128L/24h。
(二)测定肾血流量
如果血浆中某一物质,在经过肾循环一周后可以被完全清除(通过滤过和分泌),亦即在肾动脉中该物质有一定浓度,但在肾静脉中其浓度接近于0,则该物质每分钟的尿中排出量(U×V),应等于每分钟通过肾的血浆中所含的量。设每分钟通过肾的血浆量为X,血浆中该物质浓度为P,即U×V=X×P,则该物质的清除率即为每分钟通过肾的血浆量。
U×V=X×P
C=U×V/P=X
如果在静脉滴注碘锐特(diodrast)或对氨基马尿酸(PAH)的钠盐后维持血浆浓度较低时(1-3mg/100ml),那末当它流经肾时,一次就能被肾几乎全部清除掉,因此,肾静脉中的浓度将接近于0(实际不是0,因为有部分血流通过肾的非泌尿部分)。因此,用此两种物质测得的清除率平均为660ml/min,这一数值代表了肾血浆流量。前述滤过分数就是根据肾小球滤过率和肾血浆流量来推算的。例如
滤过分数=125/660×100≌19%
如果血浆量占全血量的55%,则肾血流量=660/55×100=1200ml/min,约占心输出量的1/5-1/4。
供应肾的血液量应包括供应肾的泌尿部分和非泌尿部分(如肾被膜、肾盂等),而上述测得的肾血浆流量仅代表供应泌尿部分的数值,因此应称为肾有效血浆流量和肾有效血流量。
(三)推测肾小管的功能
通过肾小管滤过率的测定,以及其他物质清除率的测定,可以推测出哪些物质能被肾小管重吸收,哪些物质能被肾小管分泌。
例如,可以自由通过滤过膜的物质,如尿素和葡萄糖,它们的清除率均小于125ml/min(肾小球滤过率),尿素为70ml/min,而葡萄糖为0。这必定是该物质滤过之后遭到了重吸收,其清除率才能小于125ml/min。但是,不能由此而推断说该物质不会被分泌,因为只要重吸收量大于分泌量,其清除率仍可小于125ml/min。
一种物质清除率大于125ml/min(如肌酐的清除率可达175ml/min),这表明这时肾小管必定能分泌该物质,否则基清除率决不可能大于肾小球滤过率。但是,不能由此推断说该物质不会被重吸收,因为只要分泌量大于重吸收量,其清除率仍可大于125ml/min。
(四)自由水清除率
自由水清除率(free water clearance,CH2o),是指单位时间内必须从尿中除去或加入多少容积的纯水(即无溶质的水或称自由水)才能使尿液与血浆等渗,它是定量肾排水能力的指标。例如在水利尿进,血浆渗透浓度(Posm)下降,肾排出大量的低渗尿,尿液渗透浓度(Uosm)小于血奖学金渗透浓度,此时自由水清除率就表示血浆中有一定量的纯水被肾排到等渗尿中,才使尿液稀释和血浆渗透浓度回升。当缺水时,血浆渗透浓度升高,肾排出量少的高渗尿,尿液的渗透浓度大于血浆渗透浓度,此时自由水清除率就表示肾小排出一定量的纯水。这部分纯水保留在血浆,才使尿液浓缩和血浆渗透浓度回降。值得指出的是,血浆中并无真正的自由水存在,自由水清除率是计算出来的。
自由水清除率CH2O=V-Cosm (1)
V为每分钟尿量,Cosm为渗透物质清除率,按下式计算
Cosm=Uosm×V/Posm(2),代入(1)式得
CH2O=V-Uosm×V/Posm=V(1-Uosm/Posm)(3)
由(3)式可见,在等渗尿时,Uosm=Posm,CH2O=0,表示无自由水清除。当在缺水时,Uosm为1100mOsm/kgH2O,尿量V为0.5ml/min,血浆渗透浓度Posm称升高为308mOsm/kgH2O,那么Cosm=1100×0.5/308=1.79ml/min。CH2O=0.5-1.79=-1.29ml/min。自由水清除率为负值,表示肾排出的是浓缩尿、且量少,意味着肾从等渗尿中除去1.29ml/min纯水加入到血浆中,才使尿液浓缩和血浆渗透浓度下降。当水利尿时,尿渗透浓度Uosm为27mOsm/kgH2O,尿量V为20ml/min,血浆渗透浓度Posm为298mOsm/kgH2O,那么Cosm=27×20/298=1.81ml/min,CH2O=20-1.81=18.1ml/min,自由水清除率为正值,表示肾排出大量稀释尿,意味着肾从血浆中除去18.1ml/min纯水加入到等渗尿液中,才使尿液稀释和血浆渗透浓度升高。
第七节 尿的排放
尿的生成是个连续不断的过程。持续不断进入肾盂的尿液,由于压力差以及肾盂的收缩而被送入输尿管。输尿管中的尿液则通过输尿管的周期性蠕动而被送入到膀胱。但是,膀胱的排尿(micturition)是间歇地进行的。尿液在膀胱内贮存并达到一定量时,才能引起反射性排尿动作,将尿液经尿道排放于体外。
一、膀胱与尿道的神经支配
膀胱逼尿肌和内括约肌受交感神经和副交感神经支配。由2-4骶髓发出的盆神经中含副交感神经纤维,它的兴奋可使逼尿肌收缩、膀胱内括约肌松驰,促进排尿。交感神经纤维是由腰髓发出,经腹下神经到达膀胱。它的兴奋则使逼尿肌松驰、内括约肌收缩,阻抑尿的排放。但在排尿活动中交感神经的作用比较次要。
膀胱外括约肌受阴部神经(由骶髓发出的躯体神经)支配,它的兴奋可使外括约肌收缩。这一作用受意识控制。至于外括约肌的松驰,则是阴部神经活动的反射性抑制所造成的。
上述三种神经也含有传入纤维。膀胱充胀感觉的传入纤维在盆神经中;传导膀胱痛觉的纤维在腹下神经中;而传导尿道感觉的传入纤维在阴部神经中(图8-22)。
图8-22 膀胱和尿道的神经支配
二、排尿反射
在正常情况下,膀胱逼尿肌在副交感神经紧张冲动的影响下,处于轻度收缩状态,使膀胱内压经常保持在0.98kPa(10cm H2O),因为膀胱具有较大的伸展性,导致内压稍升高后可以很快回降。当尿量增加到400-500ml时膀胱内压才超过0.98kPa(10cm H2O)而明显升高(图8-23)。如果膀胱内尿量增加到700ml,膀胱内压随之增加至3.43kPa(35cm H2O)时,逼尿肌便出现节律性收缩,排尿欲也明显增加,但此时还可有意识地控制排尿。当膀胱内压达到6.86kPa(70cm H2O)以上时,便出现明显的痛感以致不得不排尿。
图8-23 人膀胱充盈过程中膀胱容量与压力的关系
图中压力垂直降低,表示容量恒定时膀胱的适应程度
排尿活动是一种反射活动。当膀胱尿量充盈到一定程度时(400-500ml),膀胱壁的牵张感受器受到刺激而兴奋。冲动沿盆神经传入,到达骶髓的排尿反射初级中枢;同时,冲动也到过脑干和大脑皮层的排尿反射高位中枢,并产生排尿欲。排尿反射进行时,冲动沿盆神经传出,引起逼尿肌收缩、内括约肌松驰,于是尿液进入后尿道。这时尿液还可以刺激尿道的感受器,冲动沿阴部神经再次传到脊髓排尿中枢,进一步加强其活动,使外括约肌开放,于是尿注被强大的膀胱内压(可高达14.7kPa(150cm H2O)驱出。尿液对尿道的刺激可进一步反射性地加强排尿中枢活动。这是一种正反馈,它使排尿反射一再加强,直至尿液排完为止。在排尿末期,由于尿道海绵体肌肉收缩,可将残留于尿道的尿液排出体外。此外,在排尿时,腹肌和膈肌的强大收缩也产生较高的腹内压,协助克服排尿的阻力。
大脑皮层等排尿反射高位中枢能对脊髓初级中枢施加易化或抑制性影响,以控制排尿反射活动。小儿大脑发育未臻完善,对初级中枢的控制能力较弱,所以小儿排尿次数多,且易发生在夜间遗尿现象。
排尿或贮尿任何一方发生障碍,均可出现排尿异常,临床上常见的有尿频,尿潴留和尿失禁。排放次数过多者称为尿频,常常是由于膀胱炎症或机械性刺激(如膀胱结石)而引起的。膀胱中尿液充盈过多而不能排出者称为尿潴留。尿潴留多半是由于腰骶部脊髓损伤使排尿反射初级中枢的活动发生障碍所致。但尿流受阻也能造成尿潴留。当脊髓受损,以致初级中枢与大脑皮层推动功能联系时,排尿便失去了意识控制,可出现尿失禁。
参考资料
1.徐丰彦,张镜如主编.人体生理学,北京:人民卫生出版社,1989
2.何小瑞,姚泰.管球反馈对肾小球血流动力学的影响及其机制.生理学进展 1991;22:216-220
3.潘敬运.肾神经的功能.生理科学进展 1985;16:215-219
4.Berne RM,Levy MN. Physiology 3rded, pp 719-753,CV Mosby Co,St Louis,1993
5.Stokes JB.Electroneutral NaCItransport in the distal tubule.Kidney International 1989;36:427
6.Rose BD.Diuretics.KidneyInternational 1991;39:336
7.Doucet A.Na-K-ATPase in thekidney tubule in relation to natriuresis.Kidney International 1992;41:Sll8
8.Sands JM,KokkoJP.Countercurrent system.Kidney International 1990;38:695
9.Molony DA,Reeves WB,AndreoliTE.Na+: K+;2CI-cotransport and thick ascendinglimb.kidney International 1989;36:418
10.Stokes JB.Sodium and potassiumtransport by the collecting duct.Kidney International 1990;38:678
第九章 感觉器官
第一节 概述
一、感受器、感觉器官的定义和分类
感受器是指分布在体表或组织内部的一些专门感受机体内、外环境改变的结构或装置。感受器的组成形成是多种多样的:有些感受器就是外周感觉神经末稍本身,如体表或组织内部与痛觉感受有关的游离神经末稍;有的感受器是裸露在神经末稍周围再包绕一些特殊的、由结缔组织构成的被膜样结构;但是对于一些与机体生存密切相关的感觉来说,体内存在着一些结构和功能上都高度分化了的感受细胞,它们以类似突触的形式直接或单位同感觉神经末稍相联系,如视网膜中的视杆和视锥细胞是光感受细胞,耳蜗中的毛细胞是声波感受细胞等,这些感受细胞连同它们的非神经性附属结构,构成了各种复杂的感觉器官如眼、耳等。高等动物中最重要的感觉器官,如眼、耳、前庭、嗅、味等器官,都分布在头部,称为特殊感管。
机体众多的感受器有不同的方法来分类。如根据感受器的分布部位,可分为内感受器和外感受器;根据感受器所接受刺激的性质,可分为光感受器、机械感受器、温度感受器和化学感受器等;便更常用的是结合刺激物和它们所引起的感觉或效应的性质来分类,据此所能区分出的人体的主要感觉类型和相应的感受器如表9-1所示。
表9-1 人体的主要感觉类型
| 感觉类型 | 感受器结构 | 感觉类型 | 感受器结构 |
| 视觉 | 视杆和视锥细胞 | 关节位置和运动觉 | 神经末稍 |
| 听觉 | 毛细胞 | 肌肉长度 | 神经末稍(肌梭) |
| 嗅觉 | 嗅神经元 | 肌肉张力 | 神经末稍(腱器官) |
| 味觉 | 味感受细胞 | 动脉血压 | 神经末稍 |
| 旋转加速度 | 毛细胞(三半规管) | 肺扩张 | 神经末稍 |
| 直线加速度 | 毛细胞(椭圆囊和球囊) | 头部血液温度 | 下丘脑某些神经元 |
| 触-压觉 | 神经末稍 | 动脉氧分压 | 神经末稍(?) |
| 温觉 | 神经末稍 | 脑脊液pH值 | 延髓腹外侧区感受器 |
| 冷觉 | 神经末稍 | 血浆葡萄糖 | 下丘脑某些细胞 |
| 痛觉 | 游离神经末稍 | 血浆渗透压 | 下丘脑前部某些细胞 |
表中只有前11项是通常能引起主观感觉的,其余的感受器一般只是向中枢神经系统提供内、外环境中某些因素改变的信息,引起各种调节性反应,但这时在主观上并不产生特定的感觉。
二、感受器的一般生理特性
(一)感受器官适宜刺激
各种感受器的一个共同功能特点,是它们各有自己最敏感、最容易接受的刺激形式;这就是说,用某种能量形式的刺激作用于某种感受器时,只需要极小的强度(即感觉阈值)就能引起相应的感觉。这一刺激形式或种类,就称为该感受器的适宜刺激,如在一定波长的电磁波是视网膜光感受细胞的适宜刺激,一定频率的机械震动是蜗毛细胞的适应刺激等。正因为如此,机体内、外环境中所发生的各种形式的变化,总是先作用于和它们相对应的那种感受器。这一现象的存在,是因为动物在长期的进化过程中逐步形成了具有各种特殊结构和功能的感受器以及相应的附属结构的结果,便得它们有可能对内、外环境中某些有意义的变化进入灵敏的感受和精确的分析。不同动物所处的生活环境和条件不同,因此在进化中有可能形成一些异于人体的特殊感受装置,这在广大的动物界屡见不鲜,早已引起人们极大的兴趣和注意。研究这些可能是极低等动物的特殊感受装置,不仅对理解感受器活动的一般规律有帮助,而且有很大的仿生学意义。
(二)感受器的换能作用
各种感受器在功能上的另一个共同特点,是能把作用于它们的各种刺激形式,转变成为相应的传入神经末稍或感受细胞的电反应,前者称为发生器电位(generator potential),在后者称为感受器电位(receptorpotential)。如在第二章所述,发生器电位和感受器电位的出现,实际上是传入纤维的膜或感受细胞的膜进行了跨膜信号传递或转换过程的结果。和体内一般细胞一样,所有感受器细胞对外来不同刺激信号的跨膜转换,也主要是通过两种基本方式进行的,如声波振动的感受与蜗毛顶部膜中与听毛受力有关的机械细胞对外来中与听毛受力有关的机械门控通道的开放和关闭有关,这使毛细胞出现与声波振动相一致的感受器电位(即微音器电位);视杆和视锥细胞则是由于它们的外段结构中视盘膜上存在有受体蛋白(如视紫红质),它们在吸收光子后,再通过特殊的G-蛋白和作为效应器酶的磷酸二酯酶的作用,引起光感受器细胞外段胞浆中cGMP的分解,最后使外段膜出现感受器电位。在其他一些研究过的感受器,也看到了类似的两种信号转换机制。由此可见,所有感受性神经末稍和感受器细胞出现电位变化,就是通过跨膜信号转换,把不同能量形式的外界刺激都转换成跨膜电位化的结果。
如前,发生器电位和感受器电位同终板电位和突触后电位一样,是一种过渡性慢电位,它们不具有“全或无”的特性而其幅度与外界刺激强度成比例;它不能作远距离传播而可能在局部实现时间性总和和空间性总和。正因为如此,感受器电位和发生器电位的幅度、持续时间和波动方向,就反映了外界刺激的某些特征,也就是说,外界刺激信号所携带的信息,也在换能过程中转移到了这种过渡性电变化的可变动的参数之中。
发生器电位和感受器电位的产生并不意味着感受器功能作用的完成,只有当这些过渡性电变化最终触发分布在该感受器的传入神经纤维上产生“全或无”式的可作远距离传导的动作电位序列时,才标志着这一感受器或感觉器官作用的完成;但主观感受的产生、对外界刺激信号的精细分析以及最后引起整个机体出现应答性反应和信息贮存等过程,则是传入神经纤维所输的神经信号到达各级脑中枢以后的反应,不属本章的内容。
(三)感受器的编码作用
感受器在把外界刺激转换成神经动作电位时,不仅仅是发生了能量形式的转换;更重要的是把刺激所包涵的环境变化的信息,也转移到了新的电信号系统即动作电位的序列之中,即编码作用。编码(encoding)一词,本是工程通讯理论中的一个概念,指一种信号系统(如莫尔斯电码)如何把一定的信息内容(如电文内容)包涵在少量特定信号的排列组合之中。因此,感受器将外界刺激转变成神经动作电位的序列时,同时也实现了编码作用;中枢就是根据这些电信号序列才获得对外在世界的认识的。问题是外界刺激的质和量以及其他属性,是如何编码在特有的电信号序列中的?这一问题十分复杂,目前还远远没有弄清楚,下面先从最简单的方面加以叙述。
首先考虑外界刺激的“质”,如听觉或视觉等刺激在性质上的不同是如何编码的。如所周知,不论来自何种感受器的传入神经纤维上的传入冲动,都是一些在波形和产生原理上基本相同的动作电位;例如,由视神经、听神经或皮肤感觉神经的单一纤维上记录到的动作电位,并无本质上的差别。因此,不同性质的外界刺激不可能是通过某些特异的动作电位波形或强度特性来编码的。实验和临床经验都表明,不同种类的感觉的引起,不但决定于刺激的性质和被刺激的感受器,也决定于传入冲动所到达的大脑皮层的终端部位。例如,用电刺激作用于病人视神经,使它人为地产生传向枕叶皮层的传入冲动,或者直接刺激枕叶皮层使之产生兴奋,这时都会引起光亮的感觉,而且主观上感到这些感觉是发生在视野的某一部位;同样,临床上遇到肿瘤或炎症等病变刺激听神经时,会产生耳鸣的症状,这是由于病变刺激引起的神经冲动传到了皮层听觉中枢所致;而某些痛觉传导路或相应中枢的刺激性病变,也会引起身体一定部位的疼痛。这些都说明,感觉的性质决定于传入冲动所到达的高级中枢的部位,而不是由于动作电位的波形或序列特性有什么不同;也就是说,不同性质的感觉的引起,首先是由传输某些电信号所使用的通路来决定的,即由某一专用路线(labeled line)传到特定终端部位的电信号,通常就引起某种性质的主观感觉。事实上,即使是同一性质的刺激范围内,它们的一些次级属性(如视觉刺激中不同波长的光线和听觉刺激中不同频率的振动等)也都有特殊分化了的感受器和专用传入途径。在自然状态下,由于感受器细胞在进化过程中的高度分化,使得某一感受细胞变得对某种性质的刺激或其属性十分敏感,而由此产生的传入信号又只能循特定的途径到达特定的皮层结构,引起特定性质的感觉。因此,一般无需怀疑,某种主观感觉是否是由一些非适宜刺激引起的“非真实”的感觉,只是在病理情况下有例外。
在同一感受系统或感觉类型的范围内,外界刺激的量或强度是怎样编码的呢?既然动作电位是“全或无”式的,因而刺激的强度不可能通过动作电位的幅度大小或波形改变来编码。根据在多数感受器实验中得到的实验资料,刺激的强度是通过单一神经纤维上冲动的频率高低和参加这一信息传输的神经张纤维的数目的多少来编码的。图9-1表示在人手皮肤的触压感受器所进行的实验,说明在感受器的触压重量和相应的传入纤维的动作电位发放频率之间,存在着某种对应关系。重量过轻时,神经纤维全无反应,到达感受阈值时开始有冲动产生;以后随着触压重量的增大,传入纤维上的冲动频率也越来越高。不仅如此,在触压刺激继续加大的情况下,同一刺激有可能引起较大面积的皮肤变形,使一个以上的感受器和传入纤维向中枢发放冲动。这样,刺激的强度既可通过每一条传入纤维上冲动频率的高低来反映,还可通过参与电信号传输的神经纤维的数目的多少来反映。当然,任何一个天然刺激在空间和时间上的属性都是极其复杂的(例如一个彩色电影画面所包涵的信息内容),因此,感受器的编码过程也是极其复杂的。还应该知道的是,感觉过程的编码过程并不只是感受器部位进行一次,事实上信息每通过一次神经元间的突触传递,都要进行一次重新编码,这使它有可能接受来自其他信息源的影响,使信息得到不断的处理,这当然属于中枢神经元网络的功能。
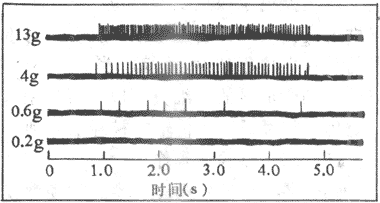
图9-1 不同重量的触压刺激是在单一传入纤维上引起的冲动频率的改变
至于刺激的物理强度如何转变成为传入神经纤维上频率不同的冲动,目前认为是由于强的刺激能引起幅度较大而持续时间较长的发生器电位,而后者引起神经末稍较高频率的冲动。
(四)感受器的适应现象
当刺激作用于感觉器时,经常看到的情况是虽然刺激仍在继续作用,但传入神经纤维的冲动频率已开始下降,这一现象称为感受器的适应(adaptation)。适应是所有感受器的一个功能特点,但它出现的快慢在不同感受器有很大的差别,通常可把它们区分为快适应和慢适应感受器两类。快适应感受器以皮肤触觉感受器为代表,当他们受刺激时只顾刺激开始后的短时间内有传入冲动发放,以后刺激仍然在作用,但传入冲动频率可以逐渐降低到零;慢适应感受器以肌梭、颈动脉窦压力感受器为代表,它们在刺激持续作用时,一般只是在刺激开始以后不久出现一次冲动频率的某些下降,但以手可以较长时间维持在这一水平,直至刺激撤除为止。感受器适应的快慢各有其生理意义,如触觉的作用一般在于探索新异的物体或障碍物,它的快适应有利于感受器及中枢再接受新事物的刺激;慢适应感受器则有利于机体对某些功能状态如姿势、血压等进行长期持续的监测,有利于对它们可能出现的波动进行随时的调整。适应并非疲劳,因为对某一刺激产生适应之后,如增加此刺激的强度,又可以引起传入冲动的增加。
感受器产生适应的机制比较复杂,有的发生在刺激引起发生器电位这一阶段;有的发生在发生器电位诱发神经动作电位这一阶段。有不少感受器,适应发生的快慢一感受末稍所具有的附属结构有关。一个有趣的例子是,作为触压感受器的皮肤(如肠系膜)环层小体,其环层结构的存在与它适应有快速出现有关:实验中如果细心剥除环层结构后,直接轻压裸露的神经末稍仍可引起传入冲动发放,而且在这种情况下感受末稍变得不易适应,与剥除环层结构前表现的快适应明显不同。这个现象的解释是,当压力直接作用于环层结构表面时,压力要经过此结构才能传递到感受末稍表面,但因为环层结构具有一定的弹性,它受压后的弹性变形和回弹有可能使末稍表面实际受到的压力减轻或消失,使刺激的实际作用减弱以至全不起作用。
在人体的主观感受方面,也常常体验到类似“入芝兰室,久而不闻其香”之类的感觉适应现象。感觉适应的产生机制可能更为复杂,其中只部分地与感受器的适应有关,因为适应的产竹敢与传导途径中的突触传递和感觉中枢的某些功能改变有关。
第二节 视觉器官
引起视觉的外周感受器官是眼,它由含有感光细胞的视网膜和作为附属结构的折光系统等部分组成。人眼的适宜刺激是波长370-740nm的电磁波;在这个可见光谱的范围内,人脑通过接受来自视网膜的传入信息,可以分辨出视网膜像的不同亮度和色泽,因而可以看清视野内发光物体工反光物质的轮廓、形状、颜色、大小、远近和表面细节等情况。自然界形形色色的物体以及文字、图形等形象,通过视觉系统在人脑得到反映。据估计,在人脑获得的全部信息中,大约有95%以上来自视觉系统,因而眼无疑是人体最重要的感觉器官。
人眼的基本结构如图9-2所示。除了控制眼球运动的眼外肌和起保持、营养作用的巩膜、脉络膜等结构外,眼内与视觉传入信息的产生直接有关的功能结构,是位于眼球正中线上的折光系统和位于眼球后部的视网膜。由角膜经房水、晶状体、玻璃体直至视网膜的前表面,都是一些透明而无血管分布的组织,它们构成了眼内的折光系统,使来自眼外的光线发生折射,最后成像在视网膜上,视网膜具有同神经组织类似的复杂结构,其中包含有对光刺激高度敏感的视杆和视锥细胞,能将外界光刺激所包含的视觉信息转变成为电信号,并在视网膜内进行初步处理,最后以视神经纤维的动作电位的形式传向大脑。因此,形容眼的功能首先要研究眼内折光系统的不学特性,搞清楚它们怎样能把不同远近的物体成像在视网膜上以及形成清晰物像的限度;其次要阐明视网膜是怎样对视网膜成像进行换能和编码的。
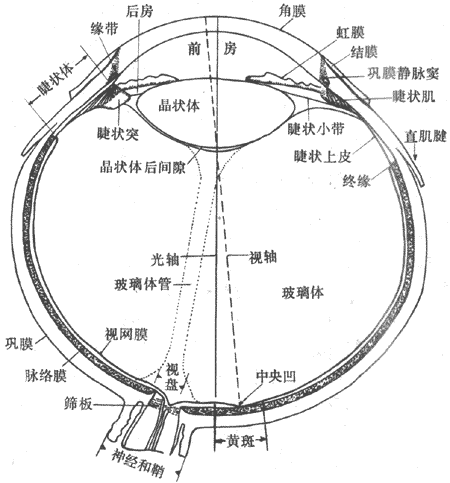
图9-2 眼球的水平切面(右眼)
一、眼的折光系统及其调节
当光线由 空气进入另一媒质构成的单球面折光体时,它进入物质的折射情况决定于该物质与空气界面的曲率半径R和该物质的折光指数n2;若空气的折光指数为n1,则关系式为
n2R/(n2-n1)=F2(1)
F2称为后主焦距或第2焦距(空气侧的焦距为前主焦距或第一焦距),指由折射面到后主焦点的距离,可以表示这一折光的折光能力。表示折光体的折光能力还可用另一种方法,即把主焦距以m(米)作单位来表示,再取该数值的倒数,后者就称为该折光体的焦度(diopter);如某一透镜的主焦距为10cm,这相当于0,1m,则该透镜的折光能力为10焦度(10D)。通常规定凸透镜的焦度为正值,凹透镜的焦度为负值。
主焦距是一个折光体最重要的光学参数,由此可算出位于任何位置的物体所形成的折射像的位置。以薄透镜为例,如果物距α是已知的,像距b可由下式算出:
1/a+1/b=1/F2(2)
由式(2)可以看出,当物距a趋于无限大时,1/a趋近于零,于是1/b接近于1/F2,亦即像距b差不多和F2相等;这就是说,当物体距一个凸透镜无限远时,它成像的位置将在后主焦点的位置。同样不难看出,凡物距小于无限大的物体,它的像距b恒大于F2,即它们将成像在比主焦点更远的地方。以上两点结论,对于理解眼的折光成像能力十分重要。
另外,根据光学原理,主焦点的位置是平行光线经过折射后聚焦成一点的位置,这一结论与上面提到的第一点结论相一致。每一物体的表面,都可认为是由无数的发光点或反光点组成,而由每一个点发出的光线都是辐散形的;只有这些点和相应的折射面的距离趋于无限大时,由这些点到达折射面的光线才能接近于平行,于是它们经折射后在主焦点所在的面上聚成一点,整个物质就达个面上形成物像。当然,无限过的概念本身决定了它是一个不可能到达的位置,实际上对人眼和一般光学系统来说,来自6m以外物体的各光点的光线,都可以认为是近于平行的,因而可能在主焦点所在的面上形成物像。
(二)眼的折光系统的光学特性
当用上述光学原理分析眼的折光特性时,首先遇到的一个困难是,眼球并非一个薄透镜或单球面折光体,而是由一系列由率半径和折光指数都不相同的折光体所组成的折光系统。显然,人眼折光系统的后主焦距不能简单地由式(1)算出,不过它的最主要的折射发生在角膜,而按几何学原理进行较复杂的计算,还是可以追踪出光线经眼内多个折光面行进的途径,并得出由这些组合的透镜组所决定的后主焦点的所在位置。
计算结果表明,正常成人眼处于安静而不进行调节的状态时,它的折光系统的后主焦点的位置,正好是其视风膜所在的位置。这一解剖关系对于理解正常眼的折光成像能力十分重要。它说明,凡是位于眼前方6m以外直至无限远处的物体,根据式(2)或由于由它们发出或反射出的光线在到达眼的折光系统时已近于平行,因而都可以在视网膜上形成基本清晰的像,这正如放置于照相机主焦点处的底片,可以拍出清晰的远景一样。当然,人眼不是无条件的看清任何远处的特体,例如,人眼可以看清楚月亮(或其他更远的星体)和它表面较大的阴影,但不能看清楚月球表面更小的物体或特征。造成后一限制的原因是,如果来自某物体的光线过弱,或它们在空间处女内传播时被散射或吸收,那么它们到达视网膜时已减弱到不足以兴奋感光细胞的程度,这样就不可能被感知;另外,如果物体过小或它们离眼的距离过大,则它们在视网膜上形成的大小,将会小到视网膜分辨能力的限度以下,因而也不能感知。
(三)眼的调节
如果安静状态的眼的折光能力正好把6m以外的物体成像在视网膜上,那么来自较6m为近的物体的光线将是不同程度呈辐射状的,它们在折射后的成像位置将在主焦点,亦即视网膜的位置之后;由于光线到达视网膜时尚未聚焦,因而物像是模糊的,由此也只能引起一个模糊的视觉形象。但正常眼在看近特时也十分清楚,这是由于眼在看近物时已进行了调节(accommodation),使进入眼内的光线经历较强的折射,结果也能成像在视网膜上。人眼的调节亦即折光能力的改变,主要是靠晶状体形状的改变;这是一个神经反射性活动,其过程如下:当模糊的视觉形象出现在视区皮层时,由此引起的下行冲动经锥体束中的皮层-中脑束到达中脑的正中核,再到达发出动眼神经中副交感节前纤维的有关核团,最后再经睫状神经节到达眼内睫状肌,使其中环行肌收缩,引起连接于水晶体囊的悬韧带放松;这样就促使水晶体由于其自身的弹性而向前方和后方凸出(以前突较为明显),使眼的总的折光能力较安静时增大,使较辐射的光线提前聚焦,也能成像在视网膜上。因9-3表示调节前后晶状体形状的改变。很明显,物体距眼球愈近,到达眼的光线辐散程度愈大,因而也需要晶状体作更大程度的变凸。调节反射进行时,除晶状体的变化外,同时还出现瞳孔的缩小和两眼视轴向鼻中线的会聚,前者的意义在于减少进入眼内光线的量(物体移近时将有较强光线到达眼球)和减少折光系统的球面像差和色像差;两眼会聚的意义在于看近物时物像仍可落在两眼视网膜的相称位置。
图9-3 眼调节前后睫状体位置和晶状体形状的改变
实线为安静时的情况,虚线为看近物经过调节后的情况,注意晶状体的前凸比后凸明显
人眼看近物的能力,亦即晶状体的调节能力是有一定限度的,这决定于水晶体变凸的最大限度。随着年龄的增加,水晶体自身的弹性将下降,因而调节能力也随年龄的增加而降低。眼的最大调节能力可用它所能看光天化日物体的最近距离来表示,这个距离或限度称为近点。近点愈近,说明晶状体的弹性愈好,亦即它的悬韧带放松时可以作较大程度的变凸,因而使距离更近的物体也能成像在视网膜上。例如,8岁左右的儿童的近点平均约8.6cm,20岁左右的成为约为10.4cm,而60岁时可增大到83.3cm。
(四)简化眼和视敏度
由于眼内有多个折光体,要用一般几何光学的原理画出光线在眼内的行进途径和成像情况时,显得十分复杂。因此,有人根据眼的实际光学特性,设计一些和正常眼在折光效果上相同、但更为简单的等效光学系统或模型,称为简化眼。简化眼只是一种假想的人工模型,但它的光学参数和其它特性与正常眼等值,故可用来分析眼的成像情况和进行其他计算。常用的一种简化眼模型,设想眼球由一个前后径为20mm的单球面折光体构成,折光指数为1.333;外界光线只在由空气进入球形界面时折射一次,此球面的曲率半径为5mm,亦即节点在球形界面后方5mm的位置,后主焦点正相当于此折光体的后极。显然,这相模型和正常安静的人眼一样,正好能使平行光线聚焦在视网膜上(图9-4)。
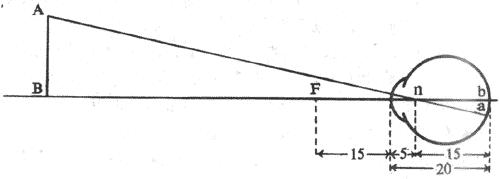
图9-4 简化眼及其成像情况
n为节点,AnB和anb是两个相似三角形;如果物距为已知,就可由物体大小算出物像
大小,也可算出两三角形对顶角(即视角)的大小
利用简化眼可以方便地计算出不远近的物体在视网膜上成像的大小。如图9-4所示,AnB和and是具有对顶角的两个相似的三角形,因而有:
![]()
其中nb固定不变,相当于15mm,那么根据物体的大小和它距眼的距离,就可算出物像的大小。此外,利用简化眼可以算出正常人眼所能看清的物体的视网膜像大小的限度。检查证明,正常人眼即使在光照良好的情况下,如果视网膜小于5μm,一般就不能引起清晰的视觉。这说明,正常人的视力或视敏度(visualacuity)有一个限度;要表示这个限度,只能用人所能看清的最小视网膜的大小,而不能用所能看清的物体大小表示,因为物像有大小与物体的大小有关,大致相当于视网膜中央凹处一个视锥细胞的平均直径(但有些视锥的直径可小于2μm)。
通常用业检查视敏度的国际通用的视力表,就是近上述原理设计的。当人眼能看清5m处的一个圆形或E字形上相距1.5mm的缺口的方向时,按简化眼计算,此缺口在视网膜像中的距离约为5μm(实际计算值为 4.5μm),说明此眼视力正常,定为1.0;由图9-4也可以算出,当物像为5μm时,由光路形成的两个三角形的对顶角即视角约相当于1分度(即1');因此,如果受试者在视角为10分分度时才能看清相应增大了视力表上的标准图形的缺口(相当于国际视力表上最上面一排图),则视力定为0.1;在表上还列出视力0.2至0.9时的逐步减小的图形;但国际视力表上对这些相应图形的大小设计是有缺点的,如相当于0.2视力的图形比视力0.1的图形小1/2,而相当于视力1.0的图形只比视力为0.9时的图形小了1/9。这种表示视力方法显然不利于临床上表示视力的改善程度,例如由原来0.9的视力改善为1.0,或由0.1的视力改善为0.2,虽然视力都增加了0.1,但其真正改善的程度并不一样,因而不能作为统计处理的数据。为了避免这一缺点,我国有人设计了一种对数视力表(缪天荣,1966),它把国际视力表上记为1.0的正常视力记为5.0,而将视角为10分度时的视力记为4.0,其间相当于视力4.1、4.2直至4.9的图形,各比上一排形成的视角小=1.259……倍,而log=0.1;这样,视力表上不论原视力为何值,改善程度的数值都具有同样的意义。
眼的折光能力和调节能力异常 正常眼的折光系统在无需进行调节的情况下,就可使平行光线聚焦在视网膜上,因而可看清远处的物体;经过调节的眼,只要物体的距离不小于近点的距离,也能在视网膜上形成清晰的像被看清,此称为正视眼。若眼的折光能力异常,或眼球的形态异常,使平行光线不能在安静未调节的眼的视网膜上成像,则称为非正视眼,其中包括近视、远视和散光眼。有些眼静息时折光能力正常,但由于水晶体的弹性减弱或丧失,看远物时的调节能力减弱,此称为老视。
近视 多数由于眼球的前后径过长(轴性近视),致使来自远方物体的平行光线在视网膜前即已聚焦,此后光线又开始分散,到视网膜时形成扩散开的光点,以致物像模糊。便近视看近物时,因这时聚焦的位置较平行光线时为后,因而眼无需进行调节或进行较小程度的调节,就可在视网膜上成像;这就使近视能看清近物,且远点比正常眼还要近。纠正近视眼的方法是在眼前增加一个一定焦度的凹透镜片,使入眼的平行光线适当辐散,以便聚焦位置移后,正好能成像在视网膜上;这样使远物可以看清,而近物则像正常眼一样,依靠眼睛自身的调节能力。近视也可由于眼的折光能力超过正常,使平行光线成像在位置正常的视网膜之前,这种近视特称为屈光近视。
远视 由于眼球前后径过短,以致主焦点的位置实际在视网膜之后,这样入眼的平行光线在到达视网膜时尚未聚焦,也形成一个模糊的像,引起模糊的视觉。这时,患者在看远物时就需使自己的调节能力,使平行光线能提前聚焦,成像在位置前的视网膜上。由此可见,远视眼的特点是在看远物时即需动用眼的调节能力,因而看近物时晶状体的凸出差差不多已达到它的最大限度,故近点距离较正常人为大,视近物能力下降,纠正的方法是戴一适当焦度的凸透镜,使看远时不需晶状体的调节亦能在像在视网膜上,于是通过调节能力就可像正视眼一样用来看近物了。
散光 正常眼的折光系统的各折光面都是正球面的,即在球表面任何一点的曲率半径都是相等的。如果由于某些原因,折光面(通常见于角膜)在某一方位上曲率半径变小,而在与之相垂直的方位上曲率半径变大(相当于在一个硬的桌面上轻压一个乒乓球时,球面的曲率半径在垂直的方位上变小,在横的方位上变大一样),在这种情况下,通过角膜不同方位的光线在眼内不能同时聚焦,这会造成物像变形和视物不清。这种情况属于规则散光,可用适当的柱面镜纠正,后者的特点正是互相垂直方位上具有不同的曲率半径,当它和角膜的曲率半径改变大小相抵消时,使角膜的曲率异常得到纠正。
二、瞳孔和瞳孔对光反应
瞳孔指虹膜中间的开孔,是光线进入眼内的门户;它在亮光处缩小,在暗光处散大。虹膜由多单位平滑肌构成;在瞳孔周围的是环形肌层,受动眼神经中的副交感神经纤维支配,收缩时使瞳孔缩小,故又称瞳孔括约肌;虹膜的外周部分是辐散状肌纤维,受由颈部上行的交感神经纤维支配,收缩时使瞳孔散大,故又称瞳孔散大肌。瞳孔的大小可以控制进入眼内的光量。一般人瞳孔的直径可变动于1.5-8.0mm之间。假定人由光亮处进入暗室时瞳孔直径可增加5倍,那么瞳孔的受光面积应增大25倍;可见瞳孔的变化,有保持在不同光照情况下进入眼内的光量较为恒定的作用。但暗室中较强阳光照射的光照强度实际减弱约100万倍,因而单靠瞳孔大小的改变,远不足以使进入眼内的光量保持恒定。事实上,人眼在不同的亮度情况下是靠视网膜中不同的感光细胞来接受光刺激的,在暗光处起作用的视杆细胞对光的敏感程度要比在亮光处起作用的视锥细胞大得多,因此在暗处看物,只需进入眼内光量适当增加即可。由此可见,通过改变瞳孔大小以调节进入眼内的光量还是有一定意义的。
瞳孔大小随光照强度而变化的反应,是一种神经反射,称为瞳孔对光反射。引起此反射的感受器就是视网膜,传入纤维在视神经中,但这部分纤维在进入中枢后不到达外侧膝状体,而在在中脑的顶盖前区换神经元,然后到同侧和对侧的动眼神经核,传出纤维主要是动眼神经中的副交感纤维,效应器也主要是瞳孔约肌。
瞳孔对光反应的特点是效应的双侧性,即如果光照的是一侧眼睛时,除被照眼出现瞳孔缩小外,同时未受光照拐殊途同归瞳孔也缩小,后者我为互感性对光反射。临床上有时可见到瞳孔对光反应消失、瞳孔左右不等、互感性瞳孔反应消失等异常情况,常常是由于与这些反射有关的反射绵弧某一部分受损的结果,因而可以藉瞳孔反应的异常帮助进行神经病变的定位诊断。
房水和眼内压 房水指充盈于眼的前、后房中的液体,其成分类似血浆,但蛋白质含量较血浆低得多,而HCO3-含量却超过血浆;因而房水的总渗透压也较血浆为高。房水的生成部位在睫状体脉络膜丛,生成后由后房经瞳孔进入前房,再由巩膜和角膜结合处的前房角进入巩膜静脉窦,最后汇入静脉系统。房水不断生成,不断回收入静脉,使它在后房和前房之间流动不息。据测定,正常时房水的生成速度每分钟约为2mm3;由于它的生成和回收之间保持着动态平衡,使得眼内政党时的房水量保持恒定,又由于眼的前、后房容积也是相对恒定的,于是使其中静水压(即眼内压)也保持相对的稳定。据国内调查资料,我国成年人眼内压正常值为2.27-3.2kPa(17-24mmHg),平均2.67kPa(17-24mmHg)。
眼内压的相对稳定,对保持眼球特别是角膜的正常开头和折光能力有重要的意义。当眼球被刺穿时,可能导致房水流失,眼内压下降,引起眼球变形,角膜也不能保持正常的曲度。人眼的总折光能力与眼内折光体都有一定关系,但最主要的折射发生在空气与角膜接触的界面上。这约占总折光能力的80%。因此,角膜的曲度和形状的改变将显著地影响眼的折光能力,严重地影响视力。房水也对它所接触的无血管组织如角膜和晶状体起着营养的作用。房水循环障碍时会造成眼内压过高,临床上称为青光眼,可导致角膜、晶状体以及虹膜等结构的代谢障碍,严重时造成角膜混浊、视力丧失。
房水生成的机制目前尚不完全明了。通常认为,除了在睫状体脉络膜丛处的毛细血管靠被动滤过(类似在一般毛细血管的动脉端生成组织液,见第四章),使血浆中的水分和盐类透出血管壁生成房水外,还有主动过程的参与,否则就难于解释房水何以有较血浆中浓度高的HCO3-等盐类离子。用组织化学的方法证明,睫状体上皮细胞含有较多的碳酸酐酶,此酶的作用是使细胞代谢过程中产生的CO2和H2O,迅速生成H2CO3,并解离出HCO3-,后者经膜上的主动转运过程进入房水,造成它的房水中的高浓度,这个高浓度造成的负电位和高渗透压还能进一步促使血浆中的Na+和水分子进入房水。临床上可以使用碳酸酐酶抑制剂(如乙酰唑胺)减少房水生成,降低眼内压,其作用机制与上述的房水生成机制有关。
三、视网膜的结构和两种感光换能系统
来自外界物体的光线,通过眼内的折光系统在视网膜上形成物像,是视网膜内的感光细胞被刺激的前提条件。视网膜像还有一个物理范畴内的内像,用几何光学的原理可以较容易地对它加以说明,和外界物体通过照相机的中的透镜组在底片上形成的物像并无原则上的区别;但视觉系统最后在主观意识上形成的“像”,则是属于意识或心理范畴的主观印象,它由来自视网膜的神经信息最终在大脑皮层等中枢结构内形成。作为感受器生理,重点是视网膜怎样把物理像转换成视神经纤维上的神经信号,以及在这些信号的序列和组合中怎样包括了视网膜像、亦即外界物体所提供的信息内容。应该提出,视觉研究的进展虽然较快,但也只是初步的。
(一)视网膜的结构特点
视网膜的厚度只有0.1-0.5mm,但结构十分复杂。它的主要部分在个体发生上来自前脑泡,故属于神经性结构,其中细胞通过突触相互联系。经典组织学将视网膜分为十层,但按主要的细胞层次简化为四层业描述,如图9-5所示。从靠近脉络膜的一侧算起,视网膜最外层是色素细胞层;这一层的来源不属神经组织,血液供应也来自脉络膜一侧,与视网膜其他层接受来自视网膜内表面的血液供应有所不同;临床上见到的视网膜剥离,就发生在此层与其它层次之间。色素细胞层对视觉的引起并非无关重要,它含在黑色素颗粒和维生素A,对同它相邻接的感光细胞起着营养和保护作用。保护作用是除了色素层可以遮继来自巩膜侧的散射光线外,色素细胞在强光照射视网膜时可以伸出伪足样突起,包被视杆细胞外段,使其相互隔离,少受其他来源的光刺激;只有在暗光条件下,视杆外段才被暴露;色素上皮的这种活动受膜上的多巴胺受体控制。此层内侧为感光细胞层。在人类和大多数哺乳动作动物,感光细胞分视杆和视锥细胞两种,它们都含有特殊的感光色素,是真正的光感受器细胞。视杆和视锥细胞在形态上都可分为四部分,由外向内依次称为外段、内段、胞体和终足(图9-6);其中外段是感光色素集中的部位,在感光换能中起重要作用。视杆和视锥细胞在形成上的区别,也主要在外段它们外形不同,所含感光色素也不同。视杆细胞外段呈长杆状,视锥细胞外段呈圆锥状。两种感光细胞都通过终足和双极细胞层内的双极细胞发生突触联系,双极细胞一般再和节细胞层中的神经节细胞联系。视网膜中除了这种纵向的细胞间联系外,还存在横向的联系,如在感光细胞层和双极细胞层之间有水平细胞,大双极细胞层和节细胞层之间有无长突细胞;这些细胞的突起在两层细胞之间横向伸展,可以在水平方向传递信息,使视网膜在不同区域之间有可能相互影响;这些无长突细胞还可直接向节细胞传递信号。近年来发现,在视网膜还存在一种网间细胞,它的细胞体位于双极细胞层和节细胞层之间,但突起却伸到感光细胞层和双极细胞层。如果把感光细胞经过双极细胞到神经节细胞的途径,看作是视觉信息的初始阶段。近年来还发现,视网膜中除了有通常的化学性突触外,还有大量电突触存在。由此可见,视网膜也和神经组织一样,各级细胞之间存在着复杂的联系,视觉信息最初在感光细胞层换能变成电信号后,将在视网膜复杂的神经元网络中经历某种处理和改变,当视神经纤维的动作电位序列作为视网膜的最终输出信号传向中枢时,它们已经是经过初步加工和处理的信息了。
盲点 由节细胞层发出的神经轴突,先在视网膜表面聚合成一整束,然后它透视网膜,在眼的后极出眼球,这就在视网膜表面形成视神经乳头。在乳头的范围内,实际上没有视网膜特有的细胞结构,因而落于该处的光线或视网膜像的组成部分,将不可能被感知,故称为盲点。两侧视神经乳头在视网膜内黄斑或中央凹中心的鼻侧约3mm处。但正常时由于用两眼看物,一侧盲点可以被对侧视觉补偿,人们并不觉察自己的视野中有一处无视觉感受的区域。盲点的存在可用专门设计的方法来证明。
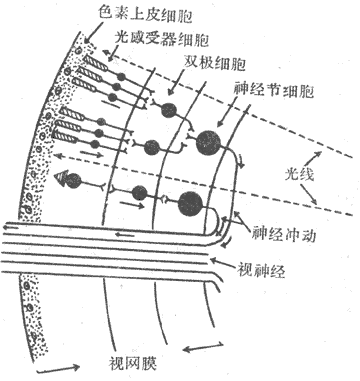
图9-5 视网膜的主要细胞层次及其联系模式图
图9-6 哺乳动物光感受器细胞模式图
(二)视网膜的两种感光换能系统
根据对视网膜结构和功能的研究,目前认为在人和大多数脊椎动物的视网膜中存在着两种感光换能系统。一种由视杆细胞和与它们相联系的双极细胞和神经节细胞等成分组成,它们对光的敏感度较高,能在昏暗的环境中感受光刺激而引起视觉,但视物无色觉而只能区别明暗;且视物时只能有较粗略的轮廓,精确性差,这称为视杆系统或晚光觉系统;另一种由视锥细胞和与它们有关的传递细胞等成分组成,它们对光的敏感性较差,只有在类似白昼的强光条例下才能被刺激,但视物时可辨别颜色,且对物体表面的细节和轮廓境界都能看得很清楚,有高分辨能力,这称为视锥系统或昼光觉系统(前述视敏度的测定实际是视锥系统视力的测定)。
证明这两种相对独立的感光-换能系统存在的主要依据是:①人视网膜中视杆和视锥细胞在空间上的分布是不均匀的,愈近视网膜周边部,视杆细胞愈多而视锥细胞愈少;愈近视网膜中心部,视杆细胞愈少而视锥细胞愈多;在黄斑中心的中央凹处,感光细胞全部是视锥而无视杆细胞;与上述细胞分布相对应,人眼视觉的特点正是中央凹在亮光处有最高的视敏度和色觉,在暗外则中央视力较差;相反地,视网膜周边部则能感受弱光的刺激,但这时无色觉而清晰度较差。②两种感光细胞和双极细胞以及节细胞形成信息传递通路时,逐级之间都有一定程度的会聚现象(参看第十章),但这种会聚在视锥系统程度较小,在中央凹处甚至可以看到一个视锥细胞只同一个双极细胞联系,而这个双极细胞也只同一个神经节细胞联系的情况,这种低程度会聚或无会聚现象的“单线联系”,显然是视锥系统有较高的精细分辨能力的结构基础;与此相对照,在视杆系统则普遍存在多个感光细胞同一个双极细胞联系,而多个双极细胞再同一个神经节细胞联系的会聚式排列;在视网膜周边部,可看到多达250个视杆细胞经少数几个双极细胞会聚于一个神经节细胞的情况;在这种情况下,当然不能期待这样的感觉系统有高的精细分辨能力。但这样的聚合系统却具有较强的总和多个弱刺激的能力。③从动物各系统特点来看,某些只在白昼活动的动物如爬虫类和鸡等,视网膜全无视杆而只胡视锥细胞。而另一些只在夜间活动的动物如地松鼠和猫头鹰等,视网膜中只胡视杆而不含视锥细胞。④视杆细胞中只含有一种感光色素,即视紫红质(rhodopsin),而视锥细胞却因所含感光色素的吸收光谱特性不同而分为三种,这是同视杆系统无色觉而视锥系统有色觉的事实相一致的。
四、视杆细胞的感光换能机制
从上世纪末开始,有人就从视网膜中提取出了一定纯度的感光色素即视紫红质,它在暗处呈红色;实验中还可以证明,提取出来的这种感光色素对不同波长光线的吸收光谱,基本上和晚光觉对光谱不同部分的敏感性曲线相一致(图9-7)。这一事实十分重要,因为既然光线对某种感光色素的光化学作用的强度正好与这些光线所引起的视觉的强度相一致,那就是提示前者可能是后者的基础。
(一)视紫红质的光化学反应及其代谢
视紫红质的分子量约为27-28kd,是一种与结合蛋白质,由一分子称为视蛋白(opsin)的蛋白质和一分子称为视黄醛(retnal)的生色基团所组成。视蛋白的肽链序列已搞清,它的肽链中有7段穿越所在膜结构、主要由疏水性氨基酸组成的α-螺旋区段,同一般的细胞膜受体具有类似的结构。视黄醛由维生素A变来,后者是一种不饱和醇,在体内一种酶的作用下可氧化成视黄醛。提纯的视紫红质在溶液中对500nm波长的光线吸收能力最强,这与人眼在弱光条件下对光就业上蓝绿光区域(相当于500nm波长附近)感觉最明亮(不是感到了蓝绿色)的事实相一致(图9-7),说明人在暗视觉与视杆细胞中所含视紫红质的光化学反应有直接的关系。
图9-7弱光条件下人眼所感到的光谱亮度曲线和实验条件下
视紫红质对光谱不同部分的吸收曲线视觉中最明亮的区域和视紫红制裁
吸收能力最强的部分都在500nm的波长附近
视紫红质在光照时迅速分解为视蛋白和视黄醛,这是一个多阶段的反应。目前认为,分解的出现首先是由于视黄醛分子在光照时发生了分子构象的改变,即它在视紫红质分子中本来呈11-顺型(一种较为弯曲的构象),但在光照时变为全反型(一种较为直的分子构象)。视黄醛分子构象的这种改变,将导致视蛋白分子构象也发生改变,经过较复杂的信号传递系统的活动,诱发视杆细胞出现感受器电位。据计算,一个光量子被视紫红质吸收,就足以使视黄醛分子结构发生改变,导致视紫红质最后分解为视蛋白和视黄醛。视紫红质分解的某些阶段伴有能量的释放,但这看来不是诱发感受器电位的直接原因。
在亮处分解的视紫红质,在暗处又可重新合成,亦即它是一个可逆反应,其反应的平衡点决定于光照的强度。视紫红质再合成的第一步,是全反型的视黄醛变为11-顺型的视黄醛,很快再同视蛋白结合。此外,贮存在视网膜的色素细胞层中的维生素A也是全反型的,它们也可在耗能的情况下变成11-顺型的,进入视杆细胞,然后再氧化成11-顺型的视黄醛,参与视紫红质的合成补充;但这个过程进行的速度较慢,不是促进视紫红制裁再合成的即时因素。人在暗处视物时,实际是既有视紫红质的分解,又有它的合成,这是人在暗光处能不断视物的基础;光线愈暗,全盛过程愈超过分解过程,视网膜中处于合成状态的视紫红质数量也愈高,这也使视网膜对弱光愈敏感;相反,人在亮光处时,视紫红质的分解增强,合成过程甚弱,这就使视网膜中有较多的视紫红质处于分解状态,使视杆细胞几乎失去了感受光刺激的能力;事实上,人的视觉在亮光处是靠另一种对光刺激较不敏感的感光系统即视锥来完成的,后一系统在弱光时不足以被刺激,而在强光系统下视杆细胞中的视紫红质较多地处于分解状态时,视锥系统就代之而成为强光刺激的感受系统。在视紫红质和再合成的过程中,有一部分视黄醛被消耗,这最终要靠由食物进入血液循环(相当部分贮存于肝)中的维生素A来补充。长期摄入维生素A不足,将会影响人在暗光处的视力,引起夜盲症。
(二)视杆细胞外段的超威结构和感受器电位的产生
感光细胞的外段是进行光-电转换的关键部位。视杆细胞外段具有特殊的超威结构,如图9-8所示。在外段部分,膜内的细胞浆甚少,绝大部分为一些整齐的重叠成层的圆盘状结构所占据,这圆盘称为视盘。每一个视盘是一个扁平的囊状物,囊膜的结构和细胞膜类似,具有一般的脂质双分子层结构,但其中镶嵌着的蛋白质绝大部分是视紫红质,亦即视杆细胞所含的视紫红质实际上几乎全部集中在视盘膜中。视盘的数目在不同动物的视杆细胞中相差很大,人的每个视杆细胞外段中它们的数目近千;每一个视盘所含的视紫红质分子约有100万个。这样的结构显然有利于使进入视网膜的光量子有更大的机会在外段中碰到视紫红质分子。
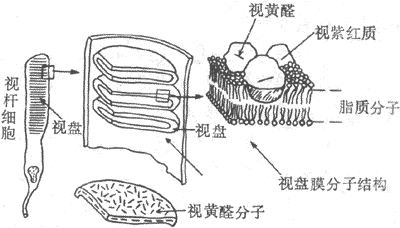
图9-8 视杆细胞外段的超微结构示意图
有人用细胞内微电极技术,研究了视杆细胞外段内外的电位差在光照前后的变化,结果发现在视网膜未经照射时,视杆细胞的静息电位只有-30?/FONT>-40mV,比一般细胞小得多。经分析表明,这是由于外段膜在无光照时,就有相当数量的Na+通道处于开放状态并有持续的Na+内流所造成,而内段膜有Na+泵的连续活动将Na+移出膜外,这样就维持了膜内外的Na+平衡。当视网膜受到光照时,可看到外段膜两侧电位短暂地向超极化的方向变化,由此可见,外段膜同一般的细胞膜不一致,它是在暗处或无光照时处于去极化状态,而在受到光刺激时,跨膜电痊反而向超极化方向变化,因此视杆细胞的感受器电位(视锥细胞也一样),表现为一种超极化型的慢电位,这在所有被研究过的发生器或感受器电位中是特殊的,它们一般都表现为膜的暂时去极化。
光子的吸收引起外段膜出现超极化电反应的机制已基本搞清,这就是光量子被作为受体的视紫红质吸收后引起视蛋白分子的变构,又激海参了视盘膜中一种称为传递蛋白(transducin)Ct的中介物,后者在结构上属于G-蛋白家庭的一员,它激活的结果是进而激活附近的磷酸二酯酶,于是使外段部分胞浆中的cGMP大量分解,而胞浆中cGMP的分解,就使未受光刺激时结合于外段膜的cGMP由也膜解离而被分解,而cGMP在膜上的存在正是这膜中存在的化学门控式Na+通道开放的条件,膜上cGMP减少,Na+通道开放减少,于是光照的结果出现了我们记录到的超极化型感受器电位。据估计,一个视紫红质被激活时,可使约500个传递蛋白被激活;虽然传递蛋白激活磷酸二酯酶是1对1的,但一个激活了的磷酸二酯酶在一秒钟内大约可使4千多个cGMP分子降解。由于酶系统的这种生物放大作用,就可以说明1个光量子的作用何以能在外段膜上引起大量化学门控式Na+通道的关闭,引起一个足以为人的视觉系统所感知的超极化型电变化。
视杆细胞外段和整个视杆细胞都没有产生动作电位的能力,由光刺激在外段膜上引起的感受器电位只能以电紧张性的扩布到达它的终足部分,影响终点(相当于轴突末稍)外的递质释放。
五、视锥系统的换能和颜色视觉
视锥系统外段也具有与视杆细胞类似的盘状结构,并含有特殊的感光色素,但分子数目较少。已知,大多数脊椎动物具有三种不同的视锥色素,各存在于不同的视锥细胞中。三种视锥色素都含有同样的11-顺型视黄醛,只是视蛋白的分子结构稍有不同。看来是视蛋白分子结构中的微小差异,决定了同它结合在一起的视黄醛分子对何种波长的光线最为敏感,因而才有视杆细胞中的视紫红质和三种不同的视锥色素的区别。光线作用于视锥细胞外段时,在它们的外段膜两侧也发生现视杆细胞类似的超级化型感受器电位,作为光-电转换的第一步。目前认为视锥细胞外段的换能机制,也与视杆细胞类似。
视锥细胞功能的重要特点,是它有辨别颜色的能力。颜色视觉是一种复杂的物理-心理现象,颜色的不同,主要是不同波长的光线作用于视网膜后在人脑引起的主观印象。人眼一般可在光谱上区分出红、橙、黄、绿、青、蓝、紫等七种颜色,每种颜色都与一定波长的光线相对应;但仔细的检查可以发现,单是人眼在光谱可区分的色泽实际不下150种,说明在可见光谱的范围内波长长度只要有3-5nm的增减,就可被视觉系统分辨为不同的颜色。很明显,设想在视网膜中存在上百种对不同波长的光线起反应的视锥细胞或感光色素,是不大可能的。但物理学上从牛顿的时代或更早就知道,一种颜色不仅可能由某一固定波长的光线所引起,而且可以由两种或更多种其他波长光线的混合作用而引起。例如,把光谱上的七色光在所谓牛顿色盘上旋转,可以在人眼引起白色的感觉;用红、绿、蓝三种色光(不是这三种颜色的颜料)作适当混合,可以引起光谱上所有任何颜色的感觉。这后一现象特别重要;这种所谓三原色混合原理不仅早已广泛地应用于彩色照像、彩色电视等方面,而且被用于说明颜色视觉的产生原理本身。早在上世纪初,Young(1809)和Helmholtz(1824)就提出了视觉的三原色学说,设想在视网膜中存在着分别对红、红、蓝的光线特别敏感的三种视锥细胞或相应的三种感光色素,并且设想当光谱上波和介于这三者之间的光线作用于视网膜时,这些光线可对敏感波长与之相近两种视锥细胞或感光色素起不同程度的刺激作用,于是在中枢引起介于此二原色之间的其他颜色的感觉。视觉三原色学说用较简单的生物感受结构的假设说明了复杂的色觉现象,一般为多数人所接受;但在实验中试图寻找出游同种类的视锥细胞或感光色素长时间未获成功。用光学显微镜和电子显微镜不能发现视锥细胞之间在结构上有什么不同,同时也未能用一般的化学方法分离邮不同的视锥感光色素。
70年代以来,由于实验技术的进步,关于视网膜中有三种对不同波长光线特别敏感的视锥细胞的假说,已经被许多出色的实验所证实,例如,有人用不超过单个视锥直径的细小单色光束,逐个检查并绘制在体(最初实验是在金公和蝾螈等动物进行,以后是人)视锥细胞的光谱吸收曲线,发现所有绘制出来的曲线不外三种类型,分别代表了三类光谱吸收特性不同的视锥细胞,一类的吸收峰值在420nm外,一类在531nm外,一类在558nm外,差不多正好相当于蓝、绿、红三色光的波长,和上述视觉三原色学说的假设相符。用微电极记录单个视锥细胞感受器电位的方法,也得到了类似的结果,即不同单分光引起的超极化型感受器电位的大小,在不同视锥细胞是不一样的(图9-9),峰值出现的情况符合于三原色学说。
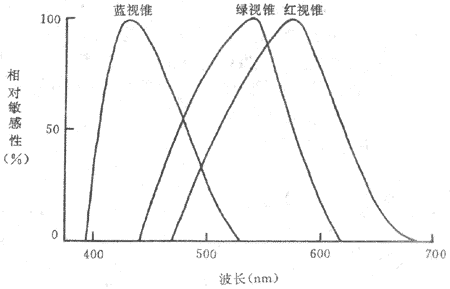
图9-9 人视网膜中三种不同视锥细胞的光谱相对敏感性
三原色学说和它的实验依据,大体上可以说明临床上遇到的所谓色盲和色弱的可能发病机制。红色盲也称第一色盲,被认为是由于缺乏对较长波长光线敏感的视锥细胞所致;此外还有绿色盲,也称第二色盲,蓝色盲也称第三原色盲,都可能是由于缺乏相应的特殊视锥细胞所致。红色盲和绿色盲较为多见,在临床上都不加以区别地称为红绿色盲;蓝色盲则极少见。色盲患者的颜色不仅不能识别绿色,也不能区分红也绿之间、绿与蓝之间的颜色等。有些色觉异常的人,只是对某种颜色的识别能力差一些,亦即他们不是由于缺乏某种视锥细胞,而只是后者的反应能力较正常人为弱的结果,这种情况有别于真正的色盲,称为色弱。色盲除了极少数可以由于视网膜后天病变引起外,绝大多数是由遗传因素决定的。
三原色学说虽然比较圆满地说明许多色觉现象和色盲产生的原因,并已在光感受细胞的一级得到了实验证实,但并不能解释所有的颜色视觉现象,如颜色对比现象就是一个例子。试将蓝色的小纸块放在黄色或其它颜色的背景上,会觉得放在黄色背景上那个蓝纸块特别蓝,同时觉得背景也比未放蓝纸块时更黄(在我国北方的黄土高原,当春天的风造成黄尘蔽日的情况时,会觉得平常的日晃灯管的光线变得较蓝了)。这种现象称为颜色对比,而黄和蓝则称为对比色或互补色。颜色对比现象只出现对比色之间,而不是任意的两种颜色之间。互为对比色的颜色对尚有:红一绿以及黑和白。根据颜色对比等不容易用三原色学说圆满视觉现象,几乎是在三原色学说提出的同时就出现了另一种色觉学说,称为对比色学说(Hering,1876)。该学说提出在视网膜中存在着三种物质,各对一组对比色的刺激起性质相反的反应。如前所述,近年来在视锥细胞一级进行的研究有利于三原色学说而不利于对比色学说,但后来在视网膜其它层细胞进行的一些实验却又符合对比色学说的推测。如在金鱼水平细胞进行的微电极研究说明,此类细胞和视杆、视锥细胞不同,既能出现超极化的跨膜电位改变,也能出现去极化型的电位改变,而且在用多种不同色光刺激时发现,有些水平细胞在黄光刺激时出现最大的去极化反应,在蓝光刺激时出出现最大的超极化型反应;另一些水平细胞则在红和绿色刺激时有类似的不同反应。这些现象是同对比色学说一致的。看来可能的是,各以部分色觉现象为出发点的两种色觉学说都是部分正确的,在视锥细胞一级,不同色光以引起三种不同视锥细胞产生不同大小的超极化型电变化进行编码;但到了水平细胞一级或其它级细胞(包括某些中枢神经元),信息又进行重新编码,不同颜色双可以用同细胞对互为对比色的颜色出现相反形式的电反应来编码。以上事实说明,颜色视觉的引起是一个十分复杂的过程,它需要有从视网膜视锥细胞到皮层神经元的多级神经成分的参与才能完成。
六、视网膜的信息处理
由视杆和视锥细胞产生的电信号,在视网膜内要经过复杂的细胞网络的传递,最后才能由神经节细胞发生的神经纤维以动作电位的形式传向中枢。由于视网膜内各种细胞之间的排列和联系非常复杂,与细胞间信息传递有关的化学物质种类繁多(除一般神经系统中常见的递质外,连同视网膜中已发现的各神经肽类物质,总数已达30余种),因而视觉信息在从感光细胞向节细胞传递时,必然要经历种种改变;这实际就是视网膜本身对视觉信息的初步处理,它是在视网膜特定的细胞构筑和化学构筑的网络中按照某些规律进行的,但对这些规律的了解还是很肤浅的。现在所能初步肯定的是,双极细胞、水平细胞和多数无长突细胞也同两种感光细胞一样,没有产生动作电位的能力(但部分无长突细胞可产生动作电位);但这三种细胞同感光细胞不同的是,它们在前一级细胞的影响之下,既能产生超级化型慢电位,也能产生去极化型慢电位(相当于一般神经元突触后膜处的IPSp 和EPSP,见第十章)。所有这些慢电位,只能作电紧张性的扩布(参看第二章局部兴奋),影响突触前膜递质释放量的改变,从而引起下一级细胞产生慢电位变化(也包括电突触性相互影响);只有当这样的慢电位传递到神经节细胞体时,由于后者有产生动作电位的能力,当两种形式的慢电位总和的结果,使节细胞的静息膜电位能够去极经到阈电位水平时,才会产生“全或无”式的动作电位,作为视网膜的最后输出信号传向中枢(图9-10)。
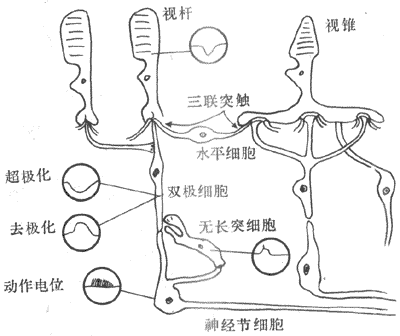
图9-10视网膜各类细胞排列及其产生的电反应的类型示意图
只有神经节细胞能产生动作电位
虽然关于视网膜中信息处理的细节还不很清楚,但不妨先把视网膜作为控制论中的“黑箱”来看待,只对它的输入(相当于作用于视网膜的光刺激)和它的输入(相当于由视神经传出的动作电位序列)进行对照分析,这样也可以初步理解视网膜信息处理和编码的最终结果。首先要记信一个事实是,视神经中纤维的总数(亦即节细胞的总数),只有全部感光细胞的1%。这一简单事实就足以说明,视神经不可能通过其纤维“点对点”地传递视网膜中各感光细胞被光照的情况(中央凹处少数视锥细胞例外);因而大多数视神经纤维所传递的信号,只能是决定于多个感光细胞并因而含较多的信息量。
用小的光点刺激猫视网膜并同时记录单条视神经纤维动作电位的方法表明,发出视神经纤维的节细胞大致可分为三类,分别称为X-、Y-和W-细胞。X-和Y-细胞的特点是它们都具有大致呈同心圆式的“中心-周边感受野”;一个节细胞的感受野在这里指视网膜上某一特定的区域,当后者受到刺激时可能使该节细胞发生反应;但这两种细胞的感受野都由两部分构成;当光线作用于感受野的中心部分时出现节细胞放电增加,而当光线作用于环绕该中心的一定范围内视网膜部分时,反而使该节细胞放电减少(图9-11,上),这是一种类型的中心-周边感受野,这就是当感受野的中心部分接受光刺激时引起节细胞放电数减少,而刺激其周边部分时,反而引起该细胞放电增多,这种节细胞可称为中心撤光反应细胞(图9-11,下)。
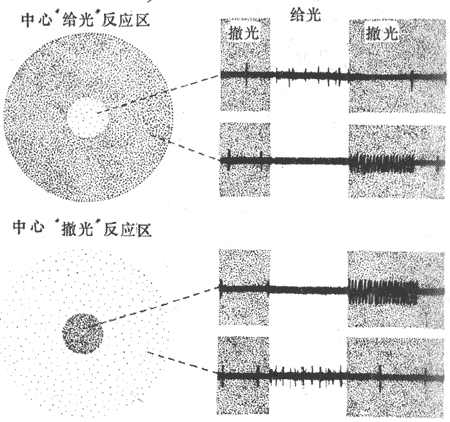
图9-11 视神经纤维的感受野和放电特征
X-和Y-细胞都可以有上述两种类型的感受野。它们之间的区别是,X-细胞的感受野较小,对刺激的反应是持续的;Y-细胞的感受野较大,而反应是时相性的,与刺激呈非线性关系。在猴子的实验中还发现,不同的X-细胞对不同波长的光线反应不同,但对光的强度变化不敏感;而Y-细胞则对光照的强度变化敏感,却对光线的波长变化不敏感。至于W-细胞,它们有较前两者大得多的感受野,或是在光刺激时有放电,或是在撤光时有放电,但对刺激的反应比较迟缓,一般没有性质相反的周边视野区域。从形态学上看,X-和X-细胞主要由双极细胞接受输入,而W细胞则主要接受无长突细胞的传入。
上述简单的事实说明,每一条视神经纤维上的神经冲动并不简单地表示某一部分视网膜受到光照或无光照,以上述X-为例,它的最强烈放电是出现在其感受野中心部分受到光照、而其外周部分全无光照(或相反的情况)的时候;如果中心和外周部同时受到光照,此节细胞的放电反而无大变化或只少有增加。另外,有事实说明,视网膜像经视网膜处理后,已被分解为不同“像素”,如有节细胞贡中枢传输图像组成波长的住处有的传输它的不同亮度。这种把感受信息分解为其组成的“要素”,再在感受通路中进行“平行”传输和处理,在脑的各种感觉功能的研究中经常遇到,但是人的视觉也和另的感觉一样,最复杂的信息处理和加工发生在中枢、特别是它的高级部分。
七、与视觉有关的其它现象
(一)暗适应和明适应
人从亮处进入暗室时,最初看不清楚任何东西,经过一定时间,视觉敏感度才逐渐增主,恢复了在暗处的视力,这称为暗适应。相反,从暗处初来到亮光处,最初感到一片耀眼的光亮,不能看清物体,只有稍待片刻才能恢复视觉,这称为明适应。
暗适应是人眼对光的敏感度在暗光处逐渐提高的过程。在进入暗室后的不同时间,连续测定人的视觉阈值,亦即测定人眼刚能感知的光刺激强度,可以看到此阈逐渐变小、亦即视觉的敏感度在暗处逐渐提高的过程。如图9-12所示,一般是在进入暗室后的最初约7分钟内,有一个阈值的明显下降期,以后又出现阈值的量明显下降;于进入暗室后的大约25-30分钟时,阈值下降到最低点,并稳定于这一状态。暗适应的产生机制与视网膜中感光色素在暗处时再合成增加,因而增加了视网膜中处于未分解状态的色素的量有关。据分析,暗适应的第一阶段主要与视锥细胞色素的合成量增加相一致;第二阶段亦即暗适应的主要构成部分,则与视杆细胞中视紫红质的合成增强有关。
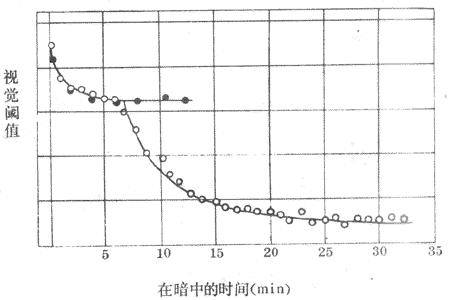
图9-12 暗适应曲线
。表示用白光对全眼的测定结果
·表示用红光对中央凹测定的结果(表示视锥细胞单独的暗适应曲线,
因中央凹为视锥细胞集中部位,且红光不易被视杆细胞所感受)
明适应出现较快,约需一分钟即可完成。耀眼的光感主要是由于在暗处蓄积起来的合成状态的视紫红质在进入亮处时先迅速分解,因为它对光的敏感性较视锥细胞中的感光色素为高;只有在较多的视杆细胞色素迅速分解之后,对光较不敏感的视锥细胞色素才能在亮光环境中感光。
(二)视野
单眼固定地注视前方一点不动,这时该眼所能看到的范围称为视野。视野的最大界限应以它和视轴(单眼注视外界某一点时,此点的像正好在视网膜黄斑中央凹处,连接这两点的假想线即视轴)所成夹角的大小来表示。在同一光照条件下,用不同颜色的目标物测得的视野大小不一样,白色视野最大,其次为黄蓝色,再次为红色,而以绿色视野为最小。设想视野的大小除与各类感光细胞在视网膜中的分布范围有关外,在细节上还缺乏圆满的解释。另外,由于面部结构阻挡视线,也影响视野的形状,如一般人颞侧视野较大,鼻侧视野较小等。临床医生检查视野,使用特制的视野仪,并用不同颜色的视标进行检查,目的在于了解视网膜的普遍感光能力,有时可藉以发现较大范围的视网膜病变。某些视网膜、视神经或视觉传导路的病变,有特殊形式的视野缺损,在诊断有意义。
(三)视网膜电图
将一个引导电极和角膜接触,另一个电极置于额部作为参考电极,当给视网膜以广泛光刺激时,可以在灵敏的电测量仪器上记录到一系列电变化,这称为视网膜电图(electroretinogram)。视网膜电图在性质上不同于同微电极在单一视网膜细胞成分记录到的电现象;视网膜电图是整个视网膜中各种成分在受到大范围光照时的多种电反应的综合反映。它通常由命名为a、b和c的三个波组成(图9-13)。据实验分析,a波主要来源于感光细胞的感受器电位;b波幅度较大,主要与双极细胞等细胞的活动有关;c波平缓而持续时间长,可能与色素细胞层的活动有关。有时在光照撤除时还可在缓慢持续的c波上再出现一个波动,称为d波,产生原因尚不明了。视网膜电图虽易于测量和描记,但反映视网膜功能状态或病变的特异性不大,目前只发现少数疾病时有特殊的视网膜电图改变,故在临床上作用的意义不很大。
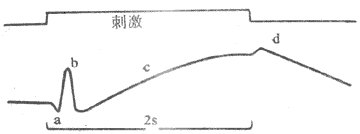
图9-13 猫的视网膜电图上面直线上的标记是给光和撤光时间
(四)双眼视觉和立体视觉
人和高等哺乳动物的双眼都在面部前方,视物时两眼视野差不多部分的像又各循自己特有的神经通路传向中枢,但正常人主观察感觉上只产生一个“物”的感觉。两眼视物而只产生一个视觉形象的前提条件是:由物质同一部分的的光线,应成像在两侧视网膜的相称点上。例如,两眼的黄斑部就互为相称点;当两眼注视培墙上一个小黑点时,由于有眼外肌的调节,此点就都正好成像在两侧眼的黄斑上,于是在视觉中只“看到”一个点;此时如用手轻推一侧眼球的外侧,使此眼视轴稍作偏移,则这时此眼视网膜上的黑点像就要从黄斑部移开,落在与对侧视网膜像非相称的点上,于是会感到墙上有两个黑点存在,这就是复视现象。显然,在黄斑部以外,一眼的颞侧视网膜和另一眼的鼻侧视网膜互相对称;而一眼的鼻侧视网膜也与另正好的颞侧视网膜互相对称。
第三节 听觉器官
听觉的外周感受器官是耳,耳的适宜刺激是一定频率范围内的声波振动。耳由外耳、中耳和内耳迷路中的耳蜗部分组成。由声源振动引起空气产生疏密波,后者通过外耳道、鼓膜和听骨链的传递,引起耳蜗中淋巴液和基底膜的振动,使耳蜗科蒂器官中的毛细胞产生兴奋。科蒂器官和其中所含的毛细胞,是真正的声音感受装置,外耳和中耳等结构只是辅助振动波到达耳蜗的传音装置。听神经纤维就分布在毛细胞下方的基底膜中;振动波的机械能在这里转变为听神经纤维上的神经冲动。并以神经冲动的不同频率和组合形式对声音信息进入编码,传送到大脑皮层听觉中构,产生听觉。听觉对动物适应环境和人类认识自然有重要的意义;在人类,有声语言是互
通信息交流思想的重要工具。
因此,在耳的生理功能研究中主要解决的问题是:声音怎样通过外耳、中耳等传音装置传到耳蜗,以及耳蜗的感音装置如何把耳蜗淋巴液和基底膜的振动转变成为神经冲动。
一、人耳的听阈和听域
耳的适宜刺激是空气振动的疏密波,但振动的频率必须在一定的范围内,并且达到一定强度,才能被耳蜗所感受,引起听觉。通常人耳能感受的振动频率在16-20000Hz之间,而且对于其中每一种频率,都有一个刚好能引起听觉的最小振动强度,称为听阈。当振动强度在吸阈以上继续增加时,听觉的感受也相应增强,但当振动强度增加到某一限度时,它引起的将不单是听觉,同时还会引起鼓膜的疼痛感觉,这个限度称为最大可听阈。由于对每一个振动频率都有自己的听阈和最大或听阈,因而就能绘制出表示人耳对振动频率和强度的感受范围的坐标图,如图9-14所示。其中下方曲线表示不同频率振动的听阈,上方曲线表示它们的最大听阈,两得所包含的面积则称为听域。凡是人所能感受的声音,它的频率和强度的坐标都应在听域的范围之内。由听域图可看出,人耳最敏感的频率在1000-3000Hz之间;而日常语言的频率较此略低,语音的强度则在听阈和最大可听阈之间的中等强度处。
二、外耳和中耳的传音作用
(一)耳廓和外耳道的集音作用和共鸣腔作用
外耳由耳廓和外耳道组成。人耳耳廓的运动能力已经退化,但前方和侧方来的声音可直接进入外耳道,且耳廓的形状有利于声波能量的聚集,引起较强的鼓膜振动;同样的声音如来自耳廓后方,则可被耳廓遮挡,音感较弱。因此,稍稍转动头的位置,根据这时两耳声音强弱的轻微变化,可以判断音源的位置。
外耳首是声波传导的通路,一端开口,一端终止于鼓膜。根据物理学原理,充气的管道可与波长4倍管长的声波产生最大的共振作用;外耳道长约2.5cm,据此计算,它作为一个共鸣腔的最佳共振频率约在3500Hz附近;这样的声音由外耳道传到鼓膜时,其强度可以增强10倍。
(二)鼓膜和中耳听骨链增压效应
中耳包括鼓膜、鼓室、听骨链、中耳小肌和咽鼓管等主要结构,其中鼓膜、听骨链和内耳卵圆窗之间的关系如图9-15所示,它们构成了声音由外耳传向耳蜗的最有效通路。声波在到达鼓膜交,由空气为振动介质;由鼓膜经听骨链到达卵圆窗膜时,振动介质变为固相的生物组织。由于不同介质的声阻拦不同,理论上当振动在这些介质之间传递时,能量衰减极大,估计可达99%或更多。但由于由鼓膜到卵圆窗膜之间的传递系统的特殊力学特性,振动经中耳传递时发生了增压效应,补偿了由声阻挡不同造成的能量耗损。
鼓膜呈椭圆形,面积约50-90mm2,厚度约0.1mm。它不是一个平面膜,呈顶点朝向中耳的漏斗形。其内侧连锤骨柄,后者位于鼓膜的纤维层和粘膜层之间,自前上方向下,终止于鼓膜中心处。鼓膜很像电话机受话器中的振膜,是一个压力承受装置,具有较好的频率响应和较小的失真度,而且它的形状有利于把振动传递给位于漏斗尖顶处的锤骨柄。据观察,当频率在2400Hz以下的声波作用于鼓膜时,鼓膜都可以复制外加振动的频率,而且鼓膜的振动与声波振动同始同终,很少残余振动。
图9-15 人中耳和耳蜗关系模式图
点线表示鼓膜向内侧振动时各有关结构的移动情况
听骨链由锤骨、砧骨及镫骨依次连接而成。锤骨柄附着于鼓膜,镫骨脚板和卵圆窗膜相接,砧骨居中,将锤骨和镫骨连接起来,使三块听小骨形成一个两壁之间呈固定角度的杠杆。锤骨柄为长臂,砧骨长突为短臂。该械杆系统的特点是支点刚好在整个听骨链的重心上,因而在能量传递过程中惰性最小,效率最高。鼓膜振动时,如锤骨柄内移,则砧骨的长突和镫骨亦和锤骨柄作同方向的内移,如图9-15中点线所示。
中耳增压泖应主要有以下两个因素:一是由于鼓膜面积和卵圆窗膜的面积大小有差别,鼓膜振动时,实际发生振动的面积约55mm2,而卵圆窗膜的面积只有3.2mm2,如果听骨链传递时总压力不变,则作用于卵圆窗膜上的压强将增大55÷3.2=17倍;二是听骨链中杠杆长臂和短臂之比约为1.3:1,即锤骨柄较长,于是短臂一侧的压力将增大为原来的1.3倍。这样算来,整个中耳传递过程的增压效应为17×1.3=22倍。
与中耳传音功能有关的,还有中耳内的两条小肌肉,其中鼓膜张肌收缩时,可使锤骨柄和鼓膜内向牵引,增加鼓膜紧张度;镫骨肌收缩时,使镫骨脚板向外后方移动。强烈的声响气流经过外耳道,以及角膜和鼻粘膜受到机械刺激时,都可以反射性地引起这两块小肌肉的收缩,其结果是使鼓膜紧张,使各听小骨之间的边境更为紧张,导致吸骨链传递振动的幅度减小;阻力加大,总的效果是使中耳的传音效能有所减弱。据认为,这一反应可以阻止较强的振动传到耳蜗,对感音装置起到某种保护作用;但由于声音引起中耳肌的反射性收缩需经过十几个毫秒的潜伏期,故它们对突然发生的短暂爆炸声的保护作用不大。
(三)咽鼓管的功能
咽鼓管亦称耳咽管,它连通鼓室和鼻咽部,这就使鼓室内空气和大气相通,因而通过咽鼓管,可以平衡鼓室内空气和大气压之间有可能出现的压力差,这对于维持鼓膜的正常位置、形状和振动性能有重要意义。咽鼓管阻塞时,鼓室气体将被吸收,使鼓室内压力下降,引起鼓膜内陷。暂时的鼓膜内外压力差,常发生在外耳道内压力首先发生改变而鼓室内压力仍处于原初的状态,如飞机的突然升降长潜水等,此时如果不能通过咽鼓管使鼓室内压力外耳道压力(或大气压)取得平衡,就会在鼓膜两侧出现巨大的压力差。据观察,这个压力差如达到9.33-10.76kPa(70-80mmHg),将会引起鼓膜强烈痛疼;压力差超过24kPa(180mmHg)时,可能造成鼓膜破裂。咽鼓管在正常情况下其鼻咽部开口常处于闭合状态,在吞咽、打呵欠或喷嚏时由于腭帆张肌等肌肉的收缩,可使管口暂时开放,有利于气压平衡。
声音的骨传导 正常时听觉的引起,是由于声波经外耳道引起鼓膜的振动,再经听骨链和卵圆窗膜进入耳蜗,这一条声音传递地途径,称为气传导。此外,声波还可以直接引起颅骨的振动,再引起位于颞骨骨质中的耳蜗内淋巴的振动,这称为骨传导。骨传导正常时较气传导不敏感得多,几乎不能感到它的存在;能察知骨传导存在的一种方面是,把一个振动阒的音叉的柄直接和颇骨接触,这时人会感到一个稍有异样的声音;当这个声音减弱到听不到以后,再把音叉迅速移到耳廓前方,这时又能听到声音的存在。这个简单实验说明骨传导的存在,也说明正常时气传导较骨传导为灵敏。可以认为,骨传导在正常听觉的引起中作用微乎其微。不过临床上常通过检查患者气传导和骨传导受损的情况,判断听觉异常的产生部位和原因。
三、耳蜗的感音换能作用
耳蜗的作用是把传到耳蜗的机械振动转变成听神经纤维的神经冲动。在这一转变过程中,耳蜗基底膜的振动是一个关键因素。它的振动使位于它上面的毛细胞受到刺激,引起耳蜗内发生各种过渡性的电变化,最后引起位于毛细胞底部的传入神经纤维产生动作电位。
(一)耳蜗的结构要点
耳蜗是一条骨质的管道围绕一个骨轴盘旋21/2-23/4周而成。在耳蜗管的横断面上可见到两个分界膜,一为斜行的前庭膜,一为横行的基底膜,此两膜将管道分为三个腔,分别称为前庭阶、鼓阶和蜗管(图9-16)。前庭附在耳蜗底部与卵圆窗膜相接,内充外淋巴;鼓阶在耳蜗底部与圆窗膜相接,也充满外淋巴,后者在耳蜗顶部和前庭阶中的外淋巴相交通;蜗管是一个盲管,其中内淋巴浸浴着位于基底膜上的螺旋器的表面。螺旋器的构造极为复杂;在蜗管的横断面上的靠蜗轴一侧,可看到有一行内毛细胞纵向排列;在蜗管的靠外一侧,有3-5行外毛细胞纵向排列(参看图9-18);此外还有其他的支持细胞和存在于这些细胞间的较大的间隙,包括内、外隧道和Nuel间隙。需要指出的是,这些间隙中的液体在成分上和外淋巴一致,它们和蜗管中的内淋巴不相交通,但可通过基底膜上的小孔与鼓阶中的外淋巴相交通。这样的结构使得毛细胞的顶部与蜗管中的内淋巴相接触,而毛细胞的周围和底部则和外淋巴相接触。每一个毛细胞的项部表面,都有上百条排列整齐的听毛,其中较长的一些埋植在盖膜的冰胶状物质中,有些则只和盖膜接触。盖膜在内侧连耳蜗轴,外侧游离在内淋巴中。
(二)基底膜的振动和行波理论
当声流振动通过听骨链到达卵圆窗膜时,压力变化立即传给隔离蜗内液体和膜性结构;如果卵圆窗膜内移,前庭膜和基底膜也将下移,最后是鼓阶的外淋巴压迫圆窗膜外移;相反,当卵圆窗膜外移时,整个耳蜗内结构又作反方向的移动,于是形成振动。可以看出,在正常气传导的过程中,圆窗膜实际起着缓冲耳蜗内压力变化的作用,是耳蜗内结构发生振动的必要条件。有人用直接观察的方法,详细记录了声音刺激引起的基底膜振动的情况,这对于了解基底膜振动的形式,以及这种振动在耳蜗接受不同频率的声音刺激时有何差异,提供了可靠的依据。观察表明,基底膜的振动是以行波(traveling wave)的方式进行的,即内淋巴的振动首先是靠近卵圆窗处引起基底膜的振动,此波动再以行波的形式沿基底膜向耳蜗的顶部方向传播,就像人在抖动一条绸带时,有行波沿绸带向远端传播一样。下一步还证明,不同频率的声音引起的行波都从基底膜的底部,即靠近卵圆窗膜处开始,但频率不同时,行波传播的远近和最大行波的出现部位有气温同,如图9-17所示,;这就是振动频率愈低,行波传播愈远,最大行波振幅出现的部位愈靠近基底膜顶部,而且在行波最大振幅出现后,行波很快消失,不再传播;相反地,高频率声音引起的基底膜振动,只局限于卵圆窗附近。
图9-16 耳蜗管的横断面图
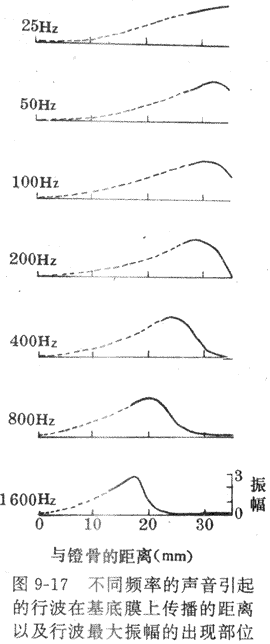
不同频率的振动引起的基底膜不同形式的行波传播,主要由基底膜的某些物理性质决定的。基底膜的长度在人约为30mm,较耳蜗略短,但宽度在靠近卵圆窗处只有0.04mm,以且逐渐加宽;与此相对应,基底膜上的蚴旋器的高度和重量,也随着基底膜的加宽而变大。这些因素决定了基底膜愈靠近底部,共振频率愈高,愈靠近顶部,共振频率愈低;这就使得低频振动引起的行波在向顶部传播时阻力较小,而高频振动引起的行波只限局在底部附近。
不同频率的声音引起的不同形式的基底膜的振动,被认为是耳蜗能区分不同声音频率的基础。破坏动物不同部位基底膜的实验和临床上不同性质耳聋原因的研究,都证明了这一结论,亦即耳蜗底部受时主要影响高频听力,耳蜗顶部受损时主要影响低频听力。不能理解,既然每一种振动频率在基底膜上都有一个特定的行波传播范围和最大振幅区,与这些区域有关的毛细胞和听神经纤维就会受到最大的刺激,这样,来自基底膜不同区域的听神经纤维的神经冲动及其组合形式,传到听觉中枢的不同部位,就可能引起不同音调的感觉。
基底膜的振动怎样使毛细胞受到刺激,如图9-18所示。毛细胞顶端的听毛有些埋在盖膜的胶状物中,有些是和盖膜的下面接触;因盖膜和基底膜的振动轴不一致,于是两膜之间有一个横向的交错移动,使听毛受到一个切向力的作用而弯曲(图9-18,下)。据研究,毛细胞听纤毛的弯曲,是耳蜗中由机械能转为电变化的第一步。
图9-18基底膜和盖膜振动时毛细胞顶部听毛受力情况
上:静止时的情况 下:基底膜在振动中上移时,因与盖膜之间的切向运动,听毛弯向蜗管外侧
(三)耳蜗的生物现象
在耳蜗结构中除了能记录到与听神经纤维兴奋有关的动作电位,还能记录到一些其他形式的电变化。在耳蜗未受到刺激时,如果把一个电极放在鼓阶外淋巴中,并接地使之保持在零电位,那么用另一个测量电极可测出蜗管内淋巴中的电位为+80mV左右,这称为内淋巴电位。如果将此测量电极刺入毛细胞膜内,则膜内电位为-70?/FONT>-80mV。毛细胞顶端膜外的浸浴液为内淋巴,则该处毛细胞内(相当于-80mV)和膜外(相当于+80mV)的电位差当为160mV;而在毛细胞周围的浸浴液为外淋巴(电位相当于零),该处膜内外的电位差只有80mV左右;这是毛细胞静息电位和一般细胞不同之处。据实验分析,内淋巴中正电位的产生和维持,同蜗管外侧壁处的血管纹结构的细胞活动有直接关系(图9-16),并且对缺 O2非常敏感;有人发现,血管纹细胞的膜含有大量活性很高的ATP酶,具有“钠泵”的作用,它们可依靠分解ATP获得能量,将血浆中的K+泵入内淋巴,将内淋巴中的Na+泵入血浆,但被转运的K+担超过了Na+的量,这就使内淋巴中有大量K+蓄积,因而使内淋巴保持了较高的正电位;缺O2使ATP的生成受阻,也使Na+泵的活动受阻,因而使内淋巴的正电位不能维持。
当耳蜗接受声音刺激时,在耳蜗及其附近结构又可记录到一种特殊的电波动,称为微音器电位。这是一种交流性质的电变化,在一定的刺激强度范围内,它的频率和幅度与声波振动完全一致(图9-19);这一现象正如向一个电话机的受话器或微音器(即麦克风)发声时,它们可将声音振动转变为波形类似的音频电信号一样,这正是把耳蜗的这种电变化称为微音器电位的原因。事实上,如果对着一个实验动物和耳廓讲话,同时在耳蜗引导它的微音器电位,并将此电位经放大后连接到一个扬声器,那么扬声器发出的声音正好是讲话的声音!这一实验生动地说明,耳蜗在这里起着类似微音器的作用,能把声波变成相应的音频电信号。微音器电位的其它一些特点是:潜伏期极短,小于0.1ms;没有不应期;对缺O2和深麻醉相对地不敏感,以及它在听神经纤维变性时仍能出现等。
图9-19 由短声刺激引起的微音器电位和听神经动作电位
CM:微音器电位 AP:耳蜗神经动作电位(包括N1、N2、N3三个负电位)
A与B对比表明,声音位相改变时,微音器电位位相倒转,但神经动作
电位位相没有变化 C:在白噪音作用下,AP消失,CM仍存在
用微电极进入毛细胞的细胞内电变化记录的实验证明,所谓微音器电位就是多个毛细胞在接受声音刺激时产生的感受器电位的复合表现;在记录单一毛细胞跨膜电位的情况下,发现听毛只要有0.1。的角位移,就可引起毛细胞出现感受器电位,而且电位变化的方向与听毛受力的方向有关,亦即此电位既可是去极化的;这就说明了为什么微音器电位的波动同声波振动的频率和幅度相一致。
由于听毛的角位移和产生感受器电位之间只有一极短潜伏期,因而认为后者的产生是由于毛细胞顶部膜中有机械门控通道的存在,听毛受力引起该处膜的轻微变形,就足以改变这种通道蛋白质的功能状态,引起跨膜离子移动和相应的电位反应。在毛细胞,它的感受器电位可引起细胞基底部的递质(可能是谷氨酸和门冬氨酸)释放量的改变,进而引起分布在附近的耳蜗传入纤维产生动作电位,传向听觉高级枢,产生听觉。至于内毛细胞和外毛细胞在功能上有何不同,有人首先注意到它们所接受的传入纤维的数目有极大差异。据计算,人一侧耳蜗内毛细胞的总数约为3500个,外毛细胞则有约15000个,但来自螺旋神经节的约32000条听神经传入纤维中约有90%分布到内毛细胞的底部,这说明一个内毛细胞可接受多条传入纤维的分布,而多个外毛细胞才能接受一个传入纤维的轴突分支。因此一般认为,内毛细胞的作用是把不同频率的声音振动转变为大量分布在它们底部的传入纤维的神经冲动,向中枢传送听觉信息,而息细胞的作用近年来却发现有些特殊。有人发现毛细胞在基底膜振动和听毛受力而出现微音器电位时,此细胞可产生形体长短的快速改变,超极化引起细胞伸长,去极化引起细胞缩短,它们的形体改变因此也和外来声音振动的频率和振幅同步。据认为,外毛细胞的这种形体改变可以使所在基底膜部分原有的振动增强,亦却对行经该处的行波起放大作用,这显然使位于该部分基底膜上的内毛细胞更易受到刺激,提高了对该振动频率的敏感性。外毛细胞因膜内外电位差改变引起的机制尚不清楚,但这使得基底膜不仅仅是以固定的结构“被动”地对外界的振动产生行波,它还可以“主动”地增强行波的振动幅度。
四、听神经动作电位
听神经纤维的动作电位,是耳蜗对声音刺激一一系列反应中最后出现的电变化,是耳蜗对声音刺激进行换能和编码作用的总结果,中枢的听觉感受只能根据这些传入来引起。图9-19中的N1、N2、N3……是从整个吸神经上记录到复合动作电位,并非由单一听神经纤维的兴奋所产生。用不同频率的纯音刺激耳蝇,同时检查不同的单一听神经纤维的冲动发放情况,就可检查行波理论是否正确,也可阐明耳蜗编码作用的某些特点。仔细分析每一条听神经纤维的放电牧场生和声音频率之间的关系时,发出如果声音强度够大时,同一纤维常可对一组频率相近的纯音刺激起反应,但如果将声音强度逐渐减弱,则可找到一个纤维的最佳反应频率,即当别的刺激频率都因强度太弱而不引起动作电位时,该频率仍能引起。每一条纤维的最佳反应频率的高低,决定于该纤维末稍的基底膜上分布位置,而这一部分正好是该频率的声音所引起的最大振幅行波的所在位置。由这类实验得出的初步结论是,当某一频率的声音强度较弱时,神经信号由少数对该频率最敏感的神经纤维向中枢传递,当这一频率的声音强度增大时,除了能引起上述纤维兴奋外,还能引起更多一些其最佳反应频率与该频率相近的神经纤维也发生兴奋,使更多的纤维参加到传音的频率及其强度的行列里来,结果是由这些纤维传递的神经冲动,共同向中枢传递这一声音的频率及其强度的信息。在自然情况下,作用于人耳的声音捱牟频率和强度的变化是十分复杂的,因此基底膜的振动形式和由此而引起听神经纤维的兴奋及其组合也是十分复杂的;人耳可以区别不同音色,其基础可能亦在于此。
第四节 前庭器官
内耳迷路中除耳蜗外,还有三个半规管、椭圆囊和球囊,后三者合称为前庭器官,是人体对自身运动状态和头在空间位置的感受器。当机体进行旋转或直线变速运动时,速度的变化(包括正、负加速度)会刺激三个半规圆或椭圆囊中的感受细胞;当头的位置和地球引力的作用方向出现相对关系的改变时,就会刺激球囊中的感受细胞。这些刺激引起的神经冲动沿第八脑神经的前庭支传向中枢,引起相应的感受和其他效应。
一、前庭器官的感受装置和适宜刺激
前庭器官的感受细胞都称为毛细胞,具有类似的结构和功能。这些毛细胞通常在顶部有60-100条纤细的毛,按一定的形式排列;其中有一条最长,位于细胞顶端的一侧边缘处,称为动毛,其余的毛较短,占据了细胞顶端的大部分区域,称静毛。图9-20是在一个半规管壶腹中的毛细胞上所做的实验,当动毛和静毛都处于自然状态时,细胞膜内外存在着约-80mV的静息电位,同时在与此毛细胞相接触的神经纤维上有中等频率的持续放电;此时如果用外力使毛细胞顶部的纤毛由静毛所在一侧倒向动毛一侧,可看到细胞的静息电位去极化到约-60mV的水平,同时有神经纤维冲动发放频率的增加;与此相反,当外力使纤毛弯曲的方向由动毛一侧向静毛一侧时,可看到细胞静息电位向超极化的方向转变,而神经纤维上的冲动发放频率也变得比纤毛处自然不受力状态时为小.这是迷路器官中所有毛感受外界刺激时的一般规律,其换能机制与前面讲到的耳蜗毛细胞类似。在正常情况下,由于各前庭器官中毛细胞的所在位置和附属结构的不同,便得不同形式的变速运动都能以特定的方式改变毛细胞纤毛的倒向,使相应的神经纤维的冲动发放频率发生改变,把机体运动状态和头在空间位置的信息传送到中枢,引起特殊的运动觉和位置觉,并出现各种躯体和内脏功能的反射性改变。
图9-20 前庭器官中毛细胞顶部纤毛受力情况影响 细胞静息电位和神经纤维冲动发放频率的实验示意图
三个半规管的形状大致相同,但各处于一个平面上,这三个平面又互相垂直。每个半规管约占2/3个圆周,一端有一个相对膨大的壶腹。两侧的水平半规管同时在一个平面上,如果人在直立时头前倾30℃,则此平面正好与地面平行;当两臂平举而肘关节呈半屈状态时,此时手臂的方位即相当于水平半规管的方位,两个拳头的位置就相当于两侧壶腹的位置。壶腹内有壶嵴,它的位置和半规管的轴垂直;在壶嵴中有一排毛细胞,面对管腔,而毛细胞顶部的纤毛又都埋植在一种胶质性的圆顶形终帽之中。毛细胞上动毛和静毛的相对位置是固定的,例如在水平半规管内,当充满管腔的内淋巴由管腔向壶腹的方向移动时,正好能使壶嵴中毛细胞顶部的静毛向动毛一侧弯曲,于是引起该侧壶腹的传入神经向中枢发放大量的神经冲动。
水平半规管的结构特点,使它能感受人体以身体长轴为轴所作的旋转变速运动。旋转开始时,由于管腔中内淋巴的惯性作用,它的起动将晚于人体和管本身的运动,因此当人体向左旋转时,左侧水平半规管中的内淋巴将压向壶腹的方向,使该侧毛细胞兴奋而产生较多的神经冲动;与此同时,右侧水平半规管中的内淋巴压力作用方向正好是离开壶腹,于是由该侧壶腹传向中枢的冲动减少。人脑正是根据来自两侧水平半规管传入信号的不同,“判定”人体是否开始旋转和向何方旋转的。当旋转变为匀速旋转时,管腔中内淋巴与整个管同步运动,于是两侧壶腹中的毛细胞都处于不受力状态,中枢获得的信息与不进行旋转时无异。但当人体停止旋转时,内淋巴运动的停止又由于惯性作用晚于管本身,于是两侧壶腹中的毛细胞又有受力情况的改变,其受力方向和冲动发放情况正好与旋转开始时相反。内耳迷路中尚有其他两对半规管,可以接受和它们所处平面方向一致的旋转变速运动的刺激。
在椭圆囊和球囊,毛细胞存在于囊斑结构中,其纤毛则埋植在一种称为耳石膜的结构内。耳石膜是一块胶质板,内含耳石,主要由蛋白质和碳酸钙所组成,比重大于内淋巴,因而也有较大的惯性。椭圆囊和球囊的不同,在于其中囊斑所在的平面和人体的相对关系不一样。人体在直立位时,椭圆囊中囊斑所处平面呈水平,囊斑表面分布的毛细胞顶部朝上,耳石膜在纤毛上方;球囊与此不同,其中囊斑所处平面在人体直立时位置和地面垂直,毛细胞和第十毛由囊斑表面向水平方向伸出,耳石膜悬在纤毛外侧,与囊斑相平行。仔细检查两个囊斑平面上分布着的各毛细胞顶部静毛和动毛的相对位置关系时,发现这在每一个毛细胞几乎都不相同(图9-21)。毛细胞纤毛的这种配置,使得它们有可能分辨人体在囊斑平面所作的各种方向的直线变速运动。例如,当人体在水平方向以任何角度作直线变速运动时,由于耳石膜的惯性,在椭圆囊囊斑上总会有一些毛细胞由于它们的静毛和动毛的独特的方位,正好能发生静毛向动毛侧的最大弯曲,于是由此引起的某些特定的传入神经纤维的冲动发放增加,引起机体产生进行着某种方向的直线变速运动的感觉。球囊囊斑上的毛细胞,则由于类似的机制,可以感受头在空间位置和重力作用方向之间的差异,因而可以“判断”头以重力作用方向为参考点的相对位置变化。
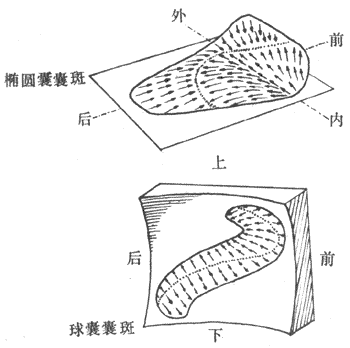
图9-21 椭圆囊和球囊中囊斑的位置以及毛细胞顶部纤毛的排列方向
箭头所指方向是该处毛细胞顶部动毛所在位置,箭尾是同一细胞的静毛所在位置,
当机体所作直线加速运动的方向与某一箭头的方向一致时,该箭头所代表的毛细胞
表面静毛向动毛侧的弯曲最明显,与此毛细胞有关的神经纤维有最大频率的冲动发放
二、前庭反应和眼震颤
来自前庭器官的传入冲动,除与运动党内和位置觉的引起有关外,还引起各种姿势调节的反射和植物性功能的改变。延髓动物(见第十章)表面的某些状态反射和中礅动物的翻正反射,就与前庭器官的传入冲动有关。人体在前庭器官受到刺激时,也会出现一些躯体调节反应,如人乘车而车突然加速时,会有背肌紧张增强而后仰,车突然减速时又有相反的情况;当电梯突然上升时,肢体伸肌抑制而屈曲,下降时伸肌紧张加强而伸直,等等。
前庭反应是中最特殊的是躯体旋转运动时出现的眼球的特殊运动,称为眼震颤,常被用来判断前庭功能是否正常。眼震颤主要由半规管的刺激引起,而且眼震颤的方向也由于受刺激半规管的不同而不同。当人体头部前倾30℃而围绕人体垂直轴旋转时,主要是两侧的水平半规管壶嵴毛细胞有刺激强度的改变,这时出现的也是水平方向的眼震颤。具体情况是,当旋转开始时,如果是向左侧旋转,则是左侧壶嵴的毛细胞受刺激增强而右侧正好相反,这时出现两侧眼球缓慢向右侧移动,这称为眼震颤的慢动相;当慢动相使眼球移动到两眼裂右侧端而不能再移时,又突然返回到眼裂正中,这称为眼震颤的慢动相;此后再出现新的慢动相和快动相,返复不已,这就是眼震颤。当旋转变为匀速运动时,旋转虽在继续,但由于两则壶嵴所受压力一样,于是眼球不再震颤而居于眼裂正中。只有当旋转停止而出现减速时,内淋巴又由于惯性作用而不能立刻停止运动,于是两侧壶嵴又再现所受压力的不同,但情况正好与旋转开始时相反,于是又引起一阵由方向相反的慢动相和快动相组成的眼震颤(图9-22)。临床和特殊从业人员常进行眼震颤试验以判断前庭功能是否正常。在同样条件下震颤时间过长或过短,说明前庭功能有过敏或减弱,前庭器官受到过强或过长刺激,或刺激未过量而前庭功能过敏时,常会引起恶心、呕吐、眩晕、皮肤苍白等现象,称为前庭植物神经性反应,具体表现为晕船、晕车和航空病。
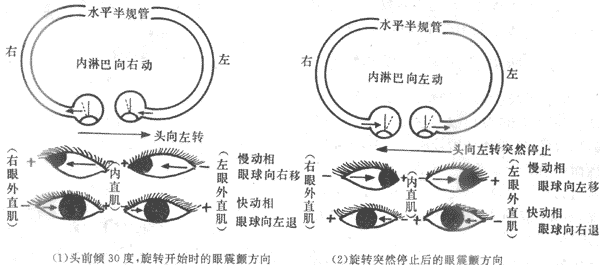
(1)头前倾30℃,旋转开始时的眼震颤方向(2)旋转突然停止后的眼震颤方向
图9-22 旋转变速运动时两侧水平半规管壶嵴毛细胞受刺激情况和眼震颤方向示意图
第五节 嗅觉和味觉
一、嗅觉感受器和嗅觉的特点
嗅觉感受器位于上鼻道及鼻中隔后上部的嗅上皮,两侧总面积约5cm2。由于它们的位置较高平静呼吸时气流不易到达。因此在嗅一些不太显着的气味时,要用力吸气,使气流上冲,才能到达嗅上皮。嗅上皮含有三种细胞,即主细胞、支持细胞和基底细胞。主细胞也称呼嗅细胞(图9-23),呈圆瓶状,细胞顶端有5-6条短的纤毛,细胞的底端有长突,它们组成嗅丝,穿过筛骨直接进入嗅球。嗅细胞的纤毛受到存在于空气中的物质分子刺激时,有神经冲动传向嗅球,进而传向更高级的嗅觉中枢,引起嗅觉。
不同动物的嗅觉敏感程度差异很大,同一动物对不同有气味物质的敏感程度也不同。嗅上皮和有关中枢究竟怎样感受并能区分出多种气味,目前已有初步了解。有人分析了600种有气味物质和它们的化学结构,提出至少存在7种基本气味;其他众多的气味则可能由这些基本扬眉吐气的组合所引起。这7种基本气味是:樟脑味、麝香味、花卉味、薄荷味、乙醚味、辛辣味和腐腥味;他们发现,大多数具有同样气味的物质,具有共同的分子结构有特殊结合能力的受体蛋白(理论上至少有7种),这种结合可通过G-蛋白而引起第二信使类物质的产生,最后导致膜上某种离子通道开放,引起Na+、K+等离子的跨膜移动,在嗅细胞的胞体膜上产生去极化型的感受器电位,后者在轴突膜上引起不同频率的动作电位发放,传入中枢。用细胞内记录法检查单一嗅细胞电反应的实验发现,每一个嗅细胞只对一种或两种特殊的气味坡反应;还证明嗅球中不同部位的细胞只对某种特殊的气味起反应。嗅觉系统也其他感觉系统类似,不同性质的气味刺激有其相对专用的感受位点和传输线路;非基本气味则由于它们在不同线路上引起的不同数量冲动的组合特点,在中枢引起特有的主观嗅觉感受。
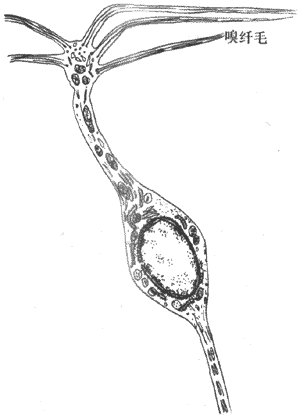
图9-23 嗅细胞(双极细胞)
二、味觉感受器和味觉的特点
味觉的感受器是味蕾,主要分布在舌背部表面和知缘,口腔和咽部粘膜的表面也有散在的味蕾存在。儿童味蕾较成人为多,老年时因萎缩而逐渐减少。每一味蕾由味觉细胞和支持细胞组成(图9-24)。味觉细胞顶端有纤毛,称为味毛,由味蕾表面的孔伸出,是味觉感受的关键部位。
舌表面不同部分对不同味刺激的敏感程度不一样。在人,一般是舌尖部对甜味道比较敏感,舌两侧对酸味比较敏感。舌两侧前部对咸味比较敏感,而软腭和舌根部对苦味比较敏感。味觉的敏感度往往受食物或刺激物本身温度的影响。在20-30℃之间,味觉的敏感度最高。另外,味觉的辨别能力也受血液化学成分的影响,例如,动物实验中正常大鼠能辩出1:2000的氯化钠深夜,而切除上腺皮质的大鼠,可能是由于血液中低Na+,可辨别出1:33000的氯化钠深夜,主动选饮这种含盐多的深夜。因此,味觉的功能不仅在于辨别不同的味道,而且与营养物的摄取和内环境恒定的调节也有关系。
人和动物 味觉系统可以感受和区分出多种味道;但很早以前就知道,众多的味道是由四种基本的味觉组合而成的,这就是甜、咸、酸和苦。不同物质的味道与它们的分子结构的形式有关,便也有例外。通常NaCI能引起典型的咸味;甜味的引起与葡萄糖的主体结构有关;而奎宁和一些有毒植物的生物碱的结构能引起典型的苦味。有趣的是,这4种基本味觉的换能或跨膜信号的转换机制并不一样,如咸和酸的刺激要通过特殊化学门控通道,甜味的引起要通过受体、G-蛋白和第二信使系统,而苦味则由于物质结构不同而通过上述两种形式换能。和前面讲过的嗅觉刺激的编码过程类似,中枢可能通过来自传导四种基本味觉的专用神经通路上的神经信号和不同组合来“认知”这些基本味觉的以外的多种味觉。
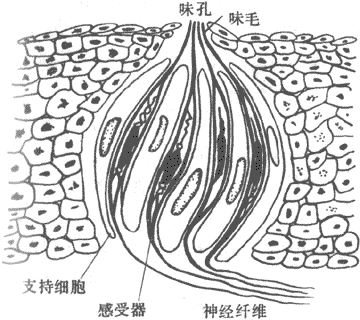
图9-24 味蕾的结构
第六节 皮肤感受
皮肤内分布着多种感受器,能产生多种感觉。一般认为皮肤感觉主要有四种,即触觉、冷觉、温觉和痛觉。用不同性质的点状刺激仔细检查人的皮肤感觉时发现,不同感觉的感受区在皮肤表面呈互相独立的点状分布;如用纤细的毛轻触皮肤表面时,只有当某些特殊的点被触及时,才能引起触觉。用类似的方法,可找到冷觉点、热点和痛点等。用组织学方法曾发出皮肤中有大量游离的感觉神经末稍和种种特殊形式的感觉小体(图9-25),因此曾推测每一种性质的感觉应当同某一特定形式的感受结构相对应;但近年来的实验结果不支持这样的观点,例如,同样的游离神经末稍可以在不同的感觉点下方发现,而触点下方可以找到游离神经末稍或种种特殊形式的感觉小体。看来,不同感受器的功能特异性应到膜的分子水平上支找,各种附属结构可能只影响感受末梢敏感性的高低和适应性的快慢。皮肤感受器的换能机制除触-压觉是通过机构门控通道外,其他尚不清楚。
触觉是微弱的机械刺激兴奋了皮肤浅层的触觉感受器引起的,压觉是指较强的机械刺激导致深部组织变形时引起的感觉,两者在性质上类似,可统称为触-压觉。触点在皮肤表面会布密度和该部位对触觉的敏感程度成正比,如颜面、口唇、指尖等处密度较高,手背、背部密度较低。皮肤在接受每秒5-40次的机械振动刺激时,还可引起振动觉,据认为这与触觉感受器有关。与触觉有关的传入纤维既有髓的Ⅱ、Ⅲ类纤维,也有纤细的N类无髓纤维。
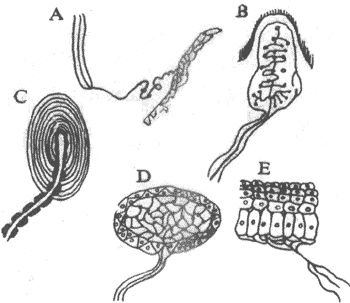
图9-25 皮肤中的各种感受装置
A:Ruffini小体 B:Meissner小体 C:环层小体即Pacini小体
D:Krause球 E:皮肤的游离神经末梢
冷觉和温觉合称温度觉,这起源于两种感受范围不同的温度感受器,因为冷不能构成一种能量长工。冷感受器在皮肤温度低于30℃时开始引起冲动发放,热感受器在超过30℃时开始引起冲动发放,47℃时频率最高。一般皮肤表面冷点约较热点多4-10倍;冷点下方主要分布有游离神经末梢,由Ⅲ类纤维传导传入冲动;热感受器可能也主要是游离神经末梢,传导纤维以N类为主。
痛觉是由有可能损伤或已经造成皮肤损伤的各种性质的刺激所引起的,它们除引起不愉快的痛苦感觉外,尚伴有强烈的情绪反应。
参考资料
1.杨雄里.光感受机制研究的重要突破.生理科学进展1987;18:1-14
2.Dowling JE,吴淼鑫,杨雄里译。视网膜。上海:上海医科大学出版社,1989
3.缪天荣,对数视力表及5分记录法。中华眼科杂志1966;13:96-104
4.Berne RM,Levy ML.Physiology 3rded,CV Mosby Co,St Louis,1993
5.Roper SD.The cell biology ofvertebrate.Ann Rev Neurosci 1989;12:329-353
6.Daw NW,Brunken WJ,ParkinsnD.The funcfion of synaptic transmitters in the retina.Ann Rev Neurosci1989;12:205-225
7.Dallos P.The active cochlea.JNeurosci 1992;12:4575-4585
8.Kinnamon SC,CummingsTA.Chemosensory transduction mechanisms in taste.Ann Rev Physiol1992;54:715-731
9.Von Beseky G.Experiments inHearing.McGraw-Hill Publishing Co,New York,1960
10.Zufall F,Firestein S,Shepherd G.Cyclicnucleotide-gated ion channels and sensory transduction in olfactory receptorneurons.Ann Rev Biophys and Biomol Structure 1944;23:577-607
第十章 神经系统
第一节 神经元活动的一般规律
人体各器官、系统的功能都是直接或间接处于神经系统的调节控制之下,神经系统是整体内起主导作用调节系统。人体是一个极为复杂的有机体,各器官、系统的功能不是孤立的,它们之间互相联系、互相制约;同时,人体生活
■[此处缺少一些内容]■
(二)神经纤维传导的速度
用电生理方法记录神经纤维的动作电位,可以精确地测定各种神经纤维的传导速度,不同种类的神经纤维具有不同的传导速度(表10-1,表10-2)。一般地说,神经纤维的直径越大,其传导速度也越大;这是因为直径大时神经纤维的内阻就小,局部电流的强度和空间跨度就大。有髓纤维的传导速度与直径成正比,其大致关系为:传导速度(m/s)=6×直径(μm)。一般据说扔髓纤维的直径是指包括轴索与髓鞘在一起的总直径,而轴索直径与总直径的比例与传导速度又有密切关系,最适宜的比例为0.6左右。
直径相同的恒温动物与变温动物的有髓纤维其传导速度亦不相同;如猫的A类纤维的传导速度为100m/s,而蛙的A类纤维只有40m/s。神经纤维的传导速度与温度有关,温度降低则传导速度减慢。
经测定,人的上肢正中神经的运动神经纤维和感觉神经纤维的传导速度分别为58m/s和65m/s。当周围神经发生病变时传导速度减慢。因此测定传导速度有助于诊断神经纤维的疾患和估计神经损伤的预后。
表10-1神经纤维的分类(一)
| 纤维分类 | A类(有髓纤维) | B类(有髓纤维) | C类(无髓纤维) | ||||||
| Aα | Aβ | Aγ | Aδ | SC | drC | ||||
| 来源 | 初级肌梭传入纤维和支配梭外肌的传出纤维 | 皮肤的触压觉传入维生素 | 支配梭内肌的传出纤维 | 皮肤痛温觉传入纤维 | 自主神经节前纤维 | 自主神经节后纤维 | 后根据中传导痛觉的传入纤维 | ||
| 纤维直径(μm) | 13-22 | 8-13 | 4-8 | 1-4 | 1-3 | 0.3-1.3 | 0.4-1.2 | ||
| 传导速度(m/s) | 70-120 | 30-70 | 15-30 | 12-30 | 3-15 | 0.7-2.3 | 0.6-2.0 | ||
| 锋电位持续时间(ms) | 0.4-0.5 | 1.2 | 2.0 | ||||||
| 负后电位 | %锋电位高度 | 3-5 | 3-5 | 无 | |||||
| 持续时间(ms) | 12-30 | -- | 50-80 | -- | |||||
| 正后电位 | %锋电位高度 | 0.2 | 1.5-4.0 | 1.5 | 10-30 | ||||
| 持续时间(ms) | 40-60 | 100-300 | 300-1000 | 75-100 | |||||
表10-2神经纤维的分类(二)
| 纤维类别 | 来源 | 直径(μm) | 传导速度(m/s) | 电生理学上的分类 |
| I | 肌梭及腱器官的传入纤维 | 12-22 | 70-120 | Aα |
| Ⅱ | 皮肤的机械感受器传入纤维(触、压、振动感受器传入纤维) | 5-12 | 25-70 | Aβ |
| Ⅲ | 皮肤痛温觉传入纤维,肌肉的深部压觉传入纤维 | 2-5 | 10-25 | Aδ |
| N | 无髓的痛觉纤维,温度、机械感受器传入纤维 | 0.1-1.3 | 1 | C |
(三)神经纤维的分类
1.根据电生理学的特性分类主要是根据传导速度(复合动作电位内各波峰出现的时间)和后电位的差异,将哺乳类动物的周围神经的纤维分为A、B、C三类(表10-1)。
A类:包括有髓鞘的躯体的传入和传出纤维,根据其平均传导速度又进一步分为α、β、γ、δ四类。
B类:有髓鞘的自主神经的节前纤维。
C类:包括无髓鞘的躯体传入纤维(drC)及自主神经节后纤维(sC)。
D类纤维的直径<3μm,传导速度<15m/s,与Aδ纤维非常近似,但两者的锋电位及后电位很不相同。Aδ纤维的锋电位时程较长,后负后电位,而有一个大的正后电位。
2.根据纤维的直径的大小及来源分类将传入纤维分为I、Ⅱ、Ⅲ、N四类(表10-2),I类纤维中包括Ia和Ib两类。
上述两种分类法在实际使用中存在一些问题,例如C类和N类纤维都可用来表示无髓纤维,Aα和I类纤维又常用来表示传导速度最快的纤维,从而造成混乱。因此,目前对传出纤维采用第一种分类法,对传入纤维则采用第二种分类法。
(四)神经纤维的轴浆运输
神经元的细胞体与轴突是一个整体,胞体和轴突之间必须经常进行物质运输和交换。实验证明,轴突内的轴浆是经常在流动的。轴浆流动是双向的,一方面部分轴浆由胞体流向轴突末梢,另一方面部分轴浆由轴突末梢反向流向胞体。胞体内具有高速度合成蛋白质的结构,其合成的物质借轴浆流动向轴突末梢运输;而反向的轴浆流动可能起着反馈控制胞体合成蛋白质的作用。在组织培养或在体的神经纤维中,用显微镜观察确实见到轴浆内颗粒具有双向流动的现象。用同位素标记的氨基酸注射到蛛网膜下腔中,可以见到注射物质首先被神经元的细胞体报到,而在胞体内出现,然后逐渐在轴突近端轴浆内出现,最后在远端轴浆内出现,说明轴浆在流动。如果轴突中断,思浆双向流动被阻断,则远侧断端和近侧断端及胞体都受到影响;因此变性反应不仅发生在远端正,也发生在胞体。
目前知道,自胞体向轴突末梢的轴浆运输分两类。一类是快速轴浆运输,指的是具有膜的细胞器(线粒体、递质囊泡、分泌颗粒等)的运输,在猴、猫等动物的坐骨神经内其运输速度为410mm/d。另一类是慢速轴浆运输,指的是由胞体合成的蛋白质所构成的微管和微丝等结构不断向前延伸,其他轴浆的可溶性成分也随之向前运输,其速度为1-12mm/d。
轴浆流动的机制目前还不十分清楚。在缺氧、氰化物毒化等情况下,神经纤维的有氧代谢扰乱使ATP减少到50%以下时,快速轴奖学金流动即停止,说明它是一种耗能过程。有人提出与肌肉收缩滑行理论相似的假说,来解释快速轴浆流动。认为囊泡等有膜的细胞器的运输与微管成微丝的功能有关,微管的成分与肌纤蛋白相似,微管上含有结合点和ATP,囊泡膜上有ATP酶和能与微管相附着的结合点;ATP酶作用于ATP,后者放出能量使微管与囊泡膜发生附着结合,而后又脱离接触,如此推动囊泡不断与下一个结合点相附着,造成囊泡等有膜细胞器沿着微管向前推移。
目前对由轴突末梢向细胞体方向的逆向轴浆流动了解得比较少。这种逆向流动的速度约为快速顺向运输速度的一半左右。有人认为,破伤风毒素、狂犬病病毒由外周向中枢神经系统转运的机制,可能就是逆向轴浆流动。近年来,运用辣根过氧化酶方法研究神经纤维的发源部位,其原理也是因为辣根过氧化酶能被轴突末梢摄取,并由轴浆流动转运到神经纤维的细胞体。
二、神圣元间相互作用的方式
(一)经典的突触概念
神经元之间在结构上并没有原生质相连,每一神经元的轴突末梢仅与其他神经元的胞体或突起相接触,引相接触的部位称为突触。主要的突触组成可分为三类:①轴突与细胞体相接触;②轴突与树突相接触;③轴突与轴突相接触(图10-1)。突触有特殊的微细结构,一个神经元的轴突末梢首先分成许多小支,每个小支的末梢部分膨大呈球状,称为突触小体,贴附在下一个神经元的胞体或树突表面。在电子显微镜下观察到,突触的接触处有两层膜,轴突末梢的轴突膜称为突触前膜,与突触前膜相对的胞体膜或树突膜则称为突触后膜,两膜之间为突触间隙。一个突触即由突触前膜、突触间隙和突触后膜三部分组成。突触前膜和后膜较一般的神经元膜稍增厚,约7.5nm左右。突触间隙约20nm左右,其间有粘多糖和糖蛋白(图10-2)。在突触前膜内侧有致密突起,致密突起和网格形成囊泡栏栅,其间隙处正好容纳一个囊泡;因此设想,这种栏栅结构具有引导囊泡与突触前膜接触的作用,促进囊泡内递质的释放。在突触小体的轴浆内,含有较多的线粒体和大量聚集的囊泡(突触小泡)。突触小泡的直径为20-80nm,它们含有高浓度的递质(图10-3)。不同突触内含的泡大小和形状不完全相同,释放乙酰胆碱的突触,其小泡直径约为30-50nm,在电镜下为均匀致密的囊泡;而释放去甲肾上腺素的小泡,直径为30-60nm,其中有一个直径为15-25nm的致密中心。突触小泡在轴浆中分布不均匀,常聚集在致密突起处。
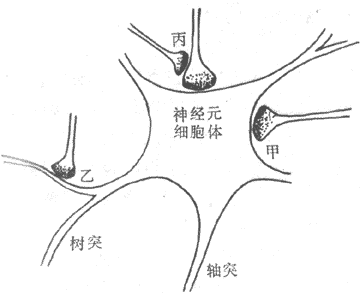
图10-1 突触类型
甲:轴突与细胞体相接触 乙:轴突与树相接触丙:轴突与轴突相接触
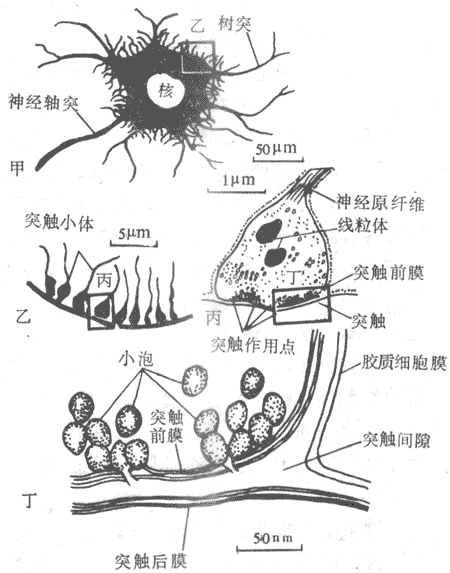
图10-2 神经突触示意图
甲、乙:光学显微镜所见 丙、丁:电子显微镜所见
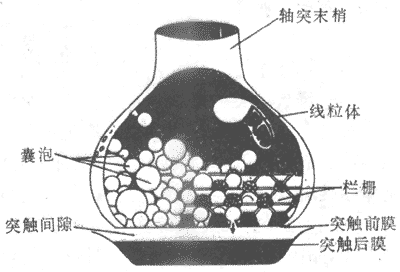
图10-3 突触结构模式图
显示囊泡栏栅引导囊泡与突触前膜接触
由于突触传递功能有兴奋性的抑制性两种,因此有人认为,突触在形态上也可能存在两种类型。例如,有人观察了小脑皮层内突触的形态特征,见到所有平行纤维与哺肯野细胞之间的兴奋性突触的小泡呈圆于形,而篮状细胞与哺氏细胞之间的抑制性突触小泡呈扁平形;由此认为,兴奋性与抑制性突触的突触小泡有形态学上的区别。兴奋性突触的前膜释放兴奋性递质,它对突触后膜的作用是产生兴奋性突触后电位;抑制性突触的前膜释放抑制性递质,它对突触后膜的作用是产生抑制性突触后电位(参见本章第二节)。
一个神经元的轴突末梢一般都分支形成许多突触小体,与其后的神经元构成突触,所以一个神经元能通过突触传递作用于许多其他神经元。另一方面,一个神经元的树突或胞体可以接受许多神经元的突触小体构成突触,因此一个神经元又可接受许多不同神经元的作用。据估算,一个脊椎前角的运动神经元的胞体和树突上可有2000个左右突触,而一个大脑皮层锥体细胞则约有30000个突触。
(二)是突触
神经元之间除了上述的经典突触联系外,还存在电突触。电突触的结构基础是缝隙连接(参见第二章),是两个神经元膜紧密接触的部位。两层膜间的间隔只有2-3nm,连接部位的神经元膜没有增厚,其旁轴浆内无突触小泡存在(图10-4)。连接部位存在在沟通两细胞胞浆的通道,带电离子可通过这些通道而传递电信号,这种电信号传递一般是双向的。因此,这种连接部位的信息传递是一种电传递,与经典突触的化学递质传递完全不同。电突触的功能可能是促进不同神经元产生同步性放电。电传递的速度快,几乎不存在潜伏期。电突触可存在于树突与树突、胞体与胞体、轴突与胞体、轴突与树突之间。
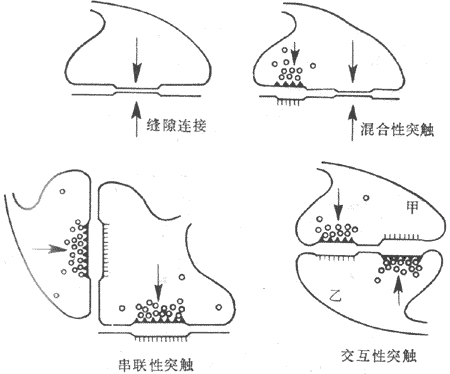
图10-4 几种不同形式突触的模式图箭头示传递方向
(三)非突触性化学传递
由于荧光组织化学等新方法的应用,目前已明确除了经典的突触能进行化学传递外,还存在非突触性化学传递(non-synaptic chemical transmission )。关于这方面的研究,首先在交感神经肾上腺素能神经元上进行。实验观察到,肾上腺素能神经元的轴突末梢有许多分支,在分支上有大量的念珠状曲张体(varicosity)。曲线体内含有大量的小泡(图10-5),是递质释放和部位。一个神经元的轴突末梢可以具有30000个曲线体,因此一个神经元具有大量的递质释放部位。但是,曲张体并不与效应细胞形成经典的突触联系,而是处在效应细胞附近。当神经冲动抵达曲张体时,递质从曲张体释放出来,通过弥散作用到效应细胞的受体,使效应细胞发生反应。由于这种化学传递不是通过经典的突触进行的,因此称为非突触性化学传递。在中枢神经系统内,也有这种传递方式存在。例如,在大脑皮层内具有直径很细的无纤维,这种纤维是去甲肾上腺素能性的,纤维分支上具有许多曲张体,能释放去甲肾上腺素递质;这种曲张体绝大部分不与支配的神经元形成经典的突触,所以进行的是非突触性化学传递。又如在黑质中,多巴胺能纤维也有许多曲张体,且绝大多数也进行非突触性化学传递。此外,中枢内5-羟色胺能纤维也能进行非突触性化学传递。由此看来,单胺类神经纤维都能进行非突触性化学传递。已知,非突触性化学传递也能在轴突末梢以外的部位进行,轴突膜也能释放化学递质(如释放胞浆中的乙酰胆碱),树突也能释放化学递质(如黑质中、树突可释放多巴胺)。
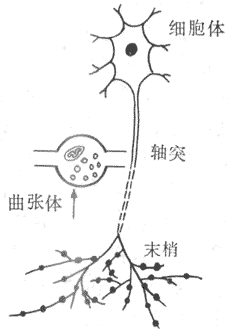
图10-5 交感神经肾上腺能神经示意图
非突触性化学传递与突触性化学传递相比,有下列几下特点:①不存在突触前膜与后膜的特化结构;②不存在一对一的支配关系,一个曲张体能支配较多的效应细胞;③曲张体与效应细胞间的距离至少在20nm以上,距离大的可达几十微米;④递质弥散的距离大,因此传递花费的时间可大于1s;⑤递质弥散到效应细胞时,能否发生传递效应取决于效应细胞上有无相应的受体。
(四)局部回路神经元和局部神经元回路
中枢神经系统中存在长轴突的神经元,也有大量短轴突和无轴突的神经元。长轴突的神经元是投射性神经元,它们投射到远隔部位,起到联系各中枢部位功能的作用;其轴突末梢通过经典的突触联系和非突触性化学传递的方式,完成神经元间的相互作用。短轴突和无轴突神经元不投射到远隔部位,它们的轴突和树突仅在某一中枢部位内部起联系作用;这些神经元称为局部回路神经元(local circuit neuron),例如大脑皮层内的星状神经元、小脑皮层内的篮状细胞和星状细胞、视网膜内的水平细胞和无长突细胞、嗅球内的颗粒细胞、脊髓内的闰绍细胞等。从进化来看,动物越高等,局部回路神经元数量越多,它们的突起越发达。局部回路神经元的活动可能与高级神经功能有密切的关系,例如学习、记忆等。
由局部回路神经元及其突起构成的神经元间相互作用的联系通路,称为局部神经元回路(local nuronal circuit)。这种回路可由几个局部回路神经元构成,例如小脑皮层内的颗粒细胞、篮状细胞、星状细胞等构成的回路。这种回路也可由一个局部回路神经元构成,例如脊髓内闰绍细胞构成的回路。这种回路还可通过局部回路神经元的一个树突或树突的某一部分构成,这种神经元间相互作用的实现不需要整个神经元参与活动。
通过对局部神经回路的研究,现已阐明除了轴突-胞体型、轴突-树突型、轴突-轴突型突触联系外,还存在树突-树突型、树突-胞体型、树突-轴突型、胞体-树突型、胞体-胞体型、胞体-轴突型联系;而且这种联系除了主要属于化学传递性质外,还有属于电传递性质的(电突触)。它们的组合形式也比较复杂,可以形成串联性突触(serial synapses)、交互性突触(reciprocal synapses)、混合性突触(mixed synapses)(图10-5)。以交互性突触为例,局部神经元回路仅在甲、乙两树突的某一部分形成;甲树突通过树突-树突型突触作用于乙树突,乙树突被作用后又通过附近的树突-树突型突触反过来作用于甲树突。这样甲乙两树突通过交互性突触构成了相互作用的局部神经元回路。这种回路不需要整个神经元参与活动,就能完成局部的整合作用。
树突多数不能产生动作电位,因为树突膜上电压门控式钠通道很少。因此,树突上的兴奋或抑制活动是以电紧张性形式扩布的,这种扩布是衰减性的。上述交互性突触中相邻两突触的相互作用就是以电紧张形式实现的。
三、 神 经 递 质
前文已述及突触传递是通过突触前膜释放化学递质来完成的(非突触性化学传递的情况也是如此)。一个化学物质被确认为神经递质,应符合以下条件:①在突触前神经元内具有全盛递质的前体物质和合成酶系,能够合成这一递质;②递质贮存于突触小泡以防止被胞浆内其它酶系所破坏,当兴奋冲动抵达神经末梢时,小泡内递质能释放入突触间隙;③递质通过突触间隙作用于突触后膜的特殊受体,发挥其生理作用,用电生理微电泳方法将递质离子施加到神经元或效应细胞旁,以模拟递质释放过程能引致相同的生理效应;④存在使这一递质失活的酶或其他环节(摄取回收);⑤用递质拟似剂或受体阻断剂能加强或阻断这一递质的突触传递作用。在神经系统内存在许多化学物质,但不一定都是神经递质,只有符合或基本上符合以上条件的化学物质才能认为它是神经递质。关于神经递质,首先是在外周迷走神经对心脏抑制作用的环节上发现的。
(一)外周神经递质
1.乙酰胆碱在蛙心灌注实验中观察到,刺激迷走神经时蛙心活动受到抑制,如将灌流液转移到另一蛙心制备中去,也可引致后一个蛙心的抑制。显然在迷走神经兴奋时,有化学物质释放出来,从而导致心脏活动的抑制。后来证明这一化学物质是乙酰胆碱,乙酰胆碱是迷走神经释放的递质。以后在许多其他器官中(例如胃肠、膀胱、颌下腺等),刺激其副交感神经也可在灌注液中找到乙酰胆碱。由此认为,副交感神经节后纤维都是释放乙酰胆碱作为递质的。释放乙酰胆碱作为递质的神经纤维,称为胆碱能纤维(图10-6)。后来有
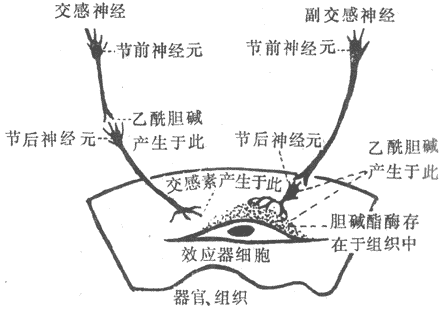
图10-6 自主神经系统神经末梢的化学传递
人进行了上颈交感神经节的灌流,见到刺激节前纤维可以灌流液中获得乙酰胆碱,所以节前纤维的递质也是乙酰胆碱。现已明确躯体运动纤维也是胆碱能纤维。节前纤维和运动神经纤维所释放的乙酰胆碱的作用,与菸碱样作用(N样作用);而副交感神经节后纤维所释放的乙酰胆碱的作用,也毒蕈碱的药理作用相同,称为毒蕈碱样作用(M样作用)。
2.去甲肾上腺素交感神经节后纤维的递质比较复杂。本世纪初,有人见到肾上腺素对效应器的广泛作用与交感神经的作用极为相似,因此设想交感神经可能是通过末梢释放肾上腺素而对效应器起作用的。后来,在猫的实验中观察到,刺激支配尾巴的交感神经可以引致尾巴上毛的竖立和血管收缩,同时该动物的去神经支配的心脏活动加速;如果将自尾巴回流的静脉结扎,再刺激这一交感神经就只能引致尾巴上毛的竖立和血管收缩,却不能引致心脏活动的加速。由此设想,支配尾巴的交感神经末梢能释放一种化学物质,由静脉回流于心脏,这种物质在当时称为交感素。交感素比乙酰胆碱的性质稳定,当有大量释放时不易破坏,在一般情况下有可能经血液循环作用于较为远隔的效应器官。后来,在刺激支配其他器官的交感神经时,均证明静脉血中出现交感素。曾有人指出,交感素是去甲肾上腺素和肾上腺素的混合物,而主要是去甲肾上腺素。现已明确,在高等动物中由交感神经节后纤维释放的递质仅是去甲肾腺上素,而不含肾上腺素;因为在神经末梢只能合成去甲肾上腺素,而不能进一步合成肾上腺素,由于末梢中不含合成肾上腺素所必需的苯乙醇胺氮位甲基移位酶。释放去甲肾上腺素作为递质的神经纤维,称为肾上腺素能纤维。但是,不是所有的交感神经节后纤维都是肾上腺素能纤维,像支配汗腺的交感神经和骨骼肌的交感舒血管纤维却是胆碱能纤维。
3.嘌呤类和肽类递质自主神经的节后纤维除胆三能和肾上腺素能纤维外,还有第三类纤维。第三类纤维末梢释放的递质是嘌呤类和肽类化学物质。有人在实验中观察到,刺激这类神经时实验标本灌流液中可以找到三磷酸腺苷及其分解产物;而三磷酸腺苷对有肠肌的作用与这类神经的作用极相似,两者均可引致肠肌的舒张和肠肌细胞电位的超极化。因此认为这类神经末梢释放的递质是三磷酸腺苷,是一种腺嘌呤化合物。但也有人认为这类神经释放的递质是肽类化合物,因为免疫细胞化学的研究证实自主神经某些纤维末梢的大颗粒囊泡中含有血管活性肠肽,刺激迷走神经时能引致血管活性肠肽的释放。血管活性肠肽能使胃肠平滑肌舒张,胃的容受性舒张可能就是由于迷走神经节后纤维释放血管活性肠肽递质而实现的。第三类纤维是非胆碱能和非肾上腺素能纤维,主要存在于胃肠,其神经元细胞体位于壁内神经丛中;在胃肠上部它接受副交感神经节前纤维的支配。
(二)中枢神经递质
1.乙酰胆碱 闰绍细胞(Renshaw cell)是脊髓前角内的一种神经元,它接受前角运动神经元轴突侧支的支配,它的活动转而反馈抑制前角运动神经元的活动。目前知道,前角运动神经元支配骨骼肌的接头处递质为乙酰胆碱,则其轴突侧支与闰绐细胞发生突触联系,也必定释放乙酰胆碱作为递质(图10-7)。用电生理微电泳法将乙酰胆碱作用于闰绍细胞,确能引致其放电;用N型受体阻断剂后,乙酰胆碱的兴奋作用即被阻断,说明这一突触联系的乙酰胆碱作用与神经肌接头处一样都是N样作用
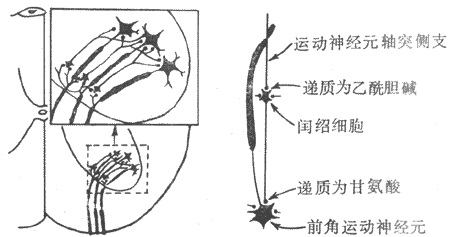
图10-7 脊髓前角运动神经元与闰绍细胞的反馈联系
位于丘脑后部腹侧的特异感觉投射神经元是胆碱能神经元,它们和相应的皮层感觉区神经元形成的突触是以乙酰胆碱为递质的。例如,刺激视神经时,枕叶皮层17区等处的乙酰胆碱释放增多。
脑干网状结构上行激动系统(参见第三节)的各个环节似乎都存在乙酰胆碱递质。例如,脑干脑状结构内某些神经元对乙酰胆碱敏感;刺激中脑网状结构使脑电出现快波时,皮层的乙酰胆碱释放明明显增加;用组织化学法显示脑干网状结构的乙酰胆碱上行通路,发现其与脑干网状结构上行激动系统通路有相似之外。
尾核含有丰富的乙酰胆碱、胆碱乙酰移位酶和胆碱酯酶,尾核内有较多的神经元对乙酰胆碱敏感,壳核与苍白球内某些神经元也对乙酰胆碱敏感。由此看来,纹状体内存在乙酰胆碱递质系统。
此外,边缘系统的梨状区、杏仁核、海马内某些神经元对乙酰胆碱也起兴奋反应,这种反应能被阿托品阻断,说明这些部位也可能存在乙酰胆碱递质系统。
综上所述,乙酰胆碱肯定是中枢的递质,而且分布比较广泛。
2.单胺类单胺类递质是指多巴胺、去甲肾上腺素和5-羟色胺。由于动物实验中采用了荧光组织化学方法,目前对中枢内单胺类递质系统了解得比较清楚(图10-8)。
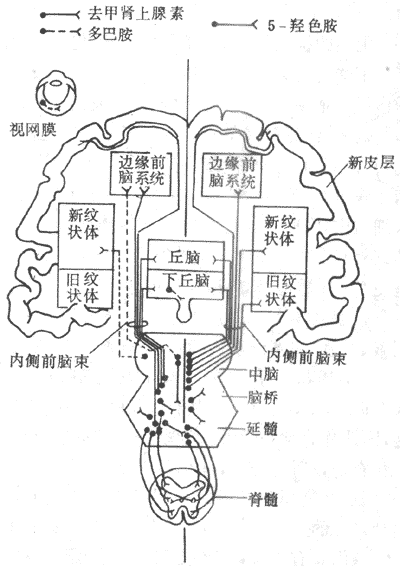
图10-8 单胺类递质的通径
多巴胺递质系统主要包括三部位:黑质-纹状体部分、中脑边缘系统部分和结节、漏斗部分。黑质-纹状体部分的多巴胺能神经元位于中脑黑质,其神经纤维投射到纹状体。脑内的多巴胺主要由黑质制造,沿黑质-纹状体投射系统分布,在纹状体贮存(其中以尾核含量最多)。破坏黑质或切断黑质-纹状体束,纹状体中多巴胺的含量即降低。用电生理微电泳法将多巴胺作用于纹状体神经元,主要起抑制反应。中脑位于边缘部分的多巴胺能神经元位于中脑脚间核头端的背侧部位,其神经纤维投射到边缘前脑。结节-漏斗部分的多巴胺能神经元位于下丘脑弓状核,其神经纤维投射到正中隆起。
去甲肾上腺素系统比较集中,极大多数的去甲肾上腺素能神经元位于低位脑干,尤其是中脑网状结构、脑桥的蓝斑以及延髓网状结构的腹外侧部分。按其纤维投射途径的不同,可分为三部分:上行部分、下行部分和支配低位脑干部分。上行部分的纤维投射到大脑皮层,边缘前脑和下丘脑。下行部分的纤维下达脊髓背角的胶质区、侧角和前角。支配低位脑干部分的纤维,分布在低位脑干内部。
5-羟色胺递质系统也比较集中,其神经元主要位于低位脑干近中线区的中缝核内。按其纤维投射途径的不同,也可分为三部分:上行部分、下行部分和支配低位脑干部分。上行部分的神经元位于中缝核上部,其神经纤维投射到纹状体、丘脑、下丘脑、边缘前脑和大脑皮层。脑内5-羟色胺主要来自中缝核上部,破坏中缝核上部可使脑内5-羟色胺含量明显降低。下行部分的神经元位于中缝核下部,其神经纤维下达脊髓背角的胶质区、侧角和前角。支配低位脑干部分的纤维,分布在低位脑干内部。
3.氨基酸类 现快明确存在氨基酸类递质,例如谷氨酸、门冬氨酸、甘氨酸和γ-氨基丁酸。
在脑脊髓内谷氨酸含量很多,分布很广,但相对来看,大脑半球和脊髓背侧部分含量较高。用电生物微电泳法将谷氨酸作用于皮层神经元和脊髓运动神经地,可引致突触后膜出现类似兴奋性突触后电位的反应,并可导致神经元放电。由此设想,谷氨酸可能是感觉传入神经纤维(粗纤维类)和大脑皮层内的兴奋型递质。
用电生理微电泳法将甘氨酸作用于脊髓运动神经元,可引致突触后膜出现类似抑制性突触后电位的反应。闰绍细胞轴突末梢释放的递质就是甘氨酸,它对运动神经元起抑制作用。
γ-氨基丁酸在大脑皮层的浅层和小脑皮层的浦肯野细胞层含量较高。用电生理微电泳法将γ-氨基丁酸作用于大脑皮层神经元和前庭外侧核神经元(直接受小脑皮层浦肯野细胞支配),可引致突触后膜超极化。由此设想,γ-氨基丁酸可能是大脑皮层部分神经元和小脑皮层浦肯野细胞的抑制性递质。此外,纹状体-黑质的纤维,也是释放γ-氨基西酸递质的。
上述的抑制是突触后膜发生超极化而发生的,因此是突触后抑制。所以甘氨酸和γ-氨基丁酸均是突触后抑制的递质。已知,γ-氨基丁酸也是突触前抑制的递质;当γ-氨基丁酸作用于轴突末梢时可引致末梢支极化,使末梢在冲动抵达时递质释放量减少,从而产生抑制效应(参见第二节)。γ-氨基丁酸对细胞体膜产生超极化,而对末梢轴突膜却产生去极化,其机制尚不完全清楚。有人认为,γ-氨基丁酸的作用是使膜对CI-的通透性增升高;在细胞体膜对CI-的通透性升高时,由于细胞外CI-浓度比细胞内CI-浓度高,CI-由细胞外进入细胞内,因此产生超极化;在末梢轴突膜对CI-通透性升高时,由于轴浆内CI-浓度比轴突外CI-高,CI-由轴突内流向轴突外,因此产生去极化。所以γ-氨基丁酸的作用是使CI-通透性升高,造成超极化还是去极化,取决于细胞内外CI-的浓度差。
4.肽类早已知道神经元能分泌肽类化学物质,例如视上核和室旁核神经元分泌升压素(九肽)和催产素(九肽);下丘脑内其他肽能神经元能分泌多种调节腺垂体活动的多肽,如促甲状腺释放激素(TRH,三肽)、促性腺素释放激素(GnRH,十肽)、生长抑素(GHRIH,十四肽)等。由于这些肽类物质在分泌后,要通过血液循环才能作用于效应细胞,因此称为神经激素。但现已知,这些肽类物质可能还是神经递质。例如,室旁核有向脑干和脊髓投射的纤维,具有调节交感和副交感神经活动的作用(其递质为催产素),并能抑制痛觉(其递质为升压素)。在下丘脑以外脑区存在TRH和相应的受体,TRH能直接影响神经元的放电活动,提示TRH可能是神经递质。
脑内具有吗啡样活性的多肽,称为阿片样肽。阿片样肽包括β-内啡肽、脑啡肽和强啡肽三类。脑啡肽是五肽化合物,有甲硫氨酸脑啡肽(M-ENK)和亮氨酸脑啡肽(L-ENK)两种。脑啡肽与阿片受体常相伴而存在,微电泳啡肽可命名大脑皮层、纹状体和中脑导水管周围灰质神经元的放电受到抑制。脑啡肽在脊髓背角胶质区浓度很高,它可能是调节痛觉纤维传入活动的神经递质。
脑内还有胃肠肽存在,例如胆囊收缩素(CCK)、促胰液素、胃泌素、胃动素、血管活性肠肽、胰高血糖素等。CCK有抑制摄食行为的作用。许多胆碱能神经元中含有血管活性肠肽,它可能具有加强乙酰胆碱作用的功能。此外,脑内还有其他肽类物质,例如P物质、神经降压素、血管紧张素Ⅱ等。P物质是十一肽,它可能是第一级感觉神经元(属于细纤维类)释放的兴奋性递质,与痛觉传入活动有关。神经降压素在边缘系统中存在。血管紧张素Ⅱ的主要作用可能在于调节单受类纤维的递质释放。
5.其他可能的递质近来年研究指出,一氧化氮具有许多神经递质的特征。某些神经元含有一氧化氮合成酶,该酶能使精氨酸生成一氧化氮。生成的一氧化氮从一个神经元弥散到另一神经元中,而后作用于鸟苷酸环化酶并提高其活力,从而发挥出生理作用。因此,一氧化氮是一个神经元间信息沟通的传递物质,但与一般递质有区别:①它不贮存于突触小泡中;②它的释放不依赖于出胞作用,而是通过弥散;③它不作用于靶细胞膜上的受体蛋白,而是作用于鸟苷酸环化酶。一氧化氮与突触活动的可塑性可能有关,因为用一氧化氮合成酶抑制剂后,海马的第时程增强效应被完全阻断(参见第六节中“学习和记忆的机制”)。此外,组织胺也可能是脑内的神经递质。
(三)递质与调质的概念
递质是指神经末梢释放的特殊化学物质,它能作用于支配的神经元或效应细胞膜上的受体,从而完成信息传递功能。调质是指神经元产生的另一类化学物质,它能调节信息传递的效率,增强或削弱递质的效应。但是也有人把递质概念规定得非常严格,认为只有作用于膜受体后导致离子通道开放从而产生兴奋或抑制的化学物质才能称为递质;其他一些作用于膜受体后通过第二信使转而改变膜的兴奋性或其它递质释放的化学物质,均应称为调质。根据后一种观点,递质为数不多,氨基酸类物质是递质,神经肌接头部位释放的乙酰胆碱也是递质,而肽类物质一般均属于调质。但是一般来说,递质与调质无明确划分的界限,调质是从递质中派生出来的概念,不少情况下递质包含调质;前文就没有把两者严格区分开来,统称为递质。
(四)递质的共存
长期来认为,一个神经元内只存在一种递质,其全部神经末梢均释放同一种递质。这一原则称为戴尔原则(Dale’s principle)。近来来,通过免疫组织化学方法观察到,一个神经元内可存在两种或两种以上递质(包括调质),因此认为戴尔原则并不正确。但是戴尔的原先观点认为,一个神经元的全部神经末梢均释放相同的递质;他并没有限定一个神经元只能含一种递质。因此,戴尔的观点还是对的,而戴尔原则则是需要修改的。
在无脊椎动物的神经元中,观察到多巴胺和5-羟色胺递质可以共存。在高等动物的交感神经节神经节发育过程中,去甲肾上腺素和乙酰胆碱可以共存。此外,在大鼠延髓的神经元中观察到5-羟色胺和P物质共存;在上颈交感神经节中神经元中观察到去甲肾上腺素和脑啡肽共存。有人认为肽类递质可能都是与其他递质共存的。递质共存的生理意义,目前尚未清楚了解;可能两种递质在同时释放后起着不同的生理作用,有利于发挥突触传递作用。
(五)递质的合成、释放和失活
1.递质的合成 乙酰胆碱是由胆碱和乙酰辅酶A在胆碱乙酰移位酶(胆碱乙酰化酶)的催化作用下合成的。由于该酶存在于胞浆中,因此乙酰胆碱在胞浆中合成,合成后由小泡摄取并贮存起来。去甲肾上腺素的合成以酪氨酸为原料,首先在酪氨酸羟化酶的催化作用下合成多巴,再在多巴脱羧酶(氨基酸脱竣酶)作用下合成多巴胺(儿茶酚乙胺),这二步是在胞浆中进行的;然后多巴胺被摄取入小泡,在小泡中由多巴胺β羟化酶催化进一步合成去甲肾上腺素,并贮存于小泡内。多巴胺的合成与去甲肾上腺素揆民前二步是完全一样的,只是在多巴胺进入小泡后不再合成去甲肾上腺素而已,因为贮存多巴胺的小铴内不含多巴胺β羟化酶。5-羟色胺的合成以色氨酸为原料,首先在色氨酸羟化酶作用下合成5-羟色氨酸,再在5-羟色胺酸脱竣酶(氨基酸脱竣酶)作用下将5-羟色氨酸合成5-羟色胺,这二步是在胞浆中进行的;然后5-羟色胺被摄取入小泡,并贮存于小泡内。γ-氨基丁酸是谷氨酸在谷氨酸脱羧催化作用下合成的。肽类递质的全盛与其他肽类激素的合成完全一样,它是由基因调控的,并在核糖体上通过翻译而合成的。
2.递质的释放当神经冲动抵达末梢时,末梢产生动作电位和离子转移Ca2+由膜外进入膜内,使一定数量的小泡与突触前膜紧贴融合起来,然后小泡与突触前膜粘合处出现破裂口,小泡内递质和其他内容物就释放到突触间隙内。突触前膜释放递质的过程,称为出胞(exocytosis)或胞裂外排。在这一过程中,Ca2+的转移很重要。如果减少细胞外Ca2+浓度,则递质释放就受到抑制;而增加细胞外Ca2+的浓度则递质释放增加。这一事实说明,Ca2+由膜外进入膜内的数量多少,直接关系到递质的释放量;Ca2+是小泡膜与突触前膜紧贴融合的必要因素。一般认为,Ca2+可能有两方面的作用:①降低轴浆的粘度,有利于小泡的移动;②消除突触前膜内的负电位,便于小泡与突触前膜接触而发生融合。小泡破裂把递质和其他内容物释放到突触间隙时,其外壳仍可留在突触前膜内(也可与突触前膜融合,成为突触前膜的组成部分),以后仍旧可以重新恢复原样,继续合成并贮存递质(图10-9)。
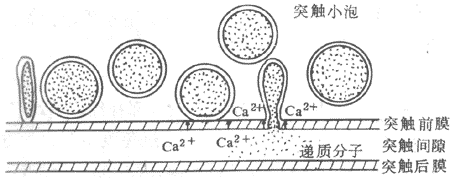
图10-9 突触处递质释放过程
3.递质的失活 进入突触间隙的乙酰胆碱作用于突触后膜发挥生理作用后,就被胆碱酯酶水解成胆碱和乙酸,这样乙酰胆碱就被破坏而推动了作用,这一过程称为失活。去甲肾上腺素进入突触间隙并发挥生理作用后,一部分被血液循环带走,再在肝中被破坏失活;另一部分在效应细胞内由儿茶酚胺内由儿茶酚胺位甲基移位酶和单胺氧化酶的作用而被破坏失活;但大部分是由突触前膜将去甲肾上腺素再摄取,回收到突触前膜处的轴浆内并重新加以利用。多巴胺的失活与去甲肾上腺素的失活相似,它也是由儿茶酚胺氧位甲基移位酶和单胺氧化酶的作用而被破坏失活。突触前膜敢能再摄取多巴胺加以重新利用。5-羟色胺的失活也与去甲肾上腺素的失活相似,单胺氧化酶等能使5-羟色胺降解破坏,突触前膜也能再摄取5-羟色胺加以重新利用。氨基酸递质在发挥作用后,能被神经元和神经胶质再摄取而失活。肽类递质的失活是依靠酶促降解,例如通过氨基肽酶、羧基肽酶和一些内肽酶的降解而失活。
(六)受体学说
1.胆碱能受体 上世纪末发现阿托品能阻断副交感神经节后纤维对效应器的作用,当时认为效应器具具有一种接受物质,阿托品与接受物质结合后就阻断了副交感神经的作用。研究证实了这一设想,例如刺激支配颌下腺的副交感神经则唾液分泌量增加,如果先用阿托品后再刺激神经则唾液分泌量不再增加,而此时末梢乙酰胆碱的释放量并不见减少。这说明阿托品不影响神经末梢递质的释放过程,而是直接作用于效应器上。效应器上的接受物质后来就称为受体。
递质的受体一般是指突触后膜或效应器细胞膜上的某些特殊部分,神经递质必须通过与受体相结合才能发挥作用。受体的本质和发挥作用和机制已在第二章详述。如果受体事先被药物结合,则递质就很难再与受体相结合,于是递质就不能发挥作用。这种能与受体相结合,从而占据受体或改变受体的空间结构形式,使递质不以发挥作用的药物称为受体阻断剂。
受体阻断剂 的不断发现,对递质与受体的作用关系有了更多的了解。前文述及乙酰胆碱有两种作用,实际上是由于存在两种不同的乙酰胆碱能受体而形成的。一种受体广泛存在于副交感神经节后纤维支配的效应细胞上,当乙酰胆碱与这类受体结合后就产生一系列副交感神经末梢兴奋的效应,包括心脏活动的抑制、支气管平滑肌的收缩、胃肠平滑肌的收缩、膀胱逼尿肌的收缩、虹膜环形肌的收缩、消化腺分泌的增加等。这类受体也能与毒蕈碱相结合,产生相似的效应。因此这类受体称为毒蕈碱受体(M型受体,muscarinic receptor),而乙酰胆碱与之结合所产生的效应称为毒蕈碱样作用(M样作用)。阿托品是M型受体阻断剂,它仅能和M型受体结合,从而阻断乙酰胆碱的M样作用。
另一种胆碱能受体存在于交感和副交感神经节神经元的突触后膜和神经肌接头的终板膜上,当乙酰胆碱与这类受体结合后就产生兴奋性突触后电位和终板电位,导致节神经元和骨骼肌的兴奋。这类受体也能与菸碱相结合,产生相似的效应。因此这类受体也称为菸碱型受体(N型受体,nicotinic receptor),而乙酰胆碱与之结合所产生的效应称为菸碱样作用(N样作用)。
通过采用不同受体阻断剂的研究,现已证明M型和N型受体均可进一步分出向种亚型。M型受体至少已分出M1、M2和M3三种亚型。M1受体主要分布在神经组织中;M2受体主要分布在心脏,在神经和平滑肌上也有少量分布;M3受体主要分布在外分泌腺上,神经和平滑肌也有少量分布。N型受体可分出N1和N2两种亚型。神经节神经元突触后膜上的受体为N1受体,终板膜上的受体为N2受体。简箭毒能阻断N1和N2受体的功能,六烃季铵主要阻断N1受体的功能,十烃季铵主要阻断N2受体的功能,从而阻断乙酰胆碱的N样作用。
支配汁腺的交感神经和骨骼肌的交感舒血管纤维,其递质也是乙酰胆碱;由于阿托品能阻断其作用,所以属于M型受体。
2.肾上腺素能受体 多数的交感神经节后纤维释放的递质是去甲肾上腺素,其对效应器的作用既有兴奋性的,也有抑制性的。效应不同的机制是由于效应器细胞上的受体不同。能与儿茶酚胺(包括去甲肾上腺素、肾上腺素等)结合的受体有两类,一类为α型肾上腺素能受体(简称α受体),另一类为β型肾上腺素能受体(简称β受体)。儿茶酚胺与α受体结合的产生的平滑肌效应主要是兴奋性的,包括血管收缩、子宫收缩、虹膜辐射状肌收缩等;但也有抑制性的,如小肠舒张。儿茶酚胺与β受体结合后产生的平滑肌效应是抑制性的,包括血管舒张、子宫舒张、小肠舒张、支气管舒张等;但产生的心肌效应却是兴奋性的。有的效应器仅有α受体,有的仅有β受体,有的α和β受体均有(表10-3)。目前知道,心肌细胞上除有β受体外,也有α受体,但受体的作用较明显。例如,心肌α受体兴奋可引致收缩力加强,但其作用比β受体兴奋的作用要弱;而且心肌β受体兴奋可引致心率加快,而α受体却不能加快心率。
表10-3 肾上腺素能受体的分布及效应
| 效应器 | 受体 | 效应 | |
| 眼 | 虹膜辐射状肌 | α1 | 收缩(扩瞳) |
| 睫状体肌 | β2 | 舒张 | |
| 心 | 窦房结 | β1 | 心率加快 |
| 房室传导系统 | β1 | 传导加快 | |
| 心肌 | α1,β1 | 收缩加强 | |
| 血管 | 冠状血管 | α1 | 收缩 |
| β2(主要) | 舒张 | ||
| 皮肤粘膜血管 | α1 | 收缩 | |
| 骨骼肌血管 | α | 收缩 | |
| β2(主要) | 舒张 | ||
| 脑血管 | α1 | 收缩 | |
| 腹腔内脏血管 | α1(主要) | 收缩 | |
| β2 | 舒张 | ||
| 唾液腺血管 | α1 | 收缩 | |
| 支气管平滑肌 | β2 | 舒张 | |
| 胃肠 | 胃平滑肌 | β2 | 舒张 |
| 小肠平滑肌 | α2 | 舒张(可能是胆碱能纤维的突触前受体,调制乙酰胆碱的释放) | |
| β2 | 舒张 | ||
| 括约肌 | α1 | 收缩 | |
| 膀胱 | 逼尿肌 | β2 | 舒张 |
| 三角区和括约肌 | α1 | 收缩 | |
| 子宫平滑肌 | α1 | 收缩(有孕子宫) | |
| β2 | 舒张(无孕子宫) | ||
| 竖毛肌 | α1 | 收缩 | |
| 糖酵解代谢 | β2 | 增加 | |
| 脂肪分解代谢 | β1 | 增加 |
α和β受体不仅对交感神经末梢释放递质起反庆,也对血液中存在的儿茶酚胺(由肾上腺髓质分泌或注射的药物)起反应。去甲肾上腺素对α受体的作用强,对β受体的作用较弱;肾上腺素对α和β受体的作用都强;异丙肾上腺素主要对β受体有强烈作用。如在动物实验中观察血压的变化,见到注射去甲肾上腺素后血压上升,这是由于α受体被激活引致广泛血管收缩而形成的;如注射异丙肾上腺素,则见到血压下降,这是由于β受体被激海参引致广泛血管舒张而形成的;如注射肾上腺素,则血压先升高后下降,这是由于α和β受体均被激活,引致广泛血管先收缩后舒张而形成的。如果,进一步采用不同的受体阻断剂进行实验,见到α受体阻断剂酚妥拉明可以消除去甲肾上腺素和肾上腺素的升压效应,但不影响肾上腺素和异丙肾上腺素的降压效应;而β受体阻断剂普萘洛尔(propranolol)可以消除肾上腺素和异丙肾上腺素的降压效应,但不影响去甲肾上腺素和肾上腺素的升压效应。由此说明,确实存在两种不贩肾上腺素能受体,即α受体和β受体,两者能分别被特异的受体阻断剂所阻断。
β受体阻断剂已应用于临床。例如,心绞痛患者应用普萘洛尔可以降低心肌的代谢和活动,得到治疗的效果。但普萘洛尔阻断β受体的作用很广泛,应用后可同时此致支气管痉挛,对伴有呼吸系统疾病的患者有危险性。研究发现,有些β受体阻断剂主要阻断心肌的β受体,而对支气管平滑肌的β受体阻断作用很小,例如阿替洛尔(atenilol)、心得宁(practolol);有些受体阻断剂对心肌的β受体阻断作用极小,而对支气管平滑肌的β受体阻断作用却很强,例如心得乐(butoxamine)。由此认为,β受体可分别分为β1和β2两个亚型,其分布及效应见表10-3。在伴有呼吸系统疾病的患者,应采用阿替洛尔,以免发生支气管痉挛。
3.突触前受体研究指出,受体不仅存在于突触前膜,而且存在于突触前膜,突触前膜的受体称为突触前受体(presynaptic receptor)。突触前受体的作用,在地调节神经末梢的递质释放。例如,肾上腺素能纤维末梢的突触前膜上存在α受体,当末梢释放的去甲肾上腺素在突触前膜处超过一定量时,即能与突触前α受体结合,从而反馈抑制末梢全盛和释放去甲肾上腺素,起到调节末梢递质释放能量的作用。在应用α受体阻断剂后,这种反馈抑制环节被阻断;这时刺激肾上腺素能纤维,末梢内合成和释放去甲肾上腺素增加。这种情况在支配心肌的肾上腺素能纤维上也存在,虽然心肌的受体为β受体,而突触前膜上的受体为α受体。由于突触前受体是感受神经末梢自身释放的递质的,因此又称为自身受体(autoreceptor)。
突触前膜的α受体不同于后膜的α受体,前者为α2型,后者为α1型。α受体区分为α1和α2两个亚型,是根据不同受体阻断剂的选择性作用来确定的。如哌唑嗪(prazosin)可选择性阻断α1受体,而育亨宾(yohimbine)可选择性阻断α2受体;酚妥拉明对α1和α2受体均有阻断作用,但对α1受体的作用比对α2受体的作用大3-5倍。必须指出,α2受体也可存在于突触后膜上,例如大脑皮层、子宫、腮腺等处突触后膜可能有α2受体。此外,突触前受体除α2型外,也可有其他类型。
4.中枢内递质的受体中枢递质种类复杂,因此相应的受体也多,除胆碱能N型和M型受体、肾上腺素能α和β受体外,还有多巴胺受体、5-羟色胺受体、兴奋性氨基酸受体、γ-氨基丁酸受体、甘氨酸受体,阿片受体(opiate receptor)等。多巴胺受体可分为D1、D2等受体亚型,5-羟色胺受体可分为5-HT1、5-HT2、5-HT3、5-HT4等受体亚型,兴奋性氨基酸受体可分为N-甲基-D-天冬氨酸型(NMDA)。使君子氨酸型(QA)和海人藻酸型(KA)等,γ-氨基丁酸受体可分为GABAA、GABAB等受体亚型,阿片受体可分为μ、δ、κ等受体亚型。这些受体也有相应的受体阻断剂,例如派迷清(pimozide)能阻断多巴胺受体,肉桂硫胺(cinanserin)能阻断5-羟色胺受体,荷包牡丹硷(bicuculine)能阻断GABAA受体,钠洛酪(naloxone)能阻断阿片μ受体等。
四、神经的营养性作用
神经对其所支配的组织能发挥两方面的作用。一方面是借助于兴奋冲动传导抵达末梢时突触前膜释放特殊的递质,而后作用于突触后膜,从而改变所支配组织的功能活动,这一作用称为功能性作用。另一方面神经还能通过末梢经常释放某些物质,持续地调整被支配组织的内在代谢活动,影响其持久性的结构、生化和生理的变化,这一作用与神经冲动无关,称为营养性作用。神经的营养性作用在正常情况下不易观察出来,但在神经切断后产生的变性与再生过程中就能明显地表现出来。
神经的营养性作用概念是上世纪提出的。有人观察到三叉神经的眼支损伤后会引致角膜炎;当时认为三叉?
■[此处缺少一些内容]■
完成的。如神经切断的部位靠近肌肉,则肌肉的内在代谢改变发展早;如切断的部位远离肌内,则内在代谢改变发展迟。因为前一种情况营养性因子耗尽快,而后一种情况耗尽慢。营养性因子可能是借轴浆流动由神经元细胞体流向末梢,而后由末梢释放到所支配的组织的。
切断运动神经后,肌肉因失去神经的营养性作用而出现萎缩;如经常用适当强度的电刺激来刺激肌肉使其收缩,则能减慢肌肉萎缩的速度。在断肢再植过程中,使用这一方法减慢肌肉萎缩,对断肢功能的恢复是有利的。
此外,在神经交叉缝合的动物实验中,如果将支配慢肌和支配快肌的神经分别切断,然后将支配快肌的中枢端与支配慢肌的神经的外周缝合,待神经再生后,慢肌(指收缩缓慢的骨骼肌)就可转变成快肌(指收缩快速的骨骼肌)。可见神经的营养性作用不仅调整着支配组织的内在代谢活动,而且还决定其生理特性。
神经元能生成营养性因子维持所支配组织的正常代谢和功能,反过来组织也持续产生营养和生长刺激因子作用于神经元。神经生长因子(nerve growth factor,NGF)是最早发出的这类因子之一,后来陆续发现多种这类因子参与神经系统的发育过程,维持神经系统的正常功能。NGF是一种蛋白质,由α、β、γ亚单位组成;其中β亚单位是具有生物活性的成分,分子量为13200,其结构与胰岛素相似。NGF是交感神经和背根神经节神经元产生存发育必要的因子,在动物胚胎期注射NGF抗体以破坏其作用,则交感神经节和背根神经节神经元退化。组织产生的NGF由神经末梢摄取,而后逆向运输到胞体发挥营养性作用。虹膜在去交感神经后,产生NGF增加,促进交感神经定身长入。
第二节 反射活动的一般规律
一、反射概念
反射是指在中枢神经系统参与下的机体对内外环境刺激的规律性应答。17世纪人们即注意到机体对一些环境的刺激具有规律性反应,例如机械刺激角膜可以规律性地引致眨眼。当时就借用了物理学中“反射”一词表示刺激与机体反应间的必然因果关系。后来,巴甫洛夫发展了反射概念,把反射区分为非条件反射和条件反射两类。
非条件反射是的指在出生后无需训练就具有的反射。按生物学意义的不同,它可分为防御反射、食物反射、性反射等。这类反射能使机体初步适应环境,对个体生存与种系生存有重要的生理意义。条件反射是指在出生后通过训练而形成的反射。它可以建立,也能消退,数量可以不断增加。条件反射的建立扩大了机体的反应范围,当生活环境改变时条件反射也跟着改变。因此,条件反射较非条件反射有更大的灵活性,更适应复杂变化的生存环境。
在个体一生中,纯粹的非条件反射仅在新生下来的时候容易见到,以后由于条件反射的不断建立,条件反射与非条件反射越来越不可分地融合在一起,而条件反射起着主导作用。至于人类,也具有非条件反射和条件反射;但是人类还有更高级的神经活动,能通过劳动实践来改造环境,与动物相比又有了质的不同,人类的神经系统活动显然是更进一步发展了。
二、反射弧
反射活动的结构基础称为反射弧,包括感受器、传入神经、神经中枢、传出神经和效应器(图10-10)。简单地说,反射过程是如下进行的:一定的刺激按一定的感受器所感受,感受器发生了兴奋;兴奋以神经冲动的方式经过传入神经传向中枢;通过中枢的分析与综合活动,中枢产生兴奋;中枢的兴奋过程;中枢的兴奋过程又经一定的传出神经到达效应器,使效应器发生相应的活动。如果中枢发生抑制,则中枢原有的传出冲动减弱或停止。在实验条件下,人工遥刺激直接作用于传入神经也可引起反射活动,但在自然条件下,反射活动一般都需经过完整的反射弧来实验,如果反射弧中任何一个环节中断,反射即不能发生。
感觉器一般是神经组织末梢的特殊结构,它能把内外界刺激的信息转变为神经的兴奋活动变化,所在感受器是一种信号转换装置。某一特定反射往往是在刺激其特定的感受器后发生的,这特定感受器所在的部位称为该反射的感受野。
中枢神经系统是由大量神经元组成的,这些神经元组合成许多不同的神经中枢。神经中枢是指调节某一特定生理功能的神经元群。一般地说,作为某一简单反射的中枢,其范围较窄,例如膝跳反射的中枢在腰脊髓,角膜反射的中枢在脑桥。但作为调节某一复杂生命活动的中枢,其范围却很广,例如调节呼吸运动的中枢分散有延髓、脑桥、下丘脑以至大脑皮层等部位内。延髓是发生呼吸活动的基本神经结构,而延髓以上部分的有关呼吸功能的神经元群,则调节呼吸活动使它更富有适应性。
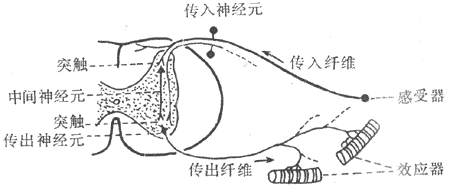
图10-10 反射弧
在整体情况下,反射活动发生时,感觉冲动传入脊髓或脑干后,除了在同一水平与传出部分发生联系并发出传出冲动外,还有上行冲动传导到更高级中枢,乃至大脑皮层的中枢,进一步通过高级水平的整合,再发出下行冲动来调整反射的传出冲动,使反射活动更具有适应性。因此,在反射发生时,既有初级水平的整合活动,也有较高级和最高级水平的整合活动;通过多级水平的整合,反射活动便具有更大的复杂性和适应性。
神经中枢的活动可以通过神经纤维直接作用于效应器,在某些情况下也可通过体液的途径间接作用于效应器,这个体液环节就是指内分泌调节。这时候反射是如下进行的:感受器→传入神经→神经中枢→传出神经→内分泌腺→激素在血液中转运→效应器。反射效应在内分泌腺的参与下,往往就变得比较缓慢、广泛而持久。例如,强烈的痛刺激可以反射性地通过交感神经引致肾上腺髓质分泌增多,从而产生广泛的反应。
三、中枢神经元的联系方式
神经元依其在反射弧中所处地位的不同可区分为传入神经元、中间神经元和传入神经元三种。人体中枢神经系统的传出神经元的数目总计为数十万;传入神经元较传出神经元多1-3倍;而中间神经元的数目最大,单数就以中间神经元组成的大脑皮层来说,就估计约有140亿,这说明了中间神经元具有重要的生理作用。
神经元的数量如此巨大,它们之间的联系也必然非常复杂。一个神经元的轴突可以通过分支与许多神经元建立突触联系,此称为辐射原则;这咱联系有可能使一个神经元的兴奋引起许多神经元的同时兴奋或抑制(图10-11)。同一神经元的细胞体与树突可接受许多不同轴突来源的突触联系,此称为聚合原则;这种联系有可能使许多神经元的作用都引起同一神经元的兴奋而发生总和,也可能使来自许多不同作用神经元的兴奋和抑制在同一神经元上发生整合。
在脊髓,传入神经元纤维进入中枢后,除以分支与本节段脊髓的中间神经元及传出神经元发生突触联系外,又有上升与下降的分支,并以其侧支在各节段脊髓与中间神经元发生突触联系;因此,在传入神经元与其他神经元发生突触联系中主要表现了辐散原则。传出神经元接受不同轴突来源的突触联系,主要表现为聚合原则。至于中间神经元之间的联系则多种多样,有的形成链锁状,有的呈环状(图10-12)。在这些联系形式中,辐散与聚合原则都是同时存在的。兴奋冲动通过链锁状联系,在空间上加大了作用范围。兴奋冲动通过环状联系,一方面可能由于反复的兴奋反馈,在时间上加强了作用的持久性,另一方面可能由于回返的抑制反馈,在时间上使活性及时终止;前者是正反馈,而执行者是负反馈。
图10-11 神经元的联系方式
1:传入纤维 2:传出神经元;3:中间神经元 4:脊神经节细胞
图10-12 中间神经元的联系形式
左:环状 右:链锁状
四、反射弧中枢部分的兴奋传布
在反射活动中兴奋还必须通过反射弧的中枢部分。反射弧中枢部分兴奋的传布,不同于神经纤维上的冲动传导,其基本原因在于反射弧中枢部分的兴奋传布必须经过一次以上的突触接替,而突触传递比冲动传导要复杂的多。
(一)兴奋性突触后电位
脊髓的单突触反射是反射弧最简单的一种反射,其感受器为肌梭,传入神经纤维与前角运动神经元直接发生突触联系,因此便于用来进行突触传递的研究。如用微电极插入脊髓前角运动神经元细胞体内,可以没得其膜电位约为-70mV左右。当刺激相应肌肉肌梭的传入神经以发动单突触反射时,见到兴奋冲动进入脊髓后约0.5ms,细胞体的突触后膜即发生去极化,产以电紧张形式扩布到整个神经元细胞体。此种电位变化称为兴奋性突触后电位(excitatory postsynaptic potential,EPSP),电位的大小决定于传入神经刺激强度的大小。假如,刺激强度小,则突触后电位的幅度较小,并在几个毫秒的时间内逐渐趋于消失;假如刺激强度加大,则参与活动的突触数增多,电位变化总和起来,以致突触后电位的幅度加大。当突触后电位加大到一定程度后(例如膜电位由-70mV去极化到-52mV左右),则在轴突的始段部位产生动作电位,沿神经轴突扩布出去(图10-13)。轴突的始段部位比较细小,当细胞体出现兴奋性突触后电位时该部位出现外向电流的电流密度较大,因此始段部位是第一个爆发动作电位的地方。从上述的突触传递现象来看,神经元之间的突触传递与神经肌接头传递很类似,兴奋性突触后电位也如终板电位一样,乃是突触后膜产生局部兴奋的表现。神经肌接头传递是由神经末梢释放乙酰胆碱,提高终板膜对一价正离子的通透性,从而引致去极化,出现终板电位。突触传递也是由突触前膜释放某种兴奋性递质,提高突触后膜对一价正离子(包括N+a和 K+,尤其是N+a)的通透性,从而引致去极化,出现兴奋性突触后电位。所以兴奋通过突触的机制如下:神经轴突的兴奋冲动→神经末梢突触前膜兴奋并释放化学递→递质经过突触间隙扩散并作用于突触后膜受体→突触后膜对一价正离子的通透性升高,产生局部兴奋,出现兴奋性突触后电位→兴奋性突触后电位在突触后神经元始段转化成锋电位,爆发扩布性兴奋→兴奋传至整个神经元。
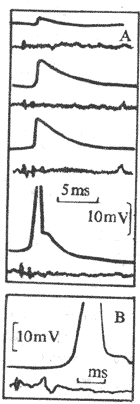
图10-13 兴奋性突触后电位
A:脊髓运动神经元细胞内电位,由传入神经冲动引起,
传入冲动多则兴奋性突触后电位加大,最后出现锋电位
B:扫描加快,以显示兴奋性突触后电位转化成锋电位
上线细胞内电位记录,下线:背根传入神经电位
在交感神经节后神经元和大脑皮层神经元细胞内电位记录中,除了能观察到上述快EPSP外,还可观察到慢突触后电位。慢突触后电位包括慢EPSP和慢IPSP(抑制性突触后电位),它们的潜伏期为100-500ms,持续可达几秒。慢EPSP一般认为是由于膜对K+的通透性下降而造成的,而慢IPSP是由于膜对K+的通透性增加而造成的。这些慢突触后电位的形成机制比较复杂,可能有不同的递质或受体参与。
(二)反射弧中枢部分兴奋传布的特征
1.单向传布在人为刺激神经时,兴奋可由刺激点爆发后沿神经纤维向两个方向传导(双向性);但在中枢内大量存在的化学性突触处,兴奋传布只能由传入神经元向传出神经元方向传布,也即兴奋只能由一个神经元的轴突向另一个神经元的胞体或突起传递,而不能逆和传布,单向传布是由突触传递的性质的决定的,因为只有突触前膜能释放神经递质。但是近来来的研究指出,突触后的靶细胞也能释放一些物质分子(如一氧化氮、多肽等)逆向传递到突触前末梢,改变突触前神经元的递质释放过程。因此,从突触前后的信息沟通角度来看,是双向的。
2.中枢延搁兴奋通过中枢部分比较缓慢,称为中枢延搁。这主要是因为兴奋越过突触要耗费比较长的时间,这里包括突触前膜释放递质和递质扩散发挥作用等环节所需的时间。根据测定,兴奋通过一个突触所需时间约为0.3-0.5ms。因此,反射进行过程通过的突触数愈多,中枢延搁所耗时间就愈长。在一些多突触接替的反射,中枢延搁可达10-20ms;而在那些和大脑皮层活动相联系的反射,可达500ms。所以,中枢延搁就是突触延搁。
3.总和在中枢内,由单根传入纤维的单一冲动,一般不能引起反射性传出效应。如果若干传入纤维同时传入冲动至同一神经中枢,则这些冲动的作用协同起来发生传入效应,这一过程称为兴奋的总和(图10-14)。因为中枢铁神经元与许多没的传入纤维发生突触联系,其中任何一个单独传入的冲动往往只引起该神经元的局部阈下兴奋,亦即产生较小的兴奋性突触后电位,而不发生扩布性兴奋。如果同时或差不多同时有较我的传入纤维兴奋,则各自产生的兴奋性突触后电位就能总和起来,在神经元的轴突始段形成较强的外向电流,从而爆发扩布性兴奋,发生反射的传出效应。局部阈下兴奋状态是神经元兴奋性提高的状态,此时神经元对原来不易发生传出效应的其他传入冲动就比较敏感,容易发生传出效应,这一现象称为易化。兴奋的总和包括空间性总和及时间性总和两类。
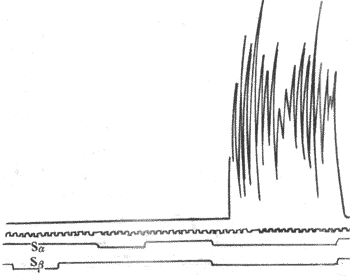
图10-14 反射弧中枢内的兴奋部和分别刺激不同皮肤部位(Sα、Sβ的下降段),不引起反射效应,如两刺激同时应用,则出现反射性肌肉收缩(上线记录),时标(中线)
4.兴奋节律的改变在一反射活动中,如同时分别记录传入与传出的冲动频率,则可测得两者的频率不同。因为传出神经的兴奋节律来自传出神经元,而传出神经元的兴奋节律除取决于传入冲动的节律外,还取决于中间神经元和传出神经元的功能状态。
5.后放(后发放、后放电)在一反射活动中,刺激停止后,传出神经仍可在一定时间内继续发放冲动,这种现象称为后放。后放的原因是多方面的,中间神经元的环状联系是产生后放的原因之一。此外,在效应器发生反射反应时,其本身的感受装置(如肌梭)又受到刺激,兴奋冲动又由传入神经传到中枢,这些继发性传入冲动的反馈作用能纠正和维持原先的反射活动,这也是产生后放的原因之一。
6.对内环境变化的敏感性和易疲劳性在反射活动中,突触部位是反射弧中最易疲劳的环节。同时,突触部位也最易受内环境变化的影响,缺氧、二氧化碳、麻醉剂等因素均可作用于中枢而改变其兴奋性,亦即改变突触部位的传递活动。
五、中枢抑制
在任何反射活动中,中枢内既有兴奋活动又有抑制活动。某一反射进行时,某些其他反射即受抑制,例如吞咽时呼吸停止、屈肌反射进行时伸肌即受抑制(图10-15)。反射活动有一定的次序、一定强度,并有一定的适应意义,是反射的协调功能的表现。反射活动所以能协调,就是因为中枢内既有兴奋活动又有抑制活动;如果中枢抑制受到破坏,则反射活动就不可能协调。例如,用士的宁破坏脊髓抑制活动后,任何一个微弱刺激会导致四肢出现强烈的痉挛性收缩,失去了反射活动的协调性。根据中枢抑制产生机制的不同,抑制可分为突触后抑制和突触前抑制两类。
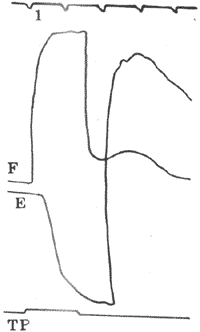
图10-15 拮抗性反射
F为半腱肌的反应(收缩),E为股中间肌的反应(驰缓),TP为刺激传入神经的记号,1为时标
(一)突触后抑制
在哺乳类动物中,所有的突触后抑制都是由抑制性中间神经元活动引起的。由这一抑制性神经元发出的轴突末梢释放的递质,能使所有与其发生突触联系的其他神经元都发生抑制,都暗生抑制性突触后电位。按此观点,一个兴奋性神经元通过突触联系能引起其他神经元产生兴奋,但不能直接引起其他神经元产生突触后抑制;它必须首先兴奋一个抑制性神经元,转而抑制其他神经元。
1.抑制性突触后电位脊髓前角运动神经元有的支配伸肌,有的支配屈肌。来自伸肌肌梭的传入神经冲动,能兴奋伸肌运动神经元,也能同时通过抑制性中间神经元转而抑制屈肌运动神经元。如用微电极插入屈肌运动神经元细胞体内,并刺激伸肌肌梭的传入神经使屈肌运动神经元发生抑制,可见到其细胞体的突触后膜出现超极化现象。这时膜电位的数值向-80mV水平靠近。这种超极化膜电位变化称为抑制性突触后电位(inhibitorypostsynaptic potential,IPSP)。抑制性突触后电位的变化与兴奋性突触后电位变化在时程上极相似,但前者为超极化,后者为去极化,变化方向恰相反(图10-16)。可以设想突触后膜在超极化状态下。轴突始段部位将出现内向电流,造成该处不易爆发动作电位,也就表现为抑制。由于这种抑制是突触后膜出现抑制性突触后电位所造成的,因此称为突触后抑制(postsynaptic inhibition)。抑制性突触后电位是突触后膜对CI-通透性增加而形成的。
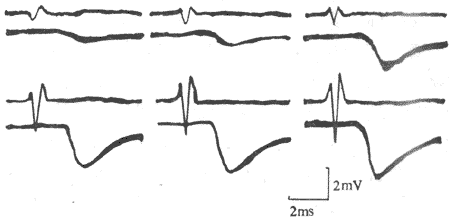
图10-16 抑制性突触后电位
每组曲线的下线为某一屈肌运动神经元细胞内电位记录,上线为刺激拮抗伸肌传入神经时的背根电位记录。当刺激强度逐步加大时,背根电位逐步增大,超极化电位变化也逐步增大
2.突触后抑制的分类根据抑制性神经元的功能和联系方式的不同,突触后抑制可分为传入侧支性抑制(afferent collateral inhibition)和回返性抑制(recurrentinhibition)(图10-17)。
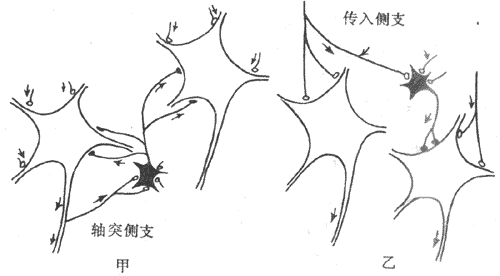
图10-17 两类突触后抑制
甲:回返性抑制 乙:传入侧支性抑制黑色神经元代表抑制性神经元
(1)传入侧支性抑制:是指在一个感觉传入纤维进入脊髓后,一方面直接兴奋某一中枢的神经元,另一方面发出其侧支兴奋另一抑制性中间神经元;然后通过抑制性神经元的活动转而抑制另一中枢的神经元。例如,伸肌的肌梭传入纤维进入中枢后,直接兴奋伸肌的α运动神经元,同时发出侧支兴奋一个抑制性神经元,转而抑制屈肌的α运动神经元,导致伸肌收缩而屈肌舒张;这种抑制曾被称为交互抑制。这种形式的抑制不是脊髓独有的,脑内也有。这种抑制能使不同中枢之间的活动协调起来。
(2)回返性抑制:是指某一中枢的神经元兴奋时,其传出冲动沿轴突外传,同时又经轴突侧支去兴奋另一抑制性中间神经元;该抑制性神经元兴奋后,其活动经轴突反过来作用于同一中枢的神经元,抑制原先发动兴奋的神经元及同一中枢的其他神经元。脊髓前角运动神经元与闰绍细胞之间的联系,
就是这种抑制的典型。前角运动神经元发出轴突支配外周的骨骼肌,同时也在脊髓内发出侧支兴奋闰绍细胞;闰绍细胞是抑制性神经元,其活动经轴突回返作用于脊髓前角运动神经元,抑制原先发动兴奋的神经元和其他神经元。这种形式的抑制在海马和丘脑内也明显存在。这种抑制是一种负反馈控制形式,它能使神经元的活动及时终止,也促使同一中枢内许多神经元之间的活动能步调一致。丘脑与海马内许多神经元的活动能够同步化,就是由于存在回返性抑制环节的缘故。闰绍细胞轴突末梢释放的递质是甘氨酸,其作用能被士的宁和破伤风毒素所破坏;在闰绍细胞功能存在后,将出现强烈的痉挛。
(二)突触前抑制
前文已述及,轴突可与另一神经元的轴突构成突触,这种突触可能是突触前抑制的结构基础。图10-18显示这种突触关系,A纤维末梢与运动神经元构成轴突-胞体型突触,能兴奋该运动神经元;b 纤维传入经过多突触接替后,末梢与A纤维末梢构成轴突-轴突型突触,不能直接影响该运动神经元活动。当A纤维兴奋传入冲动抵达末梢时,可引致运动神经元出现兴奋性突触后电位(图10-18,甲,1);当仅有B纤维兴奋冲动传入时,见不到该运动神经元有反应。如果先使B纤维兴奋,一定时间间隔后再使A纤维兴奋,则A纤维兴奋所引起的兴奋性突触后电位明显减小(图10-18,甲,2,2),说明B纤维的活动能抑制A纤维的兴奋作用。已知,抵达末梢部位的动作电位是触发神经递质释放的因素,如动作电位大则递质释放量大,运动电位小则递质释放量小;而动作电位的大小又受到轴突末梢跨膜静息电位的影响,跨膜静息电位大则动作电位大,跨膜静息电位小则动作电位也小。由此认为,突触前抑制产生的机制是:B纤维传入经多突触接替后,兴奋抵达末梢交释放递质→递质作用于A纤维末梢使其去极化,从而使末梢跨膜静息电位变小→A纤维兴奋时其末梢的动作电位变小,使释放的递质减少→运动神经元的兴奋性突触后电位减小。因此,B纤维的抑制作用是通过使A纤维释放的兴奋性递质减小而实现的。由于这种抑制是改变了突触前膜的活动而实现的,因此称为突触前抑制。
突触前抑制在中枢神经系统内广泛存在,尤其多见于感觉传入途径,对调节感觉传入活动有重要作用。突触前掏可发生在各类感受器传入活动之间,也可发生面类感受器的不同感受野活动之间;即一个感觉传入纤维的兴奋冲动进入中枢后,它本身沿特定的传导路径向高位中枢,同时通过多个神经元的接替,转而对其旁的感觉传入纤维的活动发生突触前抑制限制其他的感觉传入活动。由于突触前抑制产生的潜伏期较长,因此认为传入神经必须通过两个以上中间神经元的多突触接替,才能与其他感觉传入神经末梢形成轴突-轴突型突触联系。突触前抑制一般约在刺激传入神经后20ms左右发展到高峰,而后其抑制作用逐渐减弱,整个抑制过程可持续100-200ms。
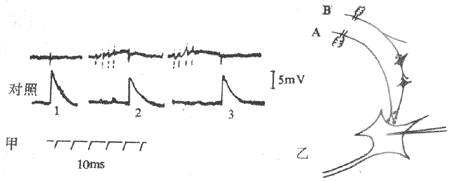
图10-18 突触前抑制
甲:每组曲线的上线为传入冲动电位记录,下线为某肌运动神经元细胞内电位记录
乙:实验方法示意图。微电极插入基本肌运动神经元细胞体内
A:来自该肌的传入神经纤维 B:来自另一肌肉的传入神经纤维
在局部神经元回路中提到了交互性突触联系,这种联系可以由一个兴奋性突触和一个抑制性突触组合而成。在这种联系中,一个树突活动时,首先通过兴奋性突触激活另一个树突;而后一个权威突活动加强时,却通过抑制性突触来抑制前一个树突的活动,使原行发动兴奋的树突很快受到反馈抑制。这种抑制,发生在局部神经元回路中,起到了局部的整合作用;而且这种抑制只有树突的一部分参与活动,不需要整个神经元参与活动。由树突-树突型突触联系产生的抑制,称为树突-树突型抑制(dendrodendritic inhibirion),它在视网膜、嗅球、丘脑内都存在,也是中枢抑制的一种形式。
六、反射活动的反馈调节
当一个刺激发致力一个反射后,效应器的活动必然又刺激本身或本系统内的感觉器发出冲动进入中枢;这个继发性的传入冲动对维持与纠正反射活动的进行有重要作用。实际上每一个反射活动都是链锁反射,一个刺激发动一个反射,反射的效应又成为新的刺激,引起继发性反射活动,使反射链锁样地进行下去实验证明,切除大量传入神经后,就使许多反射活动不可能很好地完成;背根受损的患者,动作苯拙而出现感觉性共济失调,说明肌肉肌梭的传入冲动在运动协调中起着重要作用。事实上,除了准备应器本身的感受装置发出的传入冲动对反射活动的协调有作用外,其他能感知反射效应的感觉器官也发出传入冲动进入中枢,以纠正射活动的进行。例如,视觉和内耳平衡感觉等,能不断感知躯体运动反射活动的进行将受到很大影响。
神经系统对机体的反射调节功能与工程技术上的自动调节装置有相似之处(图10-19)。自动调节装置由五个基本环节构成:发讯装置→联系线路→控制装置→联系线路→交应装置,而且自动调节系统必须具有反馈联系。反馈联系在一般情况下是指由该系统效应装置把一部分效应反过来传给发讯装置,或传给该系统的任何一个中间环节,从而进一步调整自动调节系统的活动。这些特征与反射活动过程是相似的。
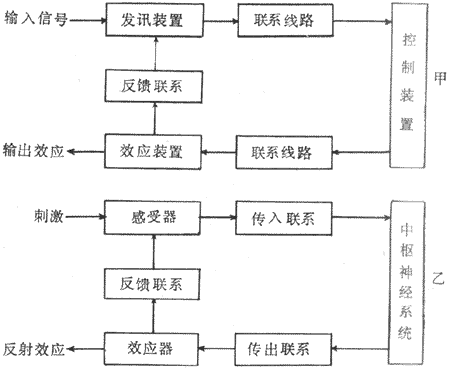
图10-19 反馈联系模式图
甲:自动调节系统 乙:反射调节系统
反馈联系有负反馈和正反馈两种。前者是指自动控制调节系统效应装置的变化,可以减弱作用于发讯装置的变化;后者是指效应装置的变化,可以加强发讯装置的变动,在反射活动过程中反馈联系表现很突出。例如,血压调节的降压反射就是如此。当由于某种原因引致血压上升时,颈动脉窦与主动脉弓区的牵张感受器传入冲动增多,信息沿传入神经传向中枢,通过心血管运动中枢的分析综合活动,控制信息沿传出神经传到效应装置,使心脏活动减退及部分血管扩张,导致血压下降,使血压上升受到限制,可以看出,降压反射是促使血压不致过分升高的控制机制。从另一个角度来看,反射的降压效应本身,又会反过来减弱牵张感觉器所受的刺激,减弱了作用于发讯装置的变化(负反馈联系),使感受器传入冲动有所减少,这样降压反射活动也不会导致血压无限制地下降。因此,在降压反射的调节下,血压就能保持在某一相对稳定的水平上。负反馈联系使自动控制系统具有自发的稳定特性。
正反馈联系在反射活动过程中也有表现,例如排尿反射进行过程中就是如此。当膀胱排尿时,尿液刺激了膀胱壁和尿道内感受装置,传入冲动信息经传入神经传向中枢,通过中枢和传出神经的活动,使膀胱副尿肌收缩加强;这样尿液排出加强,刺激也加强(正反馈联系),使排尿过程越来越强烈,直到尿液排完为止。因此,在具有正反馈联系的条件下,自动控制系统就有自我反复加强的特性,使效应装置活动愈来愈强。正反馈联系一般是在效应装置活动尚未达到最大效应之前发挥作用的。
第三节 神经系统的感觉分析功能
一、脊髓的感觉传导与分析功能
由脊髓上传到大脑皮层的感觉传导路径可分为两在类,一为浅感觉传导路径,另一为深感觉传导路径。浅感觉传导路径传导痛觉、温度觉和以触觉;其传入由后根的外侧部(细纤维部分)进入脊髓,然后在后角更换神经元,再发出纤维在中央管前进行交叉对侧,分别经脊髓丘脑侧束(痛、温觉)和脊髓丘脑前束(轻触觉)上行抵达丘脑。深感觉传导路径传导肌肉本体感觉和深部压觉,其传入纤维由后根的内侧部(粗纤维部分)进入脊髓后,其上行分支在同侧后索上行,抵达延髓下部薄束核和楔束核后更换神经元,再发出纤维进行交叉到对侧,经内侧丘系至丘脑。皮肤触觉中的辨别觉,其传导路径却和深感觉传导路径一致。因此,浅感觉传导路径是先交叉再上行,而深感觉传导路径是行上行再交叉;在脊髓半离断的情况下,浅感觉的障碍发生在离断的同侧(图10-20)。在脊髓空洞症患者,中央管部分有空腔形成,破坏了在中央管前进行交叉的浅感觉传导路径,造成浅感觉障碍。但由于痛、温觉传入纤维进入脊髓后,在进入水平的1-2个节段内更换神经元交叉到对侧,而轻触觉传入纤维进入脊髓后分成上行与下行纤维,分别在多个节段内更换神经元交叉至对侧,因此较局限地破坏中央管前交叉的浅感觉传导路径,仅使相应节段双侧皮节的痛、温觉发生障碍,而轻触觉基本不受影响(辨别觉完全不受影响),造成脊髓空洞症患者出现痛、温觉和触觉障碍的分离现象。
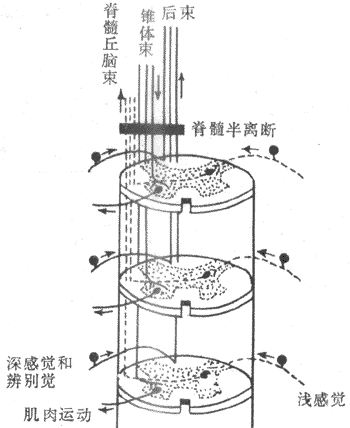
图10-20 脊髓半离断效应示意图
二、丘脑
根据我国神经生理学家张香桐的意见,丘脑的各种细胞群大致可以分为三大类(图10-21)
图10-21右侧丘脑主要核团示意图
1:网状核(大部分已除去,只显示前面一部分) 2:前核3:前腹核 4:苍白球传来纤维
5:外侧腹核 6:外髓板 7:小脑传来纤维8:内髓板及髓板内核群
9:背个侧核 10:后外侧核 11:后外侧腹核12:内侧丘系
13:背内核 14:中央中核 15:束旁核16:后内侧腹核 17:视束
18:外侧膝状体 19:内侧膝状体20:外侧丘系 21:丘脑枕
第一类是接受感觉的投射纤维,并经过换元进一步投射到大脑皮层感觉区的那些细胞群,例如后腹核的外侧与内侧部分(分别称为后外侧腹核和内侧腹核)、内侧膝状体、外侧膝状体等。后外侧腹核为脊髓丘脑束与内侧丘系的换元站,同躯干、肢体感觉的传导有关;后内侧腹核为三叉丘系的换元站,与头面部感觉的传导有关。后腹核发出的纤维向大脑皮层感觉区投射,不同部位传来的纤维在后腹核内换元有一定的空间分布,下肢感觉在后腹核的最外侧,头面部感觉在后腹核内侧,而上肢感觉在中间部位(图10-22);这种空间分布与大脑皮层感觉区的空间定位相对应。内侧膝状体是吸觉路的换元站,发出纤维向大脑皮层听区投射。外侧膝状体是视觉传导路的换元站,发出纤维向大脑皮层视区投射。因此,上述细胞群是所有特定的感觉冲动(除嗅觉外)传向大脑皮层的换元接替部位,称为感觉接替核。

图10-22猴体表在左侧丘脑后腹核(后外侧腹核和后内侧腹核)的投射
第二类细胞群,接受丘脑感觉接替核和其他皮层下中枢来的纤维(但不直接接受感觉的投射纤维),经过换元,发出纤维投射到大脑皮层的某一特定区域。例如,丘脑前核接受下丘脑乳头体来年纤维,并发出纤维投射到大脑皮层的扣带回,参与内脏活动的调节;丘脑的外侧腹核主要接受小脑、苍白球和后腹核的纤维,并发出纤维投射到大脑皮层的运动区,参与皮层对肌肉运动的调节;丘脑枕接受内侧与外侧膝状体的纤维,并发出纤维投射到大脑皮层的顶叶、枕叶和颞叶的中间联络区,参与各种感觉的联系功能。此外,丘脑还有许多细胞群,发出纤维向下丘脑、大脑皮层的前额叶和眶区或顶叶后总部联络区等区域投射。以上这些细胞群投射到大脑皮层的联络区,在功能上与各种感觉在丘脑和大脑皮层水平的联系协调有关,总称为联络核。
第三类细胞群是靠近中线的所谓内髓板以内的各种结构,主要是髓板内核群,包括中央中核、束旁核、中央外侧核等。一般认为,这一类细胞群没有直接投射到大脑皮层的纤维,但也有人认为其中部分核团可向边缘叶、眶回投射。事实上,这些细胞群可以间接地通过多突触接替换元后,然后弥散地投射到整个大脑皮层,起着维持大脑皮层兴奋状态的重要作用。一般认为,这些核群向大脑皮层作弥散性投射,是间接通过丘脑网状核等实现的,但具体投射途径还不完全清楚。对束旁核的研究指出,它可能与痛觉有关;刺激人的丘脑束旁核可加重患者的痛觉症状,而毁损此区后可缓解患者疼痛;动物束旁核的电生理研究观察到,核内确实存在对伤害性传入冲动敏感的细胞。
根据丘脑各部分向大脑皮层投射特征的不同,可把丘脑分成两大系统,一是特异投射系统,另一是非特异投射系统(或称弥散性投射系统),特异投射系统是指第一类细胞群,它们投向大脑皮层的特定区域,具有点对点的投射关系。非特异投射系统是指第三类细胞群,它们弥散地投射到大脑皮层的广泛区域,不具有点对点的投射关系。第二类细胞群在结构上大部分也与大脑皮层有特定的投射关系,投射到皮层的特定区域,所以也可以归属于特异投射系统。
三、感觉投射系统
前文述及的特异和非特异投射系统,都与感觉投射密切相关(图10-23)。
一般认为,经典的感觉传导首是由三个神经元的接替完成的。第一级神经元位于脊髓神经节或有关的脑神经感觉神经节内,第二级神经元位于脊髓后角或脑干的有关神经核内,第三级神经元就在丘脑的感觉接替核内。但特殊感觉(视、听、嗅)的传导道情况比较复杂。视觉传导道包括视杆及视锥细胞在内,则为四个神经元接替;听觉传导道更为复杂,从外周到大脑皮层很难说包含几个神经元接替;嗅觉传导道与丘脑感觉接替核无关。所以,一般经典的感觉传导道就是通过丘脑的特异投射系统而后作用于大脑皮层的;它们都投身到大脑皮层的特定区域,每一种感觉的传导投射系统都是专一的,各种感觉上传都有其专门的途径。
图10-23 感觉投射系统示意图
黑色区代表脑干网状结构 实线代表丘脑
特异投射系统 虚线代表丘脑非特异投射系统
自从对脑干网状结构的研究开展以来,逐步认识到感觉传导向大脑皮层投射还有另一条途径。那就是当上述经典传导道的第二级神经元纤维通过脑干时,发出其侧支与脑干网状结构内神经元发生突触联系;然后在网状结构内反复换元上行,抵达丘脑的第三类细胞群,进一步向大脑皮层作弥散性投射。所以,这一感觉投射途径就是通过丘脑的非特异投射系统而后作用于大脑皮层的。这一投射系统是不同感觉的共同上传途径,也就是说当不同感觉传入脑干部分由侧支进入网状结构后,就不再是专一特异的传导系统,而是由同一上行系统向上传导。电生理研究支持这一结论,当记录脑干网状结构内单个神经元电活动时,来自不同部位的感觉刺激传入冲动都可激海参或?
■[此处缺少一些内容]■
快波;而在中脑头端中断网状结构时,出现类似睡眠的现象,脑电波呈现同步化慢波(图10-24)。由此说明,在脑干网状结构内具有上行唤醒作用的功能系统,这一系统称为脑干网状结构上行激动系统(ascending reticular activaring system)。目前知道,上行激动系统主要就是通过丘脑非特异投射系统而发挥作用的,其作用就是维持与改变大脑皮层的兴奋状态。由于这一系统是一个多突触接替的上行系统,因此易于受药物的影响而发生传导阻滞。例如,巴比妥类催眠药作用可能就是由于阻断了上行激动系统的传导;一些全身麻醉药(如乙醚)也可能是首先抑制了上行激动系统和大脑皮层的活动而发挥麻醉作用的。
图10-24切断特异性传导道或非特异性传导道后猫的行为与脑电图变化
A为切断特异性传导道而不损伤非特异性传导道的猫,处于觉醒状态A′为其脑电图
B为切断非特异性传导道的猫,处于昏睡状态 B′为其脑电图
四、大脑皮层的感觉分析功能
(一)大脑皮层的结构特点与分区
人类大脑皮层内神经元的数量极大,有人估计约为140亿个,其类型也很多,神经元之间具有复杂的联系。但是,各种各样的神经元在皮层中的分布不是杂乱的,而是具有严格层次的。大脑半球内侧面的古皮层比较简单,一般只有三层:①分子层;②锥体细胞层;③多形细胞层。大脑半球外侧面等处的新皮层,具有六层:①分子层;②外颗粒层;③外锥体细胞层;④内颗粒层;⑤内锥体细胞层;⑥多形细胞层。新皮层的不同区域虽然具有相似的六层结构,但不同区域各层的相应厚度并不相同,其中所含神经元的形状与大小与不完全相同。根据神经元成分与结构特征,可以把大脑皮层分成很多区,例如有人把它分成52个区(图10-25)。
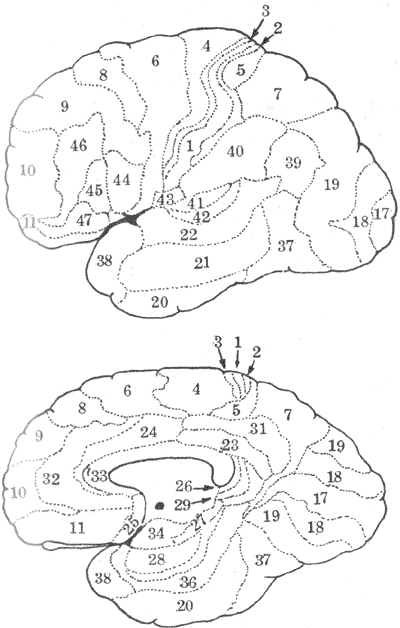
图10-25 人类大脑皮层分区
上:大脑半球外侧面 下:内侧面
对大脑体表感觉区皮层结构和功能的研究指出,皮层细胞的纵向柱状排列构成大脑皮层的最基本功能电位,称为感觉柱(sensory column)。这种柱状结构的直径为200-500μm,垂直走向脑表面,贯穿整个六层。同一柱状结构内的神经元都具有同一种功能,例如都对同一感受野的同一类型感觉刺激起反应。在同一刺激后,这些神经元发生放电的潜伏期很接近,仅相差2-4ms;说明先激活的神经元与后激活的神经元之间仅有几个神经元接替;亦说明同一柱状结构内神经元联系环路只需通过几个神经元接替就能完成。一个柱状结构是一个传入-传出信息整合处理单位,传入冲动先进入第四层,并由第四层和第二层细胞在柱内垂直扩布,最后由第三、第五、第六层发出传出冲动离开大脑皮层。第三层细胞的水平纤维还有抑制相邻细胞柱的作用;因此一柱发生兴奋活动时,其相邻细胞柱就受抑制,形成兴奋和抑制镶嵌模式。这种柱状结构的形态功能特点,在第二感觉区、视区、听区皮层和运动区皮层中也一样存在。
(二)体表感觉
中央后回(3-12区)主要是全身体感觉的投射区域。通过在灵长类动物皮层诱发电位的引导研究,找出中央后回的感觉投射规律如下:①躯体感觉传入冲动向皮层投射具有交叉的性质,即一侧传入冲动向对侧皮层投射,但头而部感觉的投身是双侧性的;②投射区域的大小与不同体表部位的感觉分辨精细程度有关,分辨愈精细有部位在中央后回的代表区也愈大,例如大拇指和食指的代表区面积比胸部十二根脊神经传入支配的代表区总面积大几倍,说明分辨精细有部位具有较大量的感受装置,皮层上与其相联系的神经元数量也必然较多,有利于精细的感觉分析;③投射区域具有一定的分野,下肢代表区在顶部(膝部以下的代表区在皮层内侧面),上肢代表区在中间部,头面部代表区在底部,总的安排是倒置的,然而头而部代表区内部的安排是正立的。人体脑外科手术过程中,用适宜强度的电刺激来刺激皮层,观察到刺激中央后回部顶部可以引致似乎来自下肢的主观感觉,刺激中央后回底部可以引致似乎来自面部的主观感觉(图10-26)。这种主观感觉属于麻木或麻电样感觉,而极少有温觉、冷觉或痛觉的主观感受;而且这种主观感觉并不清晰,与刺激一根感觉神经时的主观感受相似,而和由感受器发生的传入冲动所形成的主观感觉不同。
图10-26大脑皮层体表感觉与躯体运动功能代表区示意图
用微电极来研究皮层3-21区细胞的体表感觉定位投射,还观察到各类感觉传入的投射也有一定的规律。中央沟底部前壁的3a区是运动区和体表感觉区的移行部分,是肌肉牵张感觉的投射区;3区主要是慢适应感觉的投射区;1区主要是快适应感觉的投射区;2区是关节、骨膜、筋膜等感觉的投射区。因此,中央后回从前到后,分别接受不同的躯体感觉投射;中央后回从上到下,分别接受不同躯体部位的投射。
中央后回是第一感觉区所在部位,在人脑中央前回与岛叶之间还有第二感觉区。第二感觉区面积远比第一感觉区小,区内的投射也有一定的分布安排,安排属于正立而不倒置。刺激人脑第二感觉区可以引致体表一定部位产生主观上麻木感,这种感觉具有双侧性;但人类切除第二感觉区后,并不产生显著的感觉障碍。有人认为,第二感觉区与痛觉有较密切的关系,它可能接受痛觉传入的投射。
(三)中央前回的感觉投射
中央前回(4区)是运动区。在较低等的哺乳类动物(如猫、兔等),体表感觉区与运动区基本重合在一起,称为感觉运动区。这区域即是体表感觉和肌肉本体感觉的代表区,又是运动区。在灵长类动物(如猴、猩猩),体表感觉区与运动区逐渐分离,前者位于中央后回,后者位于中央前回,但这种分化也是相对的。在人脑,刺激中央沟周围皮层时发现,产生运动反应的机会有20%发生在中央后回,而80%发生在中央前回,所以总的来说运动区主要是在中央前回,在灵长类动物,关节和肌梭感觉传入可投射到运动区。应该指出,运动区主要接受从小脑和基底神经传来的反馈投射。
(四)内脏感觉
内脏感觉在皮层也有代表区。电生理研究指出,刺激来自内脏的传入神经可以在皮层一定区域内引出电位变化。例如,刺激内脏大神经的快速传入纤维可以在相应的躯水平体表感觉代表区引出皮层诱发电位。人脑电刺激的研究发现,第二感觉区和运动辅助区(Supplementary motor area)都与内脏感觉有关。刺激第二感觉区及其邻近部位会发生味觉、恶心或排便感等,刺激运动辅助区会产生心悸、脸发热感等。此外边缘系统的皮层部位也是内脏感觉的投射区域。
(五)视觉
枕叶皮层是视觉伯投射区域,左侧枕叶皮层接受左眼的颞侧视网膜和右眼的鼻侧视网膜的传入纤维投射,右侧枕叶皮层接受右眼的颞侧视网膜和左眼的鼻侧视网膜的传入纤维投射。枕叶皮层视觉代表区的具体部位在皮层内侧面的距状裂上下两缘,视网膜上半部投射到距状裂的上缘,下半部投射到下缘;视网膜中央的黄斑区投射到距状裂的后部,视网膜边周区投射到距状裂的前部(图10-27)。电刺激人脑的距状裂上缘(17区),可以使受试者产生简单的主观光感觉,但不能引起完善的视觉形象。对视皮层单个神经元担忧生理研究指出,极少数神经元只对单眼视觉刺激发生反应,这些神经元集中在皮层第四层内,它们接受外膝体投射纤维的传入冲动;绝大多数神经元能对双侧眼球视觉刺激发生反应,这些神经元主要分布在第四层之外的层次中,它们与双眼视觉和立体视觉功能有关。第四层中对单眼视觉刺激发生反应的神经元的感受野通常呈带状,一条光带刺激的朝向如果与该神经元的感受里的朝向一致时,则能够诱发它的最强反应;光带刺激的朝向如果与感受野的朝向垂直时,则几乎不能诱发反应。第四层以外对双眼视觉刺激发生反应的神经元的感受野也呈带状,它们对特定朝向的光带刺激有最佳的反应,并对光带运动的刺激相当敏感,即当特定朝向的光带刺激向一个方向运动时可以诱发很强的反应,而反向的运动引起的反应经弱得多。
图10-27 视网膜各部分投射到大脑皮层枕叶
(六)听觉
颞叶皮层的一定区域中听觉的投射区域,听觉的投射是双侧性的,即一侧皮层代表区与双侧耳蜗感受功能有关。在猕猴,听皮层在颞叶,位于脑岛和颞上回之间;而且耳蜗不同部位的感觉传入冲动投射到听皮层的一定部位,耳蜗底部(高频声感)投射到前部,耳蜗顶部(低频声感)投射到后部,说明不同音频感觉的投射有一定的分野。有人,听觉皮层代表区位于颞横回和颞上回(41、42区),电刺激上述区域能引致受试者产生铃声或吹风样的主观感觉。
(七)嗅觉和味觉
目前知道,嗅觉在大脑皮层的投射区随着进化而愈益缩小,在高等动物只有边缘叶的前底部区域与嗅觉功能有关(包括梨状区皮层的前部、杏仁核的一部分等)。在人脑的刺激研究中观察到,刺激这些相应的结构可以引致特殊的主观嗅觉,如焦橡胶气味等。此外,味觉投射区在中央后回头面部感觉投射区之下侧。
五、痛觉的病理生理
机体受到伤害性刺激时,往往产生痛觉。痛觉是一咱复杂的感觉,常伴有不愉快的情绪活动和防卫反应,这对于保护机体是重要的。疼痛又常是许多疾病的一种症状,因此在临床上引起很大注意。
(一)皮肤痛觉与传导通路
伤害性刺激作用于皮肤时,可先后出现两种性质不同的痛觉,即快痛和慢痛。快痛是一种尖锐而定位清楚的“刺痛”;它在刺激时很快发生,撤除刺激后很快消失。慢痛是一种定位不明确的“烧灼痛”;它在刺激后过0.5-1.0s才能被感觉到,痛感强烈而难以忍受,撤除刺激后还持续几秒钟,并伴有情绪反应及心血管和呼吸等方面的变化。
一般认为痛觉的感受器是游离神经末梢。引起痛觉不需要特殊的适宜刺激,任何形式的刺激只要达到一定强度有可能或已造成组织损伤时,都能引起痛觉,但其机制还不清楚。有人认为,这种游离神经末梢是一种化学感受器,当各种伤害性刺激作用时首先引致组织内释放某引起致痛物质(例如K+、H+组胺、5-羟色胺、缓激肽、前列腺素等),然后作用于游离神经末梢产生痛觉传入冲动,进入中枢引起痛觉。
疼痛的二重性质说明在痛觉伟存在着不同传导速度的神经纤维。实验证明,传导快痛的外周神经纤维主要是有髓鞘的Aδ类纤维,其兴奋阈较低;传导慢痛的外周神经纤维主要是无髓鞘的C类纤维,其兴奋阈较高。
痛觉的中枢传导通路比较复杂。前文已述及,痛觉传入纤维进入脊髓后,在后角更换神经元并发出纤维交叉到双侧,再经脊髓丘脑侧束上行抵达丘脑的体感觉核,转而向皮层体表感觉区投射。此外,痛觉传入冲动还在脊髓内弥散上行,沿脊髓网状纤维、脊髓中脑纤维和脊髓丘脑内侧部纤维,抵达脑干网大辩论结构、丘脑内侧部和边缘系统,引起痛的情绪反应。
(二)内脏痛的特征与牵涉痛
内脏痛是临床常见的症状。内脏痛有与皮肤痛相比较有下列特征:①缓慢、持续、定位不清楚和对刺激的分辨能力差。例如,腹痛时常不易明确分清疼痛发生的部位。②能使皮肤致痛的刺激(切割、烧灼等),作用于内脏一般不产生疼痛;而机械性牵拉、缺血、痉挛和炎症等刺激作用于内脏,则能产生疼痛。例如,内脏器官发生管道梗阻而出现异常运动、循环障碍、炎症时,往往使内脏的感觉上升致意意识并引起剧烈的疼痛。和躯体痛一样,内脏痛也可能是某些致痛物质作用于痛觉感受器引起的;例如,HCI是导致溃疡痛的主要因素,政治家5-羟色胺、组胺和缓激肽等也与溃疡痛有关。
内脏痛的传入神经主要是交感神经干内的传入纤维;它通过后根进入脊髓,然后和躯体神经基本上走着同一上行途径。但食管、气管的痛觉是通过迷走神经干内的传入纤维进入中枢而上传的;部分盆腔器官(如直肠、膀胱三角区、前列腺、子宫颈等)的痛觉传入神经纤维是沿盆神经进入骶髓的(图10-28)。
图10-28 内脏痛觉的神经支配
位于胸痛觉线和骨盆痛觉线之间的器官,其痛觉传入纤维通过交感神经;
在胸痛觉线以上和骨盆痛觉以上的器官,其痛觉传入纤维通过副交感神经
上述的内脏痛是指内脏本身受到刺激时所产生的疼痛,还有一种内脏痛是由于体腔壁浆受到刺激时产生的疼痛,称为体腔壁痛(parietal pain)。例如,胸膜或腹膜受到炎症、压力、磨擦或牵拉等刺激时,也会产生疼痛。这种疼痛与躯体痛相类似,也是由躯体神经(膈神经、肋间神经和腰上部脊神经)传入的。
内脏疾病往往引起身体远隔的体表部位发生疼痛或痛觉过敏,这种现象称为牵涉痛。例如,心肌缺血时,可发生心前区、左肩和左上臂的疼痛;胆囊病变时,右肩区会出现疼痛;阑尾炎时,常感上腹部或脐区有疼痛(表10-4)。发生牵涉痛的部位与真正发生痛觉的患病内脏部位有一定的解剖关系;它们都受同一脊髓节段的后根神经所支配,即患病内脏的传入神经纤维和被牵涉皮肤部位的传入神经纤维由同一后根进入脊髓。因此可以设想由某一内脏传入的神经和由某一皮肤区域传入的神经是在脊髓灰质内同一区域替换神经元的,亦即它们的脊髓中枢是同区域的。假如这两个中枢甚为接近,则由患病内脏传来的冲动将会提高相应的脊髓中枢的兴奋性。从而也影响邻近的中枢,以致由皮肤传入的冲动能使相应的脊髓中枢发生更大的兴奋,由此上传的冲动也可能增强,这可能是痛觉过敏的原因。假如,由患病内脏和内皮肤区域进入脊髓的神经末梢投射到同一脊髓神经元,由同一上行纤维传入脑,则在人日常生活中经常能意识到的是来自皮肤的刺激,因此此时的痛觉传入冲动虽然发源于患病内脏,但仍认为系来自皮肤。这可能是牵涉痛的原因(图10-24)。
表10-4 常见内脏疾病牵涉痛的部位和压痛区
| 患病器官 | 心 | 胃、胰 | 肝、胆囊 | 肾结石 | 阑尾炎 |
| 体表疼痛部位 | 心前区 左臂尺侧 |
左上腹 肩胛间 |
右肩胛 | 腹股沟区 | 上腹部或脐区 |
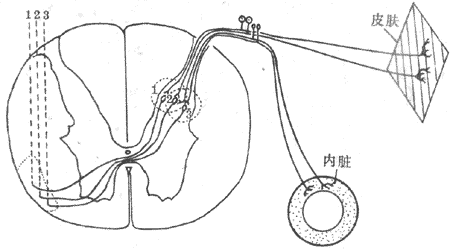
图10-29 牵涉痛产生机制示意图
1:传导体表感觉的后角细胞 2:传导体表和内脏感觉共用的后角细胞
3:传导内脏感觉的后角细胞
第四节 神经系统对躯体运动的调节
一、脊髓对躯体运动的调节
在脊髓的前角中,存在大量运动神经元(α和γ运动神经元),它们的轴突经前根离开脊髓后直达所支配的肌肉。α运动神经元的大小不等,胞体直径从几十到150μm;大α运动神经元支配快肌纤维,小α运动神经地支配慢纤维。α运动神经元接受来自皮肤、肌肉和关节等外周传入的信息,也接受从脑干到大脑皮层等主位中枢传的信息,产生一定的反射传出冲动。因此,α运动神经元是躯干骨骼肌运动反射的最后公路。
α运动神经元的轴突在离开脊髓走和肌肉时,其末梢在肌肉中分成许多小支,每一小支支配一根骨骼肌纤维。因此,在正常情况下,当这一神经元发生兴奋时,兴奋可传导到受它支配的许多肌纤维,引起其收缩。由一个α运动神经元及其支配的全部肌纤维所组成的功能单位,称为运动单位。运动单位的大小,决定于神经元轴突开梢分支数目的多少,一般是肌肉愈大,运动单位也愈大。例如,一个眼外肌运动神经元只支配6-12根肌纤维,而一个四肢肌(如三角肌)的运动神经元所支配的肌纤维数目可达2000根。前者有利于肌肉进行精细的运动,后者有利于产生巨大的肌张力。同一个运动单位的肌纤维,可以和其他运动单位的肌纤维交叉分布,使其所占有的空间范围比该单位肌纤维截而的总和大10-30倍;因此,即使只有少数运动神经元活动,在肌肉中产生的张力也是均匀的。
γ运动神经元的胞体分散在α运动神经元之间,其胞体较α运动神经元为小。γ运动神经元的轴突也经前根离开脊髓,支配骨骼肌骨的梭内肌纤维。据观察,前根中神经纤维的三分之一来自γ运动神经元。γ运动神经元的兴奋性较高,常以较主频率持续放电。在安静和麻醉的运动中都观察到,即使α运动神经元无放电,一些γ运动神经元仍持续放电。γ运动神经元和α运动神经元一样,末梢也是释放乙酰胆碱作为递质的。在一般情况下,当α运动神经元活动增加时,γ运动神经元也相应增加,从而调节着肌梭对牵拉刺激的敏感性。
(一)脊休克
在动物中将脊髓与延髓的联系切断,用以研究脊髓单独的功能;但为了保持动物的呼吸功能,常在颈脊髓第五节水平以下切断,以保留膈神经对膈肌呼吸的传出支配。这种脊髓与高位中枢离断的动物称为脊动物。在脊动物可以观察到脊髓的一些基本功能;但由于失去了高位中枢的调节,因而不能完全反映正常的脊髓功能。
与高位中枢离断的脊髓,在手术后暂时丧失反射活动的能力,进入无反应状态,这种现象称为脊休克(spinal shock)。脊休克的主要表现为:在横断面以下的脊髓所支配的骨骼肌紧张性减低甚至消失,血压下降,外周血管扩张,发汁反射不出现,坡肠和膀胱中粪尿积聚,说明动物躯体与内脏反射活动均减退以至消失。脊休克现象只发生在切断水平以下的部分。以后,一些以脊髓为中枢的反射活动可以逐渐恢复,恢复的迅速与否,与动物种类有密切关系;低等动物如蛙在脊髓离断后数分钟内反射即恢复,在犬则需几天,而在人类则需数周以至数月(人类由于外伤等原因也可出现脊休克)。显然,反射恢复的速度与不同动物脊髓反射依赖于高位中枢的程度有关。反射恢复过程中,首先是一引起比较简单、比较原始的反射先恢复,如屈肌反射、腱反射等;然后才是比较复杂的反射逐渐恢复,如对侧伸肌反射、搔爬反射等。反射恢复后的动物,血压也逐渐上升到一定水平,动物可具有一定的排粪与排快活反射,说明内脏反射活动也能部分地恢复。反射恢复后,有些反射反应比正常时加强并广泛扩散,例如屈肌反射、发汗反射等。
脊休克的产生并不由于切断损伤的刺激性影响引起的,因为反射恢复后进行第二次脊髓切断损务并不能使脊休克重现。所以,疹休克的产生原因是由于离断的脊髓突然失去了高位中枢的调节,这里主要指大脑皮层、前庭核和脑干网状结构的下行纤维对脊髓的易化作用。
脊休克的产生与恢复,说明脊髓可以完成某些简单的反射活动,但正常时它们是在高位中枢调节下进行活动的。高位中枢对脊髓反射既有易化作用的一面,也有抑制作用的一面。例如,切断脊髓后伸肌反射往往减弱,说明高位中枢对脊髓伸肌反射中枢有易化作用;而发汗反射加强,又说明高位中枢对脊髓发汗反射中枢有抑制作用。
脊动物由于脊髓离断,脊髓内上行与下行的神经束秀难重新接通,造成感觉传入冲动不能上达大脑皮层,而大脑皮层的传出冲动也不能下达脊髓,以致在离断水平以上就失去了知觉和所谓随意动作。中枢的神经轴突与外周的神经轴突都有再生能力,但由于脊髓离断部位有大量胶质细胞浸润并逐步形成瘢痕,使轴突再生受阻。
脊髓离断后屈肌反射比正常时加强,而伸肌反射往往减弱,以致屈肌反射常占优势,这不利于瘫痪肢体支持体重。因此,在低位脊髓横贯性损伤的病人,通过站立姿势的积极锻炼以发展伸肌反射是很重要的;这种锻炼使下肢伸肌具有足够的紧张性以保持伸直,以使不依靠拐杖站立或行走。同时,通过锻炼充分发挥末瘫痪肌肉的功能,例如背阔肌等由脊髓离断水平以上的神经所支配,而却随着于骨盆,这样就有可能使病人在借拐杖行走时摆动骨盆。
(二)屈肌反射与对侧伸肌反射
在脊动物的皮肤接受伤害性刺激时,受刺激一侧的肢体出现屈曲的反应,关节的屈肌收缩而伸肌驰缓,称为屈肌反射。屈肌反射具有保持性意义。屈肌反射的强度也刺激强度有关,例如足部的较弱刺激只引致踝关节屈曲,刺激强度加大,则膝关节及髋关节也可发生屈曲。如刺激强度更大,则可以同侧肢体发生屈肌反向的基础上出现对侧肢体伸直的反射活动,称为对侧伸肌反射。对侧伸肌反射是姿势反射的之一,具有维持姿势的生理意义,动物一侧肢体屈曲,对侧肢体伸直以支持体重。屈肌反射是一种多突触反射,其反射弧传出部分可通向许多关节的肌肉。
在人类由于锥体束或大脑皮层运动区的功能发生障碍,脊髓失去了运动区的调节,可出现一处特殊的反射。例如,以钝物划足跖外侧时,出现大趾背屈,其他四趾向外展开如扇形的反射,称为巴彬斯基征(Babinski’s sign)阳性。从生理学角度来看,这一反射属于屈肌反射,因为当刺激加强时还可伴有踝、膝、髋关节的屈曲。平时脊髓在大脑皮层的调节下,这一原始的屈肌反射被抑制而不表现出来。在婴儿的锥体束未发育完全以前,以及成人深睡或麻醉状态下,也可以出现巴彬斯基征阳性。
(三)牵张反射
有神经支配的骨骼肌,如受到外力牵拉使其伸长时,能产生反射效应,引起受牵扯的同一肌肉收缩,此称为牵张反射(stretch reflex)。牵张反射有两种类型,一种为腱反射,也称位相性牵张反射;另一种为肌紧张,也称紧张性牵张反射。
腱反射是指快速牵拉肌腱时发生的牵张反射。例如,叩击膝关节下的股四头肌腱使之受到牵扯,则股四头肌即发生一次收缩,这称为膝反射;叩击跟腱使之受到牵扯,则小腿腓肠肌即发生一次收缩,这称为跟腱反射。这些腱反射的感受器为肌梭,传入神经纤维的直径较粗(12-20μm)、传导速度较快(90m/s以上),效应器为同一肌肉的肌纤维;反射反应的潜伏期很短,据测算兴奋通过中枢的传布时间公0.7ms左右,只够一次突触接替的中枢延搁时间。因此,腱反射为单突触反射,传入神经纤维经背根进入脊髓灰质后,直达前角与运动神经元发生突触联系。当叩击肌腱时,肌肉内的肌梭同时受到牵张,同时发动牵张反射。因此肌肉的收缩几乎是一次同步性收缩。腱反射主要发生于肌肉内收缩较快的快肌纤维成分。
肌紧张是指缓慢持续牵拉腱时发生的牵张反射,其表现为受牵拉拢肌肉能发生紧张性收缩,阻止被拉长。肌紧张是维持躯体姿势最基本的反射活动,是姿势反向的基础。例如,由于重力影响,支持体重的关节趋向于被重力所弯曲,关节弯曲必使伸肌肌腱受到持续牵拉,从而产生牵张反射引起该肌的收缩,对抗关节的屈曲,维持站立姿势。由于重力经常作用于关节,因此这种牵张反射也就持续着。肌紧张与腱反射的反射弧基本相似,感受器也是肌梭,但中枢的突触接替可能不止一个,即可能是多突触反射,其效应器主要是肌肉内收缩较慢的慢肌纤维成分。肌紧张的反射收缩力量并不大,只是抵抗肌肉被牵拉,因此不表现明显的动作。这可能是因为在同肌肉内的不同运动单位进行交替性的收缩而不是同步性收缩,所以肌紧张能持久维持而不易疲劳。
牵张反射主要是使受牵拉的肌肉发生收缩,但同一关节的协同肌也能发生兴奋,而同一关节的颉颃肌则受到抑制(交互抑制),但并不影响其它关节的肌肉,伸肌和屈肌都有牵张反射,但脊髓的牵张反射主要表现在伸肌。屈肌的牵张反射表现不明显,主要表现为它的颉颃肌(即伸肌)受到抑制。牵张反射(尤其是肌紧张)的主要生理意义在于维持这种站立姿势,因此伸肌比屈肌的牵张反射明显符合生理情况。
牵张反射的基本反射弧是简单的,但在整体内牵张反射是受高位中枢调节的,而且可以建立条件反射。腱反射的减弱或消失,常提示反射弧的传入、传出通路受脊髓反射中枢的损害或中断;而腱反射的亢进,则常提示高位中枢的病变I(例如锥体束综合症)。因此,临床上常用测定腱反射的方法来了解神经系统的功能状态。常用的腱反射见表10-5。
表10-5 常用的腱反射
| 名称 | 检查方法 | 中枢部位 | 效应 | |
| 肘反射 | 扣击肱二头肌肌腱 | 颈5-7 | 肘部屈曲 | |
| 膝反射 | 扣击髌韧带 | 腰2-4 | 小腿伸直 | |
| 跟腱反射 | 扣击跟腱 | 腰5-骶2 | 脚向足底方向屈曲 | |
肌梭和腱器官
肌梭是一种感受肌肉长度变化或感受牵拉刺激的特殊的梭形感受装置,长几个毫米,外层为一结谛组织囊(图10-30)。肌梭囊内一般含有6-12根肌纤维,称为梭内肌纤维;而囊外的一般肌纤维就称为梭外肌纤维。整个肌梭现阶附着在梭外肌纤维上,并与其平等排列呈并联关系。梭内肌纤维的收缩成分位于纤维的两端,而感受装置位于其中间部,两者呈串联关系。因此,当梭外肌纤维收缩时,感受装置所受的牵拉刺激将减少;而当梭内肌纤维收缩时,则感受装置对牵拉刺激的敏感度增高。肌梭的传入神经支配有两类。I类传入纤维直径较粗(12-20μm),Ⅱ类传入纤维直径较细(4-12μm)。中枢有运动传出支配梭外肌和梭内肌纤维,前者称为α传出纤维(直径12-20μm),后者称为γ传出纤维(直径2-6μm)。当γ传出纤维活动加强时,梭内肌纤维收缩,可提高肌梭内感受装置的敏感性,因此γ传出纤维的活动对调节牵张反射具有重要作用。进一步研究指出,梭内肌纤维分两类:一类其细胞核集中于中央部称为核袋纤维(nuclear bag fiber),它接受γ1传出纤维支配,并对快速牵拉较敏感,其传入纤维主要属I类;另一类其细胞核分散于整个纤维称为核链纤维(nuclearchain fiber),它接受γ2传出纤维支配,并对缓慢持续牵拉较敏感,其传入纤维有I类和Ⅱ类(图10-31)。
图10-31 两类梭内肌纤维示意图
A:核袋纤维 B:核链纤维
■[此处缺少一些内容]■
拉;当牵拉力量进一步加大时,则可兴奋腱器官使牵张反射受抑制,以避免被牵拉的肌肉受到损伤。
(四)节间反射
脊动物在反射恢复的后期,可出现复杂的节间反射。例如,刺激动物腰背皮肤,可引致后肢发生一系列节奏性搔爬动作,称为搔爬反射。搔爬反向依靠脊髓上下节段的协同活动,所以是节间反射的一种表现。
二、低位脑干肌紧张的调节
(一)去大脑僵直
在中脑上、下叠体(上、下丘)之间切断脑干的动物,称为去大脑动物。去大脑动物由于脊髓与低位脑干相连接,因此不出现脊休克现象,很多躯体和内脏的反射活动可以完成,血压不下降;而在肌紧张活动方面反而出现亢进现象,动物四肢伸直,头尾昂起,脊柱挺硬,称为去大脑僵直(decerbrate rigidity)。去大脑僵直主要是伸肌(抗重力肌)紧张性亢进,四肢坚硬如柱(图10-32)。
图10-32 去大脑僵直
在去大脑动物,如以局部麻醉药注入一肌肉中,或切断相应的脊髓背根,以消除肌梭传入冲动进入中枢,则该肌的僵直现象被消失。可见,去大脑僵直是在脊髓牵张反射的基础上发展起来的,是一种增强的牵张反射。
有人用电刺激动物脑干网状结构的不同区域,观察到在网状结构中具有抑制肌紧张及肌运动的区域,称为抑制区;还有加强肌紧张及肌运动的区域,称为易化区。抑制区位于延髓网状结构的腹内侧部分,电刺激抑制区可引致去大脑僵直减退。易化区分布于广大的脑干中央区域,包括延髓网状结构的背外侧部分、脑桥的中央灰质及被盖;此外下丘脑和丘脑中线核群等部位也具有对肌紧张和肌运动的易化作用,因此也包括在易化区概念之中(图10-33)。从活动的强度来看,易化区的活动比较强,抑制区的活动比较弱;因此在肌紧张的平衡调节中,易化区略占优势。
图10-33猫脑各部位,特别是脑干网状结构下行抑制(一)和易化(+)系统示意图抑制作用(一)的路径:4为网状结构抑制区,发放下行冲动抑制脊髓牵张反射。
这一区接受大脑皮层 (1)尾状核(2)和小脑(3)传来的兴奋。
易化作用(+)的路径:5为网状结构易化区,发放下行冲动加强脊髓牵张反射。
6为延髓的前庭核,有加强脊髓牵张反射的作用。
目前知道,抑制肌紧张的中枢部位有大脑皮层运动区、纹状体、小脑前叶蚓部、延髓网状结构抑制区;易化肌紧张的中枢部位有前庭核、小脑前叶两侧部、网状结构易化区。这些结构有的在脑干外,但与脑干内部的有关功能结构有功能上的联系。例如,刺激小脑前叶蚓部,可以在网状结构抑制区获得诱发电位,因引小脑前叶蚓部的作用可能是通过网状结构抑制区来完成的;又如,大脑皮层运动区和纹状体的作用可能也是通过网状结构抑制区来完成的。这些脑干外的抑制肌紧张的区域,不仅通过加强网状结构抑制区活动,使肌紧张受到抑制;而且也能控制网状结构易化区,使易化区的活动受到压抑,转而使肌紧张减退。再如,前庭核接受内耳前庭器官传入冲动的作用,转而提高网状结构易化区的活动;小脑前叶两侧部的肌紧张易化作用,可能也是通过网状结构易化区来完成的。在去大脑动物中,由于切断了大脑皮层运动区和纹状体等部位与网状结构的功能联系,造成抑制区活动减弱而易化区活动增强,使易化区的活动占有明显的优势,以致肌紧张过度增强而出现去大脑僵直。
去大脑僵直主要是抗重力肌的肌紧张明显加强。一般情况下伸肌是抗重力肌,因此伸肌肌紧张在去大脑僵直时明显加强。有的动物,如南美洲的树赖(Sloth)栖于森林中,经常悬挂在树上,屈肌是抗重力肌;这类动物发生去大脑僵直时,屈肌的紧张明显加强。人类在某些疾病中,也可出现与动物去大脑僵直相类似的现象。例如,蝶鞍上囊肿引致皮层与皮层下失去联系时,可出现下肢明显的伸肌僵直及上肢的半屈状态,称为去皮层僵直(decorticate rigidity)。上肢的半屈状态是抗重力肌肌紧张增强的表现,因为人是直立的动物。人类的去大脑僵直,有时可在中脑具有疾患时出现,表现头后低仰,上下肢僵硬伸直,上臂内旋,手指屈曲(图10-34)。临床上如见到患者出现去大脑僵直现象,往往表明病变已严重地侵犯了脑干,预后不良的信号。
图10-34 人类去皮层僵直及去大脑僵直
A,B,C去皮层僵直 A:仰卧,头部姿势正常时,上肢半屈
B和C:转动头部时,上肢姿势 D:去大脑僵直,上下肢均伸直
(二)α僵直和γ僵直
从牵张反射的角度来分析,肌紧张加强的机制可以有二种。一种是于高位一中枢的下行性作用,直接或间接通过脊髓中间神经元提高α运动神经元的活动,从而导致肌紧张加强而出现僵直,这称为α僵直。另一种是由于高位中枢的下行性作用,首先提高脊髓γ运动神经元的活动,使肌梭的敏感性提高而传入冲动加多,转而使脊髓α运动神经元的活动提高,从而导致肌紧张加强而出现僵直,这称为γ僵直(图10-35)。由前庭核下行的作用主要是直接或间接促使α运动神经元活动加强,导致α僵直;由网状结构易化区下行的作用主要使γ运动神经元活动提高,转而发生肌紧张加强,出现γ僵直。经典的去大脑僵直主要属于γ僵直,因为在消除肌梭传入冲动对中枢的作用后,僵直现象可以消失。
图10-35高级中枢对骨骼肌运动控制的模式图
三、姿势反射
中枢神经系统调节骨骼肌的肌紧张或产生相应的运动,以保持或改正身体空间的姿势,这种反射活动总称为姿势反射。前述的牵张反射、对侧伸肌反射就是最简单的姿势反射。此外还有比较复杂的姿势反射,例如状态反射、翻正反射、直线或旋转加速运动反射(见感觉器官章)等。
(一)状态反射
头部在空间的位置改变以及头部与躯干的相对位置改变时,可以反射性地改变躯体肌肉的紧张性,这种反射称为状态反射。状态反射包括迷路紧张反射与颈紧张反射两部分。迷路紧张反射是指内耳迷路的椭圆囊和球囊的传入冲动对躯体伸肌紧张性的调节反射。在去大脑动物实验中见到,当动物仰臣卧时则伸肌紧张性最高,而当动物俯臣卧时则伸肌紧张性最低。这是由于不同头部位置会引致内耳迷路不同刺激的结果而造成的。颈紧张反射是指颈部扭曲时,颈椎关节韧带或肌肉受刺激后,对四肢肌肉紧张性的调节反射。实验证明,头向一侧扭转时,下颏所指一侧的伸肌紧张性加强;如头后仰时,则前肢伸肌紧张性加强,而后肢伸肌紧张性降低;如头前俯时,则后肢伸紧张性加强,而前肢伸肌紧张性降低,人类在去皮层僵直的基础上,也可出现颈紧张反射;即当颈扭曲时,下颏所指一侧上肢伸直,而对侧上肢则处于更屈曲状态(图10-34)。在正常人体中,由于高级中枢的存在,状态反射常被抑制不易表现出来。
(二)翻正反射
正常动物可保持六立姿势,如将其推倒则可翻正过来,这种反射称为翻正反射。如将动物四足朝天从空中掉下,则可清楚地观察到在下坠过程中,首先是头颈扭转,然后前肢和躯干跟着也扭转过来,最后后肢也扭转过来,当下坠到地面时由四足着地。这一翻正反射包括一系列反射活动,最先是由于头部位置不正常,视觉与内耳迷路感受刺激,从而引起头部的位置翻正。头部翻正以后,头与躯干的位置关系不正常,使颈部关节韧带或肌肉受到刺激,从而使躯干的位置也翻正。
四、小脑
小脑对于维持姿势、调节肌紧张、协调随意运动均有重要的作用。根据小脑的传入、传出纤维的联系,可以将小脑划分为三个主要的功能部分,即前庭小脑、脊髓小脑和皮层小脑(图10-36)
图10-36 灵长类动物小脑分叶平展示意图
(一)前庭小脑
前庭小脑主要由绒球小结叶构成,与身体平衡功能有密切关系。运动切除绒球小结叶后则平衡失调。实验观察到,切除绒球小结叶的猴,由于平衡功能失调而不能站立,只能躲在墙角里依靠墙壁而站立;但其随意运动仍然很协调,能很好地完成吃食动作。在第四脑室附近出现肿瘤的患者,由于肿瘤往往压迫损伤绒球小结叶,患者站立不稳,但其肌肉运动协调仍良好。绒球小结叶的平衡功能与前庭器官及前庭核活动有密切关系,其反射进行的途径为:前庭器官→前庭核→绒球小结叶→前庭核→脊髓运动神经元→肌肉装置。在动物实验中还观察到,犬切除小结叶后,则运动病不再发生;猫切除小结叶后,可出现位置性眼震颤(positional nystagmus),当头位固定于特定位置时眼震颤即出现。以上均说明绒球小结叶对调节前庭核的活动有重要作用。
(二)脊髓小脑
脊髓小脑由小脑前叶(包括单小叶)和后叶的中间带区(旁中央小叶)构成。这部分小脑主要接受脊髓小脑传入纤维的投身投射,其感觉传入冲动主要来自肌肉与关节等本体感受器;但是前叶还接受视觉、听觉的传入信息,而后叶的中间带区还接受脑桥纤维的投射。前叶的传出纤维主要在顶核换神经元,转而进入脑干网状结构;后叶中间带区的传出纤维经间置核到红核,有一些纤维再投射到丘脑外侧腹核,最后抵达大脑皮层运动区。
前叶与肌紧张调节有关。在去大脑动物,刺激前叶蚓部可抑制同侧伸肌紧张,使去大脑僵直减退,因此前蚓部有抑制紧张的作用。抑制肌紧张的区域在前叶蚓部有一定的空间分布,前叶明目张胆部之前端与动物尾部部及下肢肌紧张的抑制功能有关,前叶蚓部之后端及单小叶部位与上肢及头而部肌紧张的抑制功能有关,其分布安排是倒置的。前叶蚓部抑制肌紧张的作用,可能是通过延髓网状结构抑制区转而改变脊髓前角运动神经元活动的。在猴的实验中观察到,刺激小脑前叶两侧部有加强肌紧张的作用;这些肌紧张易化区也有一定的空间分布。而且安排也是倒置的,其作用可能是通过网状结构易化区转而改变脊髓前角运动神经元活动的。因此,小脑前叶对肌紧张的调节既有抑制又有易化的双重作用。在进化过程中,前叶的肌紧张抑制作用逐渐减弱,而肌紧张的易化作用逐渐占主要地位。
后叶中间带也有控制肌紧张的功能,刺激该区能使双侧肌紧张加强。由于后叶中间带还接受脑桥纤维的投射,并与大脑皮层运动区之间有环路联系,因此它在执行大脑皮层发动的随意运动方面有重要作用。当切除或损伤这部分小脑后,随意动作的力量、方向及限度将发生很大紊乱,同时肌张力减退,表现为四肢乏力。受害动物或患者不能完成精巧动作,肌肉在完成动作时抖动而把握不住动作的方向(称为意向性震颤),行走摇晃呈酩酊蹒跚状,如动作越迅速则协调障碍也越明显。患者不能进行颉颃肌轮替快复动作(例如上臂不断交替进行内旋与外旋),但当静止时则看不出肌肉有异常的运动。因此说明,这部分小脑是对肌肉在运动进行过程中起协调作用的。这种动作性协调障碍,称为小脑共济失调(cerebellar ataxia)。
(三)皮层小脑
皮层小脑指后叶的外侧部,它不接受外周感觉的传入信息,仅接受由大脑皮层广大区域(感觉区、运动区、联络区)传来的信息。这些区域的下传纤维均经脑桥换元,转而投射到对侧的后叶外侧部,后叶外侧部的传出纤维经齿状核换元,再经丘脑外侧腹核换元,然后投射到皮层运动区。皮层小脑与运动区、感觉区、联络区之间的联合活动和运动计划的形成及运动程序的编制有关。精巧运动是逐步在学习过程中形成熟练起来的。在开始学习阶段,大脑皮层通过锥体系所发动的运动不是协调的,这是因为小脑尚未发挥其协调功能。在学习过程中,大脑皮层与小脑之间不断进行着联合活动,同时小脑不断接受感觉传入冲动的信息逐步纠正运动过程中所发生的偏差,使运动逐步协调起来。在这一过程中,皮层小脑参与了运动计划的形成和运动程序的编制。当精巧运动逐渐熟练完善后,皮层小脑中就贮存了一整套程序;当大脑皮层要发动精巧运动时,首先通过下行通路从皮层小脑中提取贮存的程序,并将程序回输到大脑皮层运动区,再通过锥体束发动运动。这时候所发动的运动可以非常协调而精巧,而且动作快速几乎不需要思考。例如,学习打安运动的过程或演奏动作的过程,都是这样一个过程。
(四)小脑内局部神经元回路
小脑皮层各区的组织结构都是相似的。进入小脑皮层的纤维只有攀缘纤维和藓苔纤维两类,两者均起到兴奋作用。小脑皮层内有五类神经元,即颗粒细胞、高尔其细胞、篮状细胞、星状细胞和浦氏细胞;除颗粒细胞为兴奋性神经元外,其余均为抑制性神经元。浦氏细胞的轴突是小脑皮层唯一的传出细胞,它与小脑深部核团(顶核、间置核、齿状核)发生突触联系,抑制核团内神经元(兴奋性神经元)的紧张性放电活动。攀缘纤维主要来自延崩的下橄榄核,进入小脑皮层起到强烈的兴奋作用。藓苔纤维是进入小脑皮层的主要传入纤维,来源很广泛,进入小脑皮层后与颗粒细胞发生突触联系,起着兴奋颗粒细胞的作用。颗粒细胞的轴突进入小脑皮层浅层后形成平行纤维,转而兴奋其他神经元。例如兴奋高尔基细胞,高尔基细胞兴奋后即反馈抑制颗粒细胞的活动。由于高尔基细胞轴突的分布比较广泛,它能同时抑制许多颗粒细胞的活动,造成颗粒细胞的兴奋反应在空间上局限起来。也就是说,由藓苔纤维直接兴奋的颗粒细胞处在兴奋状态,而其外围的颗粒细胞则外在抑制状态。又如平行纤维可兴奋浦肯野细胞、篮状细胞和星状细胞,而篮状细胞和星状细胞能抑制浦肯野细胞和活动;由于篮状细胞的轴突分布较广,浦肯野细胞受抑制的范围较大,造成浦肯野细胞的兴奋反应在空间上局限起来。也就是说,由平行纤维直接兴奋的浦肯野细胞处在兴奋状态,其邻旁的浦步野细胞则处在抑制状态。因此,通过上述局部神经元回路的作用,使许多不同来源的藓苔纤维的冲动进入小脑皮层后,出现许多兴奋与抑制镶嵌的区域,这对于小脑精确地调节不同部位的肌肉肌紧张或协调其随意运动是很重要的(图10-37)。
图10-37 小脑内局部神经元回路示意图
1:藓苔纤维 2:攀缘纤维 3:小脑深部核团细胞4:颗粒细胞 5:高尔基细胞 6:浦肯野细胞
7:篮状细胞 8:星状细胞 9:平行纤维黑色细胞均为抑制性神经元
五、基底神经节
基底神经节包括尾(状)核、壳核、苍白球、丘脑底核、黑质和红核。尾核、壳核和苍白球统称纹状体;其中苍白球是较古老的部分,称为旧纹状体,而尾核和壳核则进化较新,称为新纹状体。尾核、壳核、苍白球与丘脑底核、黑质在结构与功能上是紧密相联系的。其中苍白球是纤维联系的中心,尾核、壳核、丘脑底核、黑质均发出纤维投射到苍白球,而苍白球也发出纤维与丘脑底核、黑质相联系。
基底神经节有重要的运动调节功能,它对随意运动的稳定、肌紧张的控制、本体感觉传入冲动信息的处理都有关系。在清醒猴,记录苍白球单个神经元的放电活动时观察到,当肢体进行随意运动时神经元活动发生明显明确的变化;有的神经元在肢体屈曲时放电增多,说明在底神经节与随意运动有关。电刺激纹状体的动物实验中观察到,单纯刺激纹状体并不能引起运动效应;但如在刺激大脑皮层运动区的同时,再刺激尾核或苍白球,则皮层运动区发出的运动反应即迅速抑制,并在刺激停止后抑制效还可继续存留一定时间。在猴,单侧损毁苍白球后,则对侧上肢的运用就不如同侧上肢的运用那样灵便。以上均说明基底神经节的功能与躯体运动有密切关系,便这些实验事实仍不能说清楚基底神经节是如何调节身体运动的。为一进一步阐明基底神经节的功能,有必要简述基底神经节功能紊乱的疾病。临床上在底神经节损害的主要表现可分为两大类:一类是具有运动过多而肌紧张不全的综合症,另一类是具有运动过少而肌紧张过强的综合症。前者的实例是舞蹈病与手足徐动症等,后者的实例是震颤麻痹(帕金森病)。临床病理的研究指出,舞蹈病与手足徐动症的病变主要位于纹状体,而震颤麻痹的病变主要位于黑质。
震颤麻痹患者的症状是:全射肌紧张增高、肌肉强直、随意运动减少、动作缓慢、面部表情呆板。此外,患者常伴有静止性震颤,此种震颤多见于上肢(尤其是手部),其次是下肢及头部;震颤节律每秒钟约4-6次,静止时出现,情绪激动时增强,进入自主运动时减少,入睡后停止。关于震颤麻痹的产生原因,目前已有较多的了解。近年来,通过对中枢递质的研究,已明确中脑黑质是多巴胺能神经元存在的主要部位,其纤维上行可抵达纹状体(图10-38)。震颤麻痹患者的病理研究证明,其黑质有病变,同时脑内多巴胺会计师明显下降。在动物中,如用药物(利血平)使儿茶酚胺(包括多巴胺)耗竭,则动物会出现类似震颤麻痹的症状;如进一步给予左旋多巴(L-dopa,多巴胺之前体,能通过血脑屏障进入中枢神经系统)治疗,使体内多巴胺合成增加,则症状好转。由此说明,中脑黑质的多巴胺能神经元功能被破坏,是震颤麻痹的主要原因。
图10-38 黑质纹状体环路示意图
1:多巴胺能神经元 2:碱能神经元3:γ-氨基丁酸能神经元
4:纹状体5:黑质致密部6:黑质网状部
早已知道,震颤麻痹患者能用M型胆碱能受体阻断剂(东莨菪碱、安坦)治疗,说明震颤麻痹的产生与乙酰胆碱递质功能的改变也有关系。在震颤麻痹患者进行苍白球破坏手术治疗过程中,如将乙酰胆碱直接注入苍白球,则导致对侧肢体症状加剧,而注入M受体阻断剂则症状减退。由此说明,纹状体内存在乙酰胆碱递质系统,其功能的加强将导致震颤麻痹症状的出现。总结多巴胺递质系统和乙酰胆碱递质系统的不同作用,目前认为黑质上行抵达纹状体的多巴胺递质系统的功能,在于抑制纹状体内乙酰胆碱递质系统的功能;震颤麻痹患者由于多巴胺递质系统功能受损,导致乙酰胆碱递质系统功能的亢进,才出现一系列症状。如果应用左旋多巴以增强多巴胺的合成,或应用M受体阻断剂以阻断乙酰胆碱的作用,均对震颤麻痹有一定的治疗作用。
静止性震颤的发生,可能与丘脑外侧腹核等结构的异常活动有关。用微电极记录震颤麻痹患者丘脑外侧腹核的神经元放电,可以观察到某些神经元具有周期性短串放电,其周期节律与震颤肢体的节律相同步,破坏丘脑这些区域后则静止性震颤消失。有人认为,这种异常活动是神经环路活动的结构结果,其通路可能为:丘脑外侧腹核→大脑皮层运动区→纹状体→丘脑外侧腹核。因为,切断苍白球至丘脑外侧腹核的纤维联系后,也可使静止性震颤消失。
舞蹈病患才的主要临床表现为不自主的上肢和头部的舞蹈样动作,并伴有肌张力降低等。病理研究证明,遗传性舞蹈病患者有显著的纹状体神经元病变,新纹状体严重萎缩,而黑质-纹状体通路是完好的,脑内多巴胺含量一般也正常。在这类患者,若采用左旋多巴进入治疗反而使症状加剧,而用利血平耗竭包括多巴胺在内的神经递质,却可使症状缓解。神经生化的研究发现,患者的纹状体中胆碱能神经元与γ-氨基丁酸能神经元的功能明显减退。因此认为,舞蹈病病变主要是纹状体内的胆碱能和γ-氨基丁酸能神经元功能减退,而黑质多巴胺能神经元功能相对亢进,这和震颤麻痹的病变正好相反。目前知道,黑质和纹状体之间有环路联系的;黑质的多巴胺能神经元的轴突上行抵达纹状体,能控制纹状体内的胆碱能神经元的活动,转而改变纹状体内γ-氨基丁酸能神经元的活动,然后γ-氨基丁酸能神经元的轴突下行抵达黑质,反馈控制多巴胺能神经元的活动(图10-38)。当纹状体内的胆碱能和γ-氨基丁酸能神经元病变时,上述环路功能受损,导致多巴胺能神经元活动亢进。
六、大脑皮层对躯体运动的调节
(一)大脑皮层的主要运动区
大脑皮层的基本些区域与躯体运动功能有比较密切的关系。在灵长类动物,中央前区的4区和6区是控制躯体运动的运动区。运动区有下列的功能特征:①对躯体运动的调节支配具有交叉的性质,即一侧皮层主要支配对侧躯体的肌肉。蛤这种交叉性质不是绝对的,例如头面部肌肉的支配多数是双侧性的,像咀嚼运动、喉运动及脸上运动的肌肉的支配是双侧性的;然而面神经支配的下部面肌及舌下神经支配的舌肌却主要受对侧皮层控制。因此,在一侧内囊损伤后,产生所谓上运动神经元麻痹时,头面部多数肌肉并不完全麻痹,但对侧下部面肌及舌肌发生麻痹。②具有精细的功能定位,即一定部位皮层的刺激引起一定肌肉的收缩。功能代表区的大小与运动的精细复杂程度有关;运动愈精细而复杂的肌肉,其代表区也愈大,手与五指所占的区域几乎与整个下肢所占的区域大小相等。③从运动区的上下分布来看,其定位安排呈身体的倒影;下肢代表区在顶部(膝关节以下肌肉代表区在皮层内侧面),上肢代表区在中间部,头而部肌肉代表区在底部(头面部代表区内部的安排仍为正立而不倒置)。从运动区的前后分布来看,躯干和肢体近端肌肉的代表区在前部(6区),肢体远端肌肉的代表区在后部(4区),手指、足趾、唇和舌的肌肉代表区在中央沟前缘。对正常人脑进行局部血流测定时观察到,足部运动时运动区足部代表区血流增加,手指运动时手部代表区血流增加(图10-26)。
在动物实验中还观察到,电刺激8区可引致眼外肌的运动反应,刺激枕叶18、19区也可获得较为微弱的眼外肌运动反应。此外,在猴与人的大脑皮层,用电刺激法还可以找到运动辅助区。该区在皮层内侧面(两半球纵裂的侧壁)4区之前,刺激该区可以引致肢体运动和发声,反应一般为双侧性。
在大脑皮层运动区的垂直切面上,可以见到该区细胞和前述的皮层感觉区类似,,也呈纵向柱状排列,组成大脑皮层的基本功能单位,称为运动柱(motor columm)。一个运动柱可控制同一关节的几块肌肉的活动,而一个肌肉可接受几个运动柱的控制。
(二)锥体系
皮层的躯体运动调节功能,是通过锥体系和锥体外系下传而完成的。锥体系一般是指由皮层发出经延髓锥体而后下达脊髓的传导系(即锥体系、或称皮层脊髓束);然而由皮层发出抵达脑神经神经运动核的纤维(皮层脑干束),虽不通过延髓锥体,也应包括在锥体系的概念之中。因为,后者与前者在功能上是相似的,两者都是由皮层运动神经元(上运动神经元)下传抵达支配肌肉的下运动神经元(脊髓前角运动神经元和脑神经核运动神经元)的最直接通路。
曾认为锥体束下传的纤维均直接与下运动神经元发生突触联系,但已知有80%-90%的上下运动神经元之间还间隔有一个以上中间神经元的接替。由于皮层4区是躯体运动调节的主要区域,在4区灰质第五层内有大锥体细胞(贝茨细胞),其纤维下传是通过锥体束抵达下运动神经元的,因此很容易误解为锥体束纤维的组成仅来自4区的大锥体细胞。事宜上,每一侧皮层4区大锥体细胞在人类总共约34000个左右,而每一侧锥体束却含有直径大小不等的纤维总数达100万左右,显然不能认为锥体束纤维仅由4区的大锥体细胞发出。在锥体束中直径较为粗大的(11-20μm)有髓纤维约占总数的2%-3%,看来由4区大锥体细胞发出的纤维仅属锥体束内直径粗大的纤维。此外,由4区发出进入锥体束的纤维还有来自该区第三至第六层的小细胞,但是破坏4区后锥体束内发生变性的纤维仅占27%-40%,因此仍然不能认为锥体束纤维仅由4区发出。研究指出,6区、3-1-2区、5区、7区等都有纤维进入锥体束;因为电刺激延髓锥体记录逆行性皮层诱发电位,可在上述区域观察到电位变化。
上文指出,上下运动神经元之间多数存在中间神经元的接替,仅有10%-20%上下运动神经元之间的联系是直接的,亦即属于单突触联系的。电生理研究指出,这种单突触直接联系在前肢运动神经元比后肢运动神经元多,而且在肢体远端肌肉的运动神经元又比近端肌肉的运动神经元多。由此可见,运动愈精细的肌肉,大脑皮层对其运动神经元的支配具有愈多的单突触直接联系。从进化来看,猫和犬没有这种直接的单突触联系;浣熊的前掌指有一定灵巧性,已证明其锥体束有单突触联系;大多数灵长类的锥体束有单突触联系,而以人的单突触联系数量为最大。这种单突触联系可使α运动神经元产生兴奋性突触后电位,并使神经元发出冲动以发动肌肉收缩。锥体束下传冲动也与脊髓前角γ运动神经元有联系,并可激活该运动神经元;但没有证据说明,锥体束下传冲动运动是通过γ运动神经元环路的(指间接通过肌梭传入冲动的增加,来兴奋α运动神经元)。因此,锥体束可分别控制α运动神经元和γ运动神经元的活动,前者在于发动肌肉运动,后者在于调整肌梭的敏感性以配合运动,两者活动协同控制着肌肉的收缩。此外,锥体束下行纤维配脊髓中间神经元也有突触联系,从而改变脊髓颉颃肌运动神经元之间的对抗平衡,使肢体运动具有合适的强度,保持运动的协调性。
(三)锥体外系
上世纪有人认为,皮层下的某些核团(尾核、壳核、苍白球、黑质、红核等)有下行通路控制脊髓的运动神经元活动,由于它们的通路在延髓锥体之外,因此称为锥体外系。经典的锥体外系概念认为这一系统与大脑皮层无关;但是后来发现这些核团不仅直接接受大脑皮层下行纤维的联系,而且还接受锥体束下行纤维侧支的联系,同时还经过丘脑对大脑皮层有上行纤维的联系。为区别于经典的锥体外系概念,由大脑皮层下行通过皮层并通过皮层下核团接替转而控制脊髓运动神经元的传导系统,称为皮层起源的锥全外系(cortically originating extrapyramidal system);由锥体束侧进入皮层下核团转而控制脊髓运动神经元的传导系统,称为旁锥体系(parapyramidal system)(图10-39)。
皮层起源的锥体外系是大脑皮层控制躯体运动的另一下行传导通路。锥体外系的皮层起源比较广泛,几乎包括全部大脑皮层,但主要来自是额叶和顶叶的感觉区、运动区和运动辅助区。因此,皮层的锥体系和锥体外系的起源是相互重迭的。皮层锥体外系的细胞一般属于中、小型锥体细胞,它们的轴突较短,离开大脑皮层后终止于皮层下基底神经节、丘脑、脑桥和延髓的网状结构,通过一次以上神经元的接替,最后经网状脊髓束、顶盖脊髓束、红核脊髓束和前庭脊髓束下达脊髓,控制脊髓的运动神经元。锥体外系对脊髓反射的控制常是双侧性的,其功能主要与调节肌紧张、肌群的协调性运动的关。
图10-39 锥体系和锥体外系示意图
1:大脑皮层 2:皮层下核团 3:延髓锥体 4:脊髓 5:锥体束
6:旁锥体束 7:皮层起源的锥体外系 8:锥体外系
(四)皮层运动区和锥体系功能障碍对运动的影响
由于锥体系和锥体外系在皮层的起源互相重迭的,因此皮层运动区的损伤效应就难于分清是属于锥体系还锥体外系功能缺损。同时,锥体束下行经过脑干时,还发现许多侧支进入皮层下核团调节锥体外系的活动。所以,从皮层到脑干之间,由于种种病理过程产生的运动障碍往往是由于锥体系和锥体外系合并损伤的结果。但是到达延髓尾端水平,锥体束出现相对独立性,延髓锥体的损伤效应可以认为主要是锥体系功能缺损。
单侧切断猫或猴的延髓锥体所造成的结果并不严重;动物仅表现对侧肌张力减退,肢体远端肌肉麻痹(随意运动消失)和腱反射减弱,巴彬斯基阳性,以及动物减少对此肢体的运用和永远失去其敏捷灵巧活动的能力等。若切断双侧延髓锥体,则上述缺损表现在双侧肌肉。可见,锥体束的功能主要是对四肢远端肌肉活动的精细调节。中央前回运动区的损伤,在不同动物表现不一样。猫和犬双侧大脑皮层运动区切除后仍能站立、奔跑;灵长类动物双侧大脑皮层运动区切除后,动物完全麻痹,四肢肌张力亢进。但单侧切除猴的大脑皮层运动区,则功能缺损比双侧切除轻和多,运动能运用其四肢,虽然对侧手指的动作苯拙而不灵巧,但奔跑和站立并无明显困难;说明猴的大脑皮层运动区对肌肉运动的调节虽然以对侧为主,但在失去对侧皮层功能的情况下,可以对双侧的运动进行调节。在人类,单侧是央前回的扣伤则使对侧肢体完全失随意运动的能力,手和脚的肌肉常完全麻痹,关于中央前回运动区损伤后产生痉挛麻痹还是柔软性麻痹,这一问题已争论多年。目前知道,严格的4区损伤出现肢体远端肌肉麻痹,并不产生痉挛,一般是柔软性麻痹;损伤6区后则肢体近端肌肉麻痹并伴有痉挛;若整个中央前回运动区损伤,则肢体全部肌肉麻痹并伴有痉挛,出现痉挛性麻痹。
临床上把涉及锥体束损害的一系列表现称为锥体束综合征(上运动神经元麻痹)。它包括随意运动的丧失,肌紧张加强,腱反射亢进以致出现阵挛,巴彬斯基征阳性,部分浅反射减退或消失等。肌紧张加强或腱反射亢进,都是牵张反射亢进的表现;所谓阵挛也是由于牵张反射过强,以致人工持续牵拉肌腱会反射一系列连续的腱反射。部分浅反射减退或消失,是指腹壁反射(轻划腹皮肤引致壁肌收缩)、提睾反射(轻划肭内侧皮肤引致提睾肌反收缩)等减退或消失,其原因还不完全清楚。一般认为,这类浅反射存在经由大脑皮层的反射通路,以致锥体束损害就使反射弧中断,反射发生障碍;但也有人认为,浅反射减退或消失仅是由于锥体束损伤后脊髓浅反射中枢的兴奋性减退所致。因为有人用肌电图法观察人体腹壁反射,发现其中枢延搁时间很短(3.55-5.4ms),与动物的脊髓多突触反射相当,似乎不存在大脑皮层的反射通路,所谓锥体束综合征实际上是锥体系和锥体外系合并损伤的结果,而不是严格的单纯锥体束传导中断的表现。为此,有些人反对采用传统的锥体系和锥体外系概念,因为这两个系统在功能上和在损伤后功能缺损上无法完全区分。上运动神经元损害了下运动神经元损害的是临床表现是不同的,见表10-6。
表10-6 上、下运动神经元麻痹的区别
| 表现 | 上运动神经元麻痹(硬瘫、弃挛性瘫、中枢性瘫) | 下运动神经元麻痹(软瘫、萎缩性瘫、周围性瘫) |
| 损害部位 | 皮层运动区或锥体束 | 脊髓前角运动神经元或运动神经 |
| 麻痹范围 | 常为广泛的 | 常为局限的 |
| 肌紧张 | 张力过强、痉挛 | 张力减退、松驰 |
| 腱反射 | 增强 | 减弱或消失 |
| 浅反射 | 减弱或消失 | 减弱或消失 |
| 病理反射 | 巴彬斯基征阳性 | 无 |
| 肌萎缩 | 不明显 | 明显(肌肉失去了神经的营养性作用) |
第五节 神经系统对内脏活动的调节
一、自主神经系统
自主神经系统是指调节内脏功能的神经装置,也可称为植物性神经系统或内脏神经系统。实际上,自主神经系统还是接受中枢神经系统的控制的,并不是完全独立自主的。按一般惯例,自主神经系统仅指支配内脏器官的传出神经,而不包括传入神经;并将其分成交感神经和副交感神经两部分。(图10-40)
图10-40 自主神经分布示意图
(一)交感和副交感神经的特征
从中枢发出的自主神经在抵达效应器官前必须先进入外周神经节(肾上腺髓质的交感神经支配是一个例外),此纤维终止于节仙神经元上,由节内神经元再发现纤维支配效应器官。由中枢发出的纤维称为节前纤维,由节内神经元发出的纤维称为节后纤维。节前纤维性有髓鞘B类神经纤维,传导速度较快;节后纤维性无髓鞘C类神经纤维,传导速度较慢。交感神经节离效应器官较远,因此节前纤维短而节后纤维长;副交感神经节离效应器官较近,有的神经节就在效应器官壁内,因此节前纤维长而节后纤维短。
交感神经起自脊髓胸腰段的外侧柱。副交感神经的起源比较分解,其一部分起正脑干的缩瞳核、上唾液核、下唾液核、迷走背核、疑核,另一部分起自脊髓骶部相当于侧角的部位。交感神经的全身分布广泛,几乎所有内脏器官都受它支配;而副交感神经的会布较局限,某些器官不具有副交感神经支配。例如,皮肤和肌肉内的血管、一般的汗腺、竖毛肌、肾上腺髓质、肾就只有交感神经支配。
刺激交感神经的节前纤维,反应比较弥散;刺激副交感神经的节前纤维,反应比较局限,因为一根交感节前纤维往往和多个节内神经元发生突触联系,而副交感神经则不同。例如,猫颈上神经节内的交感节前与节后纤维之比为1:11-17,睫状神经节内的副交感节前与节后纤维之比为1:2。
通过荧光组织化学的研究,发现哺乳动物中,交感神经节后纤维交不都是支配效应器官细胞的。在心脏和膀胱中,少量交感神经节后纤维支配器官壁内的神经节细胞;在胃和小肠中,多数交感神经节后纤维支配器官壁内的神经节细胞。由此看来,交感和副交感神经的相互作用,可以发生在器官壁内神经节细胞水平上,而不一定发生在效应器官细胞水平上。
(二)交感和副交感神经系统的功能
自主神经系统的功能在于调节心肌、平滑肌和腺体(消化腺、汗腺、部分内分泌腺)的活动(表10-7)。除少数器官外,一般组织器官都接受交感和副交感的双重支配。在具有双重支配的器官中,交感和副交感神经的作用往往具有拮抗的性质。例如,对于心脏,迷走神经具有抑制作用,而交感神经具有兴奋作用;对于小肠平滑肌,迷走神经具有增强其运动的作用,而交感神经却具有抑制作用。这种拮抗性使神经系统能够从正反两个方面调节内脏的活动,拮抗作用的对立统一是神经系统对内脏活动调节的特点。在一般情况下,交感神经中枢的活动和副交感神经中枢的活动是对立的,也就是说当交感神经系统活动相对加强时,副交感神经系统活动就处于相对减退的地位,而在外周作用方面却表现协调一致。但是,在某些情况下,也可出现交感和副交感神经系统活动都增强或都减退,然而两者间必有一个占优势。在某些外周效应器上,交感和副交感神经的作用是一致的,例如唾液腺的交感神经和副交感神经支配都有促进分泌的作用;但两者的作用也有差别,前者的分泌粘稠,后者的分泌稀薄。
表10-7 自主神经的主要功能
| 器官 | 交感神经 | 副交感神经 |
| 循环器官 | 心跳加快加强 腹腔内脏血管、皮肤血管以及分布于唾液腺与外生殖器官的血管均收缩,脾包囊收缩,肌肉血管可收缩(肾上腺素能)或舒张(胆碱能) |
心跳减慢,心房收缩减弱 部分血管(如软脑膜动脉与分布于外生殖器的血管等)舒张 |
| 呼吸器官 | 支气管平滑肌舒张 | 支气管平滑肌收缩,促进粘膜腺分泌 |
| 消化器官 | 分泌粘稠唾液,抑制胃肠运动,促进括约肌收缩,抑制胆囊活动 | 分泌稀薄唾液,促进胃液、胰液分泌,促进胃肠运动和使括约肌舒张,促进胆囊收缩 |
| 泌尿生殖器官 | 促进肾小管的重吸收,使逼尿肌舒张和括约肌收缩,使有孕子宫收缩,无孕子宫舒张 | 使逼尿肌收缩和括约肌舒张 |
| 眼 | 使虹膜辐射肌收缩,瞳孔扩大使睫状体辐射状肌收缩,睫状体增大 使上眼睑平滑肌收缩 |
使虹膜环形肌收缩,瞳孔缩小 使眼下状体环形肌收缩,睫状体环缩小 促进泪腺分泌 |
| 皮肤 | 竖毛肌收缩,汗腺分泌 | |
| 代谢 | 促进糖原分解,促进肾上腺髓质分泌 | 促进胰岛素分泌 |
自主神经对效应器的支配,一般具有持久的紧张性作用,例如,切断支配心脏的迷走神经,则心率增加,说明心迷走神经本来有紧张性冲动传出,对心脏具有持久的抑制作用;切断心交感神经,则心率关慢,说明心交感神经也有紧张性冲动传出。又如,切断支配虹膜的副交感神经,则瞳孔散大;切断其交感神经,则瞳孔缩小,也说明自主神经的活动具有紧张性。自主神经中枢具有紧张性冲动传出的原因是多方面的,其中有反射性和体液性原因。例如来自主动脉弓和颈动脉窦区域的压力和化学感受器的传入冲动,对维持自主神经的紧张性活动有重要作用;而中枢神经组织内CO2浓度,对维持交感缩血管中枢的紧张性活动与有重要作用。
自主神经的外周性作用与效应器本身的功能状态有关。例如,刺激交感神经可引致动物无孕子宫的运动受到抑制,而对有孕子宫却可加强其运动(因为无孕与有孕子宫的受体不一样);又如,胃幽门如果原来处于收缩状态,则刺激迷走神经使之舒张,如原来处于舒张状态,则刺激迷走神经使之收缩。
交感神经系统的活动一般比较广泛,常以整个系统参与反应。例如,当交感神经系统发生反射性兴奋时,除心血管功能亢进外,还伴有瞳孔散大、支气管扩张、胃肠活动抑制等反应。有人认为,交感神经系统作为一个完整的系统进行活动时,其主要作用在于促使运动机体能适应环境的急聚变化。在剧烈肌肉运动、窒息、失血或冷冻等情况下,机体出现心率加速、皮肤与腹腔内脏血管收缩、血液贮存库排出血液以增加循环轿量、红细胞计数增加、支气管扩张、胆糖原分解加速以及血糖浓度上升、肾上腺素分泌增加等现象,这些现象大多是由于交感神经系统活动亢进所造成的。所以,交感神经系统在环境争聚变化的条件下,可以动员机体许多器官的潜在力量,以适应环境的急变。交感神经系统活动具有广泛性,但并不意味着毫无选择性,实际上交感神经系统发生反射反应时,各部位的交感神经活动还是有判别的。例如,失血后的开始10分钟内,交感神经传出的活动增加,主要表现为心脏活动增强与腹腔内脏血管的收缩,而其他反应就明显;又如,加温刺激下丘脑引致体温调节反应时,皮肤血管的交感神经活动减弱而使皮肤血流增加,但内脏血管的交感神经活动却增强。这些都说明,交感神经系统的反应还是具有相对选择性的。
副交感神经系统的活动,不如交感神经系统的活动那样广泛,而是比较局限的。其整个系统的活动主要在于保护机体、休整恢复、促进消化、积蓄能量以及加强排泄和生殖功能等方面。例如,心脏活动的抑制,瞳孔缩小避免强光的进入,消化道功能增强以促进营养物质吸收和能量补给等,这些都是副交感神经积蓄能量和保护机体的例子。
前文已述及自主神经末梢是通过释放递质而发挥作用的,副交感神经节后纤维和支配汗腺的交感神经节后纤维的递质均为乙酰胆碱,且均属于M样作用;当有机磷中毒时,由于胆碱酯酶失去活性,乙酰胆碱不能被水解而失活,这时就会出现广泛的副交感神经系统兴奋的症状(支气管痉挛、瞳孔缩小、流涎、大小便失禁等),同时大汗淋漓。这些症状均可被大剂量M受体阻断剂(阿托品)所解除,起到抢救的作用。但是,阿托品并不能恢复胆碱酯酶的活性,也不能解除乙酰胆碱N样作用的症状(如骨骼肌颤动),因此抢救时要和胆碱酯酶复活剂(解磷定、氯磷定)联合使用,才能收到更好效果。
二、脊髓对内脏活动的调节
交感神经和部分副交感神经发源于脊髓的外侧柱及相当于外侧柱的部位,因此脊髓可以成为内脏反射活动的初级中枢。在脊髓颈第五节段以上离断的动物,脊休克过去以后,血压可以上升恢复到一定水平,说明脊髓中枢可以完成基本的血管张力反射,以维持血管的紧张性,保持一定的外周阻力;同时还可具有反射性排尿和排粪的能力,说明基本的排尿反射与排便反射可以脊髓中枢内完成。在脊髓高位离断的病人,脊休克过去以后,也可见到血管张力发射、发汗反射、排快活反射、勃起反射的恢复。但是,这种反射调节功能是初级的,不能很好适应生理功能的需要。例如,当由平卧位转成站立时,病人就感到头晕;因为这时体位性血压反射的调节能力很差,脊髓以上的心血管中枢活动已不能控制脊髓的初级中枢,血管的外周阻已不能及时发生改变。又如,损害性刺激虽可以引致发汗反射,但由下丘控制的体温调节性发汗功能去消失。再如,基本的排尿反射可以进行,但排尿不能受意识控制,而且排尿也不完全;但在脊髓离断的病人,搔爬骶部皮肤也可以反射性地引致膀胱收缩而排尿,这使病人能够在一定程度上自己掌握排尿反射。
三、低位脑干对内脏活动的调节
由延髓发出的自主神经传出纤维支配头部的所有腺体、心、支气管、喉头、食管、胃、胰腺、肝和小肠等;同时,脑干网状结构中存在许多与内脏活动功能有关的神经元,其下行纤维支配脊髓,调节着脊髓的自主神经功能。因此,许多基本生命现象(如循环、呼吸等)的反射调节在延髓水平已能初步完成。临床观察和动物实验观察证明,延髓由于受压、穿现等原因而受损时,可迅速造成死亡,以致有人称延髓为基本生命中枢,延髓中的心血管功能、呼吸功能、消化功能等反射调节中枢,已分别在有关章节叙述,不再重复。
此外,中脑是瞳孔对光反射的中枢部位。中脑和下丘脑、边缘前脑对自主神经功能的调节是不可分割的。
四、下丘脑
下丘脑大致可分为四区,即前区、内侧区、外侧区与后区(图10-41)。前区的最前端为视前核,严格说来它属于前脑的范畴,稍后为视上核、视交叉上核、室旁核,再后是下丘脑前核,内侧区又称结节区,紧靠着下丘脑前核,其中有腹内侧核、背内侧核、结节核与灰白结节,还有弓状核与结节乳头核。外侧区有分解的下丘脑外侧核,其间穿插有内侧前脑束。后区主要是下丘脑后核与乳头体核。
图10-41 下丘脑神经核群示意图
A:前连合 1:外侧视前核 2:内侧视前核 3:室旁核4:下丘脑前核
5:视交叉上核 6:视上核 7:下丘脑背侧核8:下丘脑腹侧内侧核
9:下丘脑后侧核 10,11,12,13:乳头体核群14:中脑之脚间核
15:下丘脑外侧核 16:缰纹 17:穹窿 18:后屈束横贯下丘脑的纤维为内侧前脑束
下丘脑与边缘前脑及脑干网状结构紧密的形态和功能方面的联系,共同调节着内脏的省城。进入下丘脑的传入冲动可来逢边缘前脑、丘脑、脑干网状结构;其传出冲动也可抵达这些部位,还可通过垂体门脉系统和下丘脑-垂体束调节垂体前叶和后叶的活动。垂体门脉是正中隆起(灰白结节的内侧前部)与腺垂体之间的门脉系统;许多含有分泌颗粒的神经末梢终止于正中隆起,其分泌物可通过这一门脉系统到达腺垂体,调节腺垂体的活动,下丘脑-垂体束是由视上核、室旁核和结节发出的神经纤维束,它经垂体柄到达神经垂体,与神经垂体的活动密切相关。
在实验中,曾经观察到电刺激下丘脑的后区可获得血压升高、心率加速、瞳孔散大等交感神经性反应;因此有人认为下丘脑的后部是交感神经中枢,而前部是副交感神经不枢。但这个概念没有得到足够实验事实的支持,已不被公认。现在知道,下丘脑不是单纯的交感和副交感神经中枢,而是较高级的调节内脏活动和其它生理活动联系起来,调节着体温、营养摄取、水平衡、内分泌、情绪反应、生物节律等重要生理过程。
(一)体温调节
哺乳类运动在下丘脑以下部位横切脑干后,即不能保持体温的相对稳定;而在间脑以上切除大脑皮层的动物,体温仍能基本保持相对稳定。可见在间脑水平存在着体温调节中枢。现已肯定,调节体温的中枢在下丘脑。有人认为,体温调节中枢内有些部位能感知温度当血温超过或低于一定水平(这水平称为调定点,正常时约为36.8℃)时,即可通过调节产热和散热活动使体温保持相对稳定。体温调节中枢内的另一些部位对温度变化不敏感,但在温度敏感区的作用下,发出传出冲动以改变与产热和散絷有关器官的活动,从而保持体温的相对稳定。因此下丘脑的体温调节中枢,包括温度感受部分和控制产热和散热功能的整合作用部分。
(二)摄食行为调节
用埋藏电极刺激动物下丘脑外侧区,引致动物多食,而破坏此核后,则动物食欲增大而逐渐肥胖。由此认为,下丘脑外侧区存在摄食中枢(feeding center)。而腹内侧核存在饱中枢(satiety center),后者可以抑制前者活动。用微电极分别记录下丘脑外侧区和腹内侧核的神经元放电,观察到动物在饥锇时,前者放电频率较高而后者放电频率较低;静脉注入葡萄糖后,则前者放频率减少而后者放电频率增多。说明摄食中枢与饱中枢的神经元活动具有相互制约的关系,而且这些神经元对血糖敏感,血糖水平的高低可能调节着摄食中枢和饱中枢的活动。用电渗法(electroosmosis)法将葡萄糖注到腹内侧核神经元旁,也能使神经元放电频率增加,进一步说明饱中枢神经元对葡萄糖敏感。
(三)水平衡调节
水平衡包括水的摄入与排出两个方面,人体通过渴觉引起摄水,而排水则主要取决于肾的活动。损坏下丘脑可引致烦渴与多尿,说明下丘脑对水的摄入与排出调节均关系。
下丘脑控制摄入的区域与上述摄食中枢极为靠近。破坏下丘脑外侧区后,动物除拒食外,饮水也明显减少;刺激下丘脑外侧区某引起部位,则可引致动物饮水增多但是,控制摄水的中枢在确切部位还不清楚,不同动物的实验结果也不一致。
下丘脑控制排水的功能是通过改变抗利尿激素的分泌来完成的。抗利尿激素是由视上核和室旁核的神经元(神经分泌大细胞)合成的,神经分泌颗粒沿下丘脑-垂体束纤维向外周运输(轴浆运输)而贮存于神经垂体。以高渗盐水注入动物的颈内动脉,则能刺激抗利尿激素的分泌;如注入低渗盐水则抑制抗利尿激素的分泌。因此认为,下丘脑内存在着渗透压感受器,它能按血液的渗透压变化来调节抗利尿激素的分泌,此种感受器可能就在视上核和室旁核内。电生理研究观察到,当颈动脉内注入高渗盐水时,视上核内某些神经元放电增多,这一事实支持渗透压感受器就在视上核内的推测。一般认为,下丘脑控制摄水的区域与控制抗利尿激素分泌的核团在功能上是有联系的,两者协同调节着水平衡。
(四)对腺垂体激素分泌的调节
下丘脑内有些神经元(神经分泌小细胞)能正确性调节腺垂体激素分泌的肽类物质,这些物质是:促甲状腺素释放激素、促性腺素释放激素、生长至少释放抑制激素、生长素释放激素、促肾上腺皮质激素释放激素、促黑素细胞激素释放因子、促黑素细胞激素释放抑制因子、催乳素释放因子、催乳素释放抑制因子等。这些肽类物质在全盛后即经轴突运输并分泌到正中隆起,由此经垂体门脉系统到达腺垂体,促进或抑制某种腺垂体激素的分泌。此处,下丘脑还有些神经元对血液中某些激素浓度的变化比较敏感,这咱神经元称为监察细胞;例如,前区的某些神经元对卵巢激素敏感,内侧区的某些神经元对肾上腺皮质激素敏感,另有一些区域的某些神经元对各种垂体促激素很敏感。这些监察细胞在感受血液中激素浓度变化的信息后,可以反馈调节上述肽类物质的分泌,从而更好地控制腺垂体的激素分泌活动。
(五)对情绪生理反应的影响
情绪是人类一种心理现象,但伴随着情绪活动也发生一系列生理变化。这些客观的生理变化,称为情绪生理反应。自主神经系统的情绪反应,可以表现为交感神经系统活动相对亢进的现象,例如猫对痛刺激产生情绪反应时,可以出现心率加速、血压上升、胃肠运动抑制、脚掌出汗、竖毛、瞳孔散大、脾收缩而血液中红细胞计数增加、血糖浓度上升,同时呼吸往往加深加快。人类在发怒情况下,也可见到类似的现象。自主神经系统的情绪反应,在某些情况下也可表现为副交感神经系统活动相对亢进的现象。例如,食物性嗅觉刺激可引致消化液分泌增加和胃肠运动加强,动物发生性兴奋时则生殖器官血管舒张;人类焦急不安可引致排尿排便次数加频,忧虑可引致消化液分泌加多,悲伤则流泪,某些人受惊吓会引致心率减慢。因此,情绪生理反应主要是交感和副交感神经系统活动两者对立统一的改变。持久的情绪活动会造成自主神经系统功能的紊乱。
在间脑水平以上切除大脑的猫,常出现一系列交感神经系统兴奋亢进的现象,并且张牙舞爪,好似正常猫在搏斗时一样,故称之为假怒(sham rage)。平时下丘脑的这种活动受到大脑的抑制而不易表现。切除大脑后则抑制解除,下丘脑的防御反应功能被释放出来,在微弱的刺激下就能激发强烈假怒反应。研究指出,下丘脑内存防御反应区(defense zone),它主要位于下丘脑近中线两旁的腹内侧区。在动物麻醉条件下,电刺激该区可获得骨骼肌的舒血管效应(通过胆碱能交感舒血管纤维),同时伴有血压上升、皮肤及小肠血管收缩、心率加速和其他交感神经性反应。在动物清醒条件下,电刺激该区还可出现防御性行为。此外,电刺激下丘脑外侧区或引致动物出现攻击撕杀行为,电刺激下丘脑背侧区则出现逃避性行为。可见,下丘脑与情绪生理反应的关系很密切。人类下丘脑的疾病也往往伴随着不正常的情绪生理反应。
(六)对生物节律的控制
机体内的各种活动常按一定的时间顺序发生变化,这种变化的节律称为生物节律(biorhygym)。人体许多生理功能都有日周期节律,例如血细胞数、体温、促肾上腺皮质激素分泌等一瑚一个波动周期。身体内各种不同细胞都具有各自的日周期节律,但在自然环境中生活的人体器官组织却表现统一的日周期节律,这说明体内有一个总的控制生物节律的中心,它能使各种位相不同的生物节律统一起来,趋于同步化。研究指出,下丘脑的视交叉上核可能是生物节律的控制中心,在小鼠中观察到视交叉上核神经元的代谢强度和放电活动都表现明确的日周期节律。在胚胎期,当视交叉上核与周围组织还未建立联系时,其代谢和放电活动的日周期节律就已存在。破坏小鼠的视交叉上核,可使原有的日周期节律性活动(如饮水、排尿)的日周期丧失。视交叉上核可通过视网膜-视交叉上核束与视觉感受装置发生联系,因此外环境的昼夜光照变化可影响视交叉上核的活动,从而使体内日周期节律与外环境的昼夜节律同步起来。
五、神经、内分泌和免疫功能的关系
神经与内分泌功能间有密切的关系,近来年的研究发现,神经、内分泌和免疫功能间也有密切的关系,并认为三者共同构成一个完整的调节网络。
(一)神经对免疫功能的作用
神经可以通过两条途径来影响免疫功能,一条是通过神经释放递质来发挥作用,另一条是通过改变内分泌的活动转而影响免疫功能。骨髓、胸腺、淋巴结等免疫器官均有自主神经进入,虽然神经纤维主要是支配血管的,但末梢释放的递质(去甲肾上腺素、乙酰胆碱、肽类)可以通过弥散而作用于免疫细胞。
去甲肾上腺素能抑制免疫反应,免疫细胞上有相应有肾上腺素能受体。乙酰胆碱能增强免疫反应,免疫细胞上的胆碱能受体主要为M型。脑啡肽能增强免疫反应,而β-内啡肽的作用比较多样,有时能促进免疫反应,有时则抑制免疫反应。
神经细胞在特定的条件下也可产生免疫因子,例如在内毒素处理后可产生白细胞介素-1(白介素-1)等。
(二)免疫系统对神经活动的影响
在大鼠实验中观察到,用注入羊红细胞的方法来诱导免疫反应,当抗体生成增多达顶峰时,下丘脑某些神经元的电活动增加1倍以上,提示免疫反应可以改变神经活动。在裸鼠中注入白介素-1,可以使下丘脑有关神经元释放更多的促肾上腺皮持激素释放激素,导致血中促肾上腺皮质激素和糖皮质激素升高几倍,说明白介素-1可以作用于下丘脑神经元。
(三)内分泌系统对免疫功能的影响
促肾上腺皮质激素释放激素能直接促使人外周白细胞(经内毒素预处理后)产生促肾上腺皮质激素和内啡肽。促肾上腺皮质激素具有抑制免疫反应的作用,糖皮质激素一般也具有抑制免疫反应的作用。雌激素、孕激素和雄激素均有抑制免疫功能的作用。促甲状腺素释放激素、促甲状腺素、甲状腺激素均有增强免疫功能的作用。生长激素也有增强免疫功能的作用。
(四)免疫系统对内分泌功能的影响
前文已述及白介素-1能作用于下丘脑而增加促肾上腺皮质激素和糖皮质激素的血中含量。在大鼠中观察到,注入羊红细胞诱导免疫反应达到高峰期间,血中糖皮质激素含量上升而甲状腺激素含量下降,这一机制可能是一种负反馈调节,使免疫反应受到压抑而不致过分。此外,较低浓度的白介素-1能使胰岛B细胞的胰岛素分泌增加。
免疫细胞具有内分泌细胞样功能。免疫细胞分泌折各种免疫因子均为多肽或蛋白质,可以认为免疫因子是免疫细胞产生的内分泌样物质。此外,免疫细胞还可产生一般的激素。例如,巨噬细胞经内毒素外理后能分泌促肾上腺皮质激素、β-内啡肽和脑啡肽,外周淋巴细胞在葡萄糖菌毒素A的刺激下可产生促甲状腺素。
六、大脑皮层对内脏活动的调节
(一)新皮层
在动物实验中电刺激新皮层,除了能引致躯体运动等反应以外,也可引致内脏活动的变化。刺激皮层内侧面4区一定部位,会产生直肠与膀胱运动的变化;刺激皮层外侧面一定部位,会产生呼吸、血管运动的变化;刺激4区底部,会产生消化道运动及唾液分泌的变化。刺激6区一定部位可引致竖毛与出汗,也会引致下肢血管反应的区域也与下肢躯体运动代表区相对应。此外,刺激8区和19区等,除了可引致眼外肌运动外,也可引致瞳也的反应。所有这些结果,说明新皮层与内脏活动有关,而且区域分布和躯体运动代表区的分布有一定的地方。电刺激人类大脑皮层也以觅到类似的结果。
(二)边缘叶
大脑半球内侧面皮层与脑干连接部和胼胝体旁的环周结构,曾被称为边缘叶(图10-42)。这部分结构从进化上是比较古旧的;其最内侧的一环状结构(包括海马、穹窿等)称为古皮层,其较外圈的一环状结构(包括扣带回、海马回等)称为旧皮层。这部分结构曾被认为只与嗅觉联系,而称为嗅脑;但现已明确,其功能远不止这些,而是调节内脏活动的重要中枢。由于边缘叶在结构和功能上和大脑皮层的岛叶、颞极、眶回等,以及皮层下的杏仁核、隔区、下丘脑前核等,是密切相关的,于是有人把边缘叶连同这些结构统称为边缘系统。更有人见到中脑的中央灰质、被盖等也与上述结构有密切的上下行纤维双向联系,于是把中脑的部分结构也包括在边缘系统和概念之中。由此出现了边缘前脑(limbicforebrain)与边缘中脑(limbic midbrain)的概念,前者包括海马、穹窿、海马回、扣带回、杏仁核、隔区、梨状区、岛叶、颞极、眶回等结构,后者指中脑的中央灰质、被盖的中央部分及外侧部、脚间核等。边缘前脑的功能比较复杂,现仅述及其几个方面。
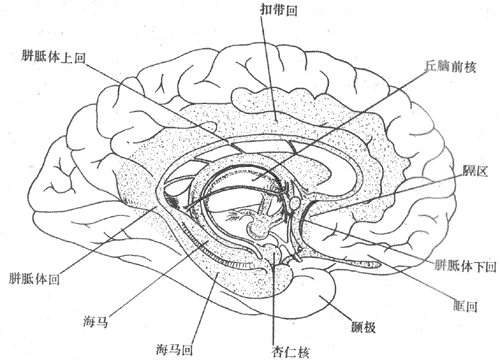
图10-42 大脑内侧面示边缘系统各部位
1.对情绪反应的影响 已知,杏仁核的活动与情绪反应有较密切的关系。杏仁核是由几个核群组成的复合体,从进化来看其中皮层内侧核群比较古老,基底外侧核群进化上较新。皮层内侧核群经终纹与下丘脑腹内侧核有联系,而基底外侧核群经腹侧杏仁传出系统也腹内侧核也有联系;前者有抑制腹内侧核神经元活动的作用,执行者有促进作用,。下丘脑与腹内侧区是防御反应区,因此前者有抑制防御反应作用,后者有易化防御反应的作用。
2.对摄食行为的影响下丘脑腹内侧核区即是防御反应区,也是饱中枢所在的部位。因此杏仁核队了能影响防御反应外,也能影响摄食行为。实验观察到,破坏杏仁核的猫,由于摄食过多而肥胖;用埋藏电极刺激杏仁核的基底外侧核群,可抑制摄食活动;同时记录杏佳核基底外侧核群和下丘脑外侧区(摄食中枢)的神经元放电,可见到两者的自发放电呈相互制约的关系,即当一个放电增我则另一个放电就减少。由此看来,杏仁核的基底外侧核群能易化饱中枢并抑制摄食中枢的活动。此外,刺激隔区,也可观察到相似的结果,即易化饱中枢并抑制摄食中枢的活动。
3.与记忆功能的关系海马与记忆功能有关。由于治疗的需要而手术切除双侧颞中叶的病人,如扣伤了海马及有关结构,则引致近期记忆功能的丧失;手术后对日常遇到的事件丧失记忆力,丧失的程度常决定于损伤部位的大小。临床上还观察到,由于手术切除第三脑室囊肿而损伤了穹窿,也能使患者丧失近期记忆能力;而且还观察到乳头体或乳头体丘脑束的疾患也会导致近期记忆能力的丧失。把这些事实联系起来,可以认为与近期记忆功能有关的神经结构是一个环路结构,即海马→穹窿→下丘脑乳头体→丘脑前核→扣带回→海马,称为海马环路。
4.对其他内脏活动反应的影响刺激边缘前脑不同部位所引起的内脏活动反应是很复杂的,血压可以表现升高或降低,呼吸可以加快或抑制,胃肠运动可以加强或减弱,瞳孔可以扩大或缩小等。例如,刺激扣带回前部可出现患得患失抑制(刺激过强则呼吸加强)、血压下降或上升、心率变慢、胃运动抑制、瞳孔扩大或缩小;刺激杏仁核出现咀嚼、唾液分泌增加、胃酸分泌增加、胃蠕动增加、排便、心率减慢、瞳孔扩大;刺激隔区出现阴茎勃起、血压下降或上升、呼吸暂停或加强。这些运动实验结果,说明边缘前脑的功能和初级中枢的功能不一样;刺激初级中枢扳应比较肯定而一致,而刺激边缘前脑的结果变化较大。可以设想,初级中枢的功能比较局限,活动反应比较单纯;而边缘前脑是许多初级中枢活动的调节者,它能通过促进或抑制各初级中枢的活动,调节更为复杂的生理功能活动,因此活动反应也就复杂而多变。
第六节 脑的高级功能和脑电图
一、学习和记忆
学习和记忆是两个相联系的神经过程。学习指人和运动依赖于经验来改变自身行为以适应环境的神经活动过程。记忆则是学习到的信息贮存和“读出”的神经活动过程。
(一)学习的形式
1.简单学习简单学习不老大哥在刺激和反应之间形成某种明确的联系,又称为非联合型学习(nonassociative learning)。习惯化和敏感化属于这种类型的学习。习惯化是指当一个不产生伤害性效应的刺激重复作用时,机体对该刺激的反射反应逐渐减弱的过程,例如人们对有规律而重要出现的强噪音逐渐不再对它产生反应。敏感化是指反射反应加强的过程,例如一个弱伤害性刺激本仅引起弱的反应,但在强伤害性刺激作用后弱刺激的反应就明显加强。在这里,强刺激与弱刺激之间并不需要建立什么联系。
2.联合型学习经典条件反射和操作式条件反射均属于联合型学习。
(1)经典条件反射:在动物实验中,给狗吃食物会引起唾液分泌,这是非条件反射。给狗以铃声则不会引起唾液分泌,因为铃声与食物无关,这种情况下铃声称为无关刺激。但是,如果每次给狗吃食物以前先出现一次铃声,然后再给以食物,这样多次结合以后,当铃声一出现,动物就会出现唾液分泌。铃声本来是无关刺激,现在由于多次与食物结合应用,铃声具有了引起唾液分泌的作用,即铃声已成为进食(非条件刺激)的信号。所以这时就把铃声称为信号刺激或条件刺激,这样的反射就称为条件反射。可见,条件反射是在后天生活中形成的。形成条件反射的基本条件就是无关刺激与非条件刺激在时间上的结合,这个过程称为强化。任何无关刺激与非条件刺激结合应用,都可以形成条件反射。
(2)操作式条件反射:操作式条件反射比较复杂,它要求动物完成一定的操作。例如,将大鼠放入实验箱内,当它在走动中偶然踩在杠杆上时,即喂食以强化这一操作;如此重复多次,大鼠即学会了自动踩杠杆而得食。然后,在此基础上进一步训练动物只有当再现某一特定的信号(如灯光)后踩杠杆,才能得到食物的强化。在训练完成后,动物见到特定的信号,就去踩杠杆而得食。这类条件反射的特点是,动物必须通过自己完成某种运动或操作后才能得到强化,所以称为操作式条件反射。
(二)条件反射活动活动的基本规律
1.经典条件反射建立需要的基本条件条件反射的建立要求在时间上把某一无关刺激与非条件刺激结合多次,一般条件刺激要先于非条件刺激而出现。条件反射的建立与动物机体的状态有很密切的关系,例如处于饱食状态的运动则很难建立食物性条件反射,动物处于困倦状态也很难建立条件反射。一般来说,任何一个能为机体所感觉的动因均可作为条件刺激,而且在所有的非条件刺激的基础上都可建立条件反射,例如食物性条件反射、防御性条件反射等。
2.经典条件反射的消退条件反射建立之后,如果反复应用条件刺激而不给予非条件刺激强化,条件反射就会逐渐减弱,最后完全不出现。这称为条件反射的消退。例如,铃声与食物多次结合应用,使狗建立了条件反射;然后,反复单独应用铃声而不给予食物(不强化)则铃声引起的唾液分泌量会逐渐减少,最后完全不能引起分泌。巴甫洛夫认为,条件反射的消退是由于在不强化的条件下,原来引起唾液分泌的条件刺激,转化成为引起中枢发生抑制的刺激。从这一观点出发,条件反射的消退并不是条件反射的丧失,而是人原先引起兴奋(有唾液分泌)的条件反射转化为引起抑制(无唾液分泌)的条件反射;前者称为阳性条件反射,后者称为阴性条件反射。
3.人类的条件反射研究动物条件反射的方法,原则上也可用于研究人的条件反射活动;例如,将无关刺激与食物性唾液分泌非条件反射相结合(可用于儿童),或将无关刺激与防御性运动非条件反射相结合等等。此外,人类还可应用词语强化的运动条件反射研究法;例如,当红光在受度儿童面前出现时,实验者说:“按”,受试儿音即用手按压橡皮球。在这一实验中,红光是条件刺激,“按”是词语强化。用词语强化与红光结合2-3次后,如果受试者见到红光信号出现后,立即自动按球,这就形成了对红光的条件反射。
4.两种信号系统学说在人同样可以用光、声、嗅、味、触等感觉刺激作为信号来形成条件反射;这种信号直接作用于眼、耳、鼻、舌、身等感受装置,都是现实具体的信号。此外,抽象的语词也可以代替具体的信号而引起条件反射反应。例如,受试者对每分钟摆动120次的快节拍器声音形成了用温热刺激强化的手臂血管舒张反射,而对每分钟摆动60次的慢节拍器声音形成了用冷刺激强化的血管收缩反射;当这些条件反射被巩固后,实验者对受试者说“快节拍器音”或“慢节拍器音”,这些语词也分别能引起相应有血管舒张或血管收缩反应。如果说具体的信号是第一信号,则相应的语词是第一信号的信号,即第二信号。因此,在人类有两种性质完全不同的信号,第一信号是具体的信号,第二信号(语词)是抽象的信号。巴甫洛夫提出人脑有两个信号系统。第一信号系统是对第一信号发生反应的大脑皮层功能系统,第二信号系统是对第二信号发生反应的大脑皮层功能系统。动物只有一个信号系统,相当于人的第一信号系统;而人类才具有两个信号系统,这是人类区别于动物的主要特征。第二信号系统的发生与发展是人类社会的产物,人类由于社会性劳动与交往产生了语言,语词是现实有概括和抽象化;人类借助于语词来表达其思维,并进行抽象的思维。
(三)记忆的过程
外界通过感觉器官进入大脑的信息量是很大的,但估计仅有1%的信息能被较长期地贮存记忆,而大部分却被遗忘。能被长期贮存的信息都是对个体具有重要意义的,而且是反复作用的信息。因此,在信息贮存过程中必然包含着对信息的选择和遗忘两个因素。信息的贮存要经过多个步骤,但简略地可把记忆划分为两个阶段,即短时性记忆和长时性记忆。在短时性记忆中,信息的贮存是不牢固的,例如,对于一个电话号码,当人们刚刚看过但没有通过反复运用而转入长时性记忆的话,很快便会遗忘。但如果通过较长时间的反复运动,则所形成的痕迹将随每一次的使用而加强起来;最后可形成一种非常牢固的记忆,这种记忆不易受干扰而发生障碍。
人类的记忆过程可以细分成四个阶段(图10-43),即感觉性记忆、第一级记忆、第二级记忆和第三级记忆;前二个阶段相当于上述的短时性记忆,后二个阶段相当于长时性记忆。感觉性记忆是指通过感觉系统获得信息后,首先在脑的感觉区内贮存的阶段;这阶段贮存的时间很短,一般不超过1分钟,如果没有经过注意和处理就会很快消失。如果住处在这阶段经过加工处理,把那引起不持续的、先后进来的信息整合成新的连续的印象,就可以从短暂的感觉性记忆转入第一级记忆。这种转移一般可通过两种途径来实现,一种是通过把感觉性高蛋白的资料变成口头表达性的符号(如语言符号)而转移到第一级记忆,这是最常见的;另一种非口头表达性的途径,这在目前还了解得不多,但它必然是幼儿学习所必须采取的途径。但是,信息在第一级记忆中停留的的时间仍然很短暂,平均约几秒钟;通过反复运用学习,信息便在第一级记忆中循环,从布延长了信息在第一级记忆中停留的时间,这样就使信息容易转入第二级记忆之中。第二级记忆是一个大而持久的贮存系统。发生在第二级记忆内的遗忘,似乎是由于先前的或后来的信息的干扰所造成的;这种干扰分别称为前活动干扰和后活动性干扰。有些记忆的痕迹,如自己的名字和每天都在进行操作的手艺等,通过长年累月的运动,是不易遗忘的,这一类记忆是贮存在第三级记忆中的。
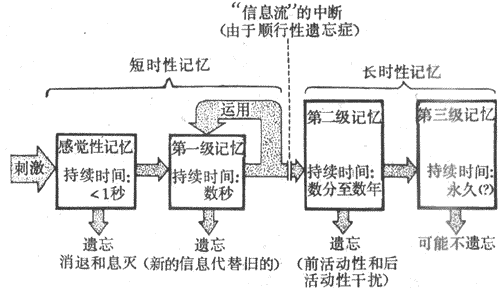
图10-43从感觉性记忆至第三级记忆的信息流图解
(四)记忆障碍
临床上把记忆障碍分为两类,即顺行性遗忘症(anterogradeamnesia)和逆行性遗忘症(retrograde amnesia)。凡不能保留新近获得的信息的称为顺行性遗忘症。患者对于一个新的感觉性信息虽能作出合适的反应,但只限于该刺激出现时,一旦该刺激物消失,患者在数秒钟就失去作出正确反应的能力。所以患者易忘近事,而远的记忆仍存在。本症多见于慢性酒精中毒者。发生本症的机制,可能是由于信息不能从第一级记忆转入第二级记忆;一般认为,这种障碍与海马的功能损坏有关。前文已述及,海马及其环路的功能遭受破坏,会发生近期记忆障碍。凡正常脑功能发生障碍之前的一段时间内的记忆均已丧失的,称为逆行性遗忘症;患者不能回忆起紧接着本症发生前一段时间的经历。一些非特异性脑疾患(脑震荡、电击等)和麻醉均可引起本症。例如,车祸造成脑震荡的患者在恢复后,不能记起发生车祸前一段时期内的事情,但自己的名字等仍能记得。所以,发生本症的机制可能是第二级记忆发生了紊乱,而第三级记忆却不受影响。
(五)学习和记忆的机制
1.从神经生理角度看学习和记忆的机制从神经生理的角度来看,感觉性记忆和第一级记忆主要是神经元生理活动的菜单现。神经元活动具有一定的后作用,在刺激作用过去以后,活动仍存留一定时间,这是记忆的最简单的形式,感觉性记忆的机制可能属于这一类,在神经系统中,神经元之间形成许多环路联系,环路的连续活动也是记忆的一种形式,第一级记忆的机制可能属于这一类。例如,海马环路的活动就与第一级记忆的保持以及第一级记忆转入第二级记忆有关。
近年来对突触传递过程的变化与学习记忆的关系进行了许多研究。在海兔(一种海洋软体动物)的缩鳃反射的研究中观察到,习惯化的发生是由于突触传递出现了改变,突触前末梢的递质释放量减少导致突触后电位减少,从而使反射反应逐渐减弱;敏感化的机制是突触传递效能的增强,突触前末梢的递质释放量增加。在高等动物中也观察到突触传递具有可塑性。有人在麻醉兔中,记录海马齿状回颗粒细胞的电活动观察到,如先以一串电脉冲刺激海马的传入纤维(前穿质纤维),再用单个电刺激来测试颗粒细胞电活动改变,则兴奋性突触后电位和锋电位波幅增大,锋电位的潜伏期缩短。这种易化现象持续时间可长达10小时以上,并被称为长时程增强(long-term potentiation)。不少人把长时程增强与学习记忆联系起来,认为它可能是学习记忆的神经基础。在训练大鼠进行旋转平台的空间分辨学习过程中,记忆能力强的大鼠海马长时程增强反应大,而记忆能力差的大鼠长时程增强反应小。
2.从神经生化角度看学习和记忆的机制从神经生化的角度来看,较长时性的记忆必然与脑内的物质代谢有关,尤其是与脑内蛋白质的合成有关。在金鱼建立条件反射的过程中,如用嘌呤霉素(puromycin)注入动物脑内以抑制脑内蛋白质的合成,则运动不能完成条件反射的建立,学习记忆能力发生明显障碍。人类的第二级记忆可能与这一类机制关系较大。在逆行性遗忘症中,可能就是由于脑内蛋白质合成代谢受到了破坏,以致使前一段时间的记忆丧失。
中枢递质与学习记忆活动也有关。运动学习训练后注射拟胆碱药毒扁豆碱可加强记忆活动,而注射抗胆碱药东莨菪硷可使学习记忆减退。用利血平使脑内儿茶酚胺耗竭,则破坏学习记忆过程。动物在训练后,在脑室内注入γ-氨基丁酸可加速学习。动物训练后将加压素注入海马齿状回可增强记忆,而注入催产素则使记忆减退。一定量的脑啡可使动物学习过程遭受破坏,而纳洛酮可增强记忆。临床研究发现,老年人血液中垂体后叶激素含量减少,用加压素喷鼻可使记忆效率提高;用加压素治疗遗忘症亦收到满意效果。
3.从神经解剖角度看学习和记忆的机制从神经解剖的角度来看,持久性记忆可能与新的突触联系的建立有关。动物实验中观察到,生活在复杂环境中的大鼠,其大脑皮层的厚度大,而生活在简单环境中的大鼠,其大脑皮层的厚度小;说明学习记忆活动多的大鼠,其大脑皮层发达,突触的联系多。人类的第三级记忆的机制可能属于这一类。
二、大脑皮层的语言中枢和一侧优势
(一)两侧大脑皮层功能的相关
两侧大脑皮层之间有许多连合纤维,在哺乳类动物中最大的连合纤维结构是胼胝体;进化愈高等则胼胝体愈发达,人类的胼胝体估计含有100万根纤维。有人观察到,当在犬的身体一侧皮肤上给予刺激,并与食物或酸防御性唾液分泌反射相结合形成条件反射后,则另一侧皮肤相应部位的机械刺激也自然具有阳性的条件反射效应。如果事先将该动物的胼胝体切断,则这种现象就不能出现。还有人事先切断猫视交叉的交叉纤维,使一侧眼睛的视网膜传入冲动仅向同侧皮层投射,然后将动物一眼蒙蔽,用另正好学习对图案的鉴别能力;待其学会后将该眼蒙蔽,测定先前被蒙蔽眼的图案鉴别能力,见到先前被蒙蔽的眼也能具有这种鉴别能力。如果事先切断这个动物的胼胝体,则这种现象就不能出现。由此看来,两侧大脑皮层的感觉分析功能是相关的,胼胝体连合纤维能将一侧皮层的活动向另一侧转送。电生理研究指出,刺激对侧皮层对应点可以加强这一侧皮层的感觉传入冲动的诱发电们,起着易化作用。这一易化作用是通过胼胝体连合纤维完成的,因为这类纤维主要联系两侧皮层相对应的部位。在人类,两侧大脑皮层的功能也是相关的,两关球之间的连合纤维对完成双侧的运动、一般感觉和视觉的功调功能有重要作用。右手学会了一种技巧运动,左手虽然没有经过训练,但在一定程度上也会完成这种技巧运动,说明一侧皮层的学习活动可以通过连合纤维向另一侧转送。
(二)大脑皮层的语言中枢
人类大脑皮层一定区域的损伤,可以引致特有的各种语言活动功能障碍(图10-44)。临床发现,损伤布洛卡(Broca)三角区(44区,在中央前回底部之前,图中S区),会引致运动失语症(motor aphasia)。病人可以看懂文字与听懂别人谈话,但自己却不会讲话,不能用语词来口头表达;然而,其与发音有关的肌肉并不麻痹,就是不能用“词”来表达自己的意思。损伤额中回后部接近中央前回手部代表区的部位(图中W区),则病人可以听懂别人的谈话,看懂文字,自己也会讲话,但不会书写;然而,其手部的其他运动并不受影响,这种情况称为失写症(agraphia)。颞上回后部(图中H区)的损伤,会引致感觉失语症(sensory aphasia),病人可以讲话及书写,也能看懂文字,但听不懂别人的谈话;事实上,病人能听到别人的发音,就是不懂其含义,但其视觉却是良好的,其他的语言活动功能仍健全,这种情况称为失读症(alexia),因此,语言活动的完整功能是与广大皮层区域的活动有关的,各区域的功能是密切相关的。严重的失语症可同时出现上述四种语言活动功能的障碍。
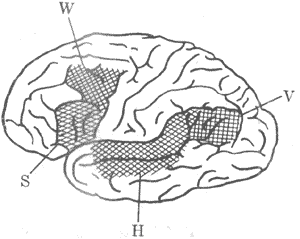
图10-44 人大脑皮层语言功能的区域
V区障碍不能认识词义 H区障碍不能听懂话 S区障碍不能讲话 W区障碍不能书写
(三)大脑皮层功能的一侧优势
产生上述各种语言活动功能障碍时,在一般运用右手劳动为主的成年人中,其大脑皮层损伤经常发生在左侧。因为绝大多数用右手劳动为主的成年人,右侧大脑皮层的44区的损伤并不发生明显的语言活动障碍;然而其左侧大脑皮层布洛卡的损伤,则可形成严重的运动失语症,这种左侧大脑皮层的语言活动功能上占优势的现象,反映了人类两侧大脑半球功能是不对等的,这种一侧优势的现象仅在人类中具有。
人类左侧大脑皮层在语言活动功能上占优势的现象,虽然与一定的遗传因素有关,但主要是在后天生活实践中逐步形成的,这与人类习惯运用右手进行劳动有密切的关系。小儿在2-3岁之前,如果发生左侧大脑半球损害时,其语言活动功能的紊乱和右侧大脑半球损害时的情况没有明显的差别;说明这时候尚未建立左侧优势,双侧大脑半班干部均与语言活动功能有关。10-12岁时,左侧优势逐步建立;但在左侧大脑半球损害后,尚有可能在右侧大脑半球损害就很难在右侧大脑皮层再建立起语言活动的中枢。在发育成年人后,左侧优势已经形成,如果发生左侧大脑半球损害就很难在右侧大脑皮层再建立起语言活动的中枢。在运用左手劳动为主的人中,则左右双侧的皮层有关区域都可能成为语言活动的中枢。有人进行过统计,在48例运用右手劳动的人中,语言中枢在左侧的为43例,在右侧的为25例,4例左右双侧均有关;在20例左右手混用的人中,语言中枢在左侧的12例,在右侧的为2例,6例左右双侧均有关。
由于左侧大脑半球在语言活动功能上占优势,因此一般称左侧半球为优势半球或主要半球,右侧半球为次要半球。但是研究指出,右侧半球也有其特殊的重要功能。目前知道,右侧大脑皮层在非语词性的认识功能上是占优势的,例如对于空间的辩认、深度知觉、触觉认识、音乐欣赏分辨等等。右侧大脑皮层顶叶损伤的病人,由于非语词性认识能力的障碍,常再现穿衣失用症(apraxia);患者虽然没有肌肉麻痹,但穿衣困难,他会将衬衣前后穿倒或只将一只胳膊伸入袖内。右侧大脑皮层顶叶、枕叶、颞叶结合处损伤的病人,常分不清左右侧,穿衣困难,不以维持绘制图表。右侧大脑半球后部的病变,常发生视觉认识障碍;患者不能辨认别人的面部,甚至不能认识镜子里自己的面部,而且还伴有对颜色、物体、地方的认识障碍。
上述两侧大脑半球对不同认识功能的优势现象,还可通过裂脑(splitbrain)实验研究加以证实。在患有顽固性癫痫发作的病人,为了控制癫痫在两半球之间传布发作,常将患者的连合纤维(胼胝体)切断;手术后患者对出现在左侧视野中的物体(视觉投射到右侧半球)不能用语词说出物体的名称,而对出现在右侧视野中的物体(视觉投射到左侧半球)就以臁出物体的名称,说明语言活动中枢在左侧半球。但是,患者右侧半球的视觉认识功能是良好的。譬如,先给患者的左侧视野看一支香烟,他不能用语词说出这一物体是“香烟”;但是患者认识到这一物体是香烟,因为他可以闭着眼睛藉助于触觉有矩手把许多香烟收集起来以表示他对这一物体的认识。在正常人,虽然语言活动中枢在左侧半球,但能对左侧视野中的物体说出其名称,这是连合纤维的功能,因为连合纤维使左右两侧半球的功能发生了联系。
一侧优势是指人类的脑的高级功能向一侧半球集中的现象;左侧半球在语词活动功能上占优势,右侧半球在非语词性认识功能止占优势。但是,这种优势敢是相对的,而不是绝对的;因为左而半球也有一定的非语词性认识功能,右侧半球也有一定的简单的语词活动功能。
三、脑电图和脑诱发电位
大脑皮层的神经元具有生物电活动,因此大脑皮层经常有持续的节律性电位改变,称为自发脑电活动。临床上在头皮用双极或单极记录法来观察皮层的电位变化,记录到的脑电波称为脑电图(图10-45)。在动物中将颅骨打开或以病人进行脑外科手术时,直接在皮层表面引导的电位变化,称为皮层电图。此外,在感觉传入冲动的激发下,脑的某一区域可以产较为局限的电位变化,称为脑诱发电位。
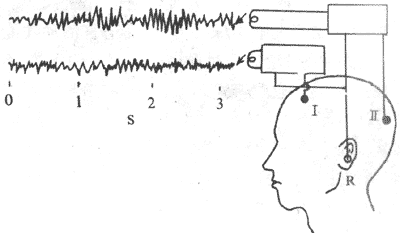
图10-45 脑电图记录示意图
无关电极放置在耳壳(R),由额叶(I)电极导出的脑电波振幅低,
由枕叶(Ⅱ)导出的脑电波振幅高频率较慢
(一)脑电图的波形
脑电图的波形分类,主要是依据其频率的不同来人工划分的。在不同条件下,波形频率的快慢可有显着的差别,每秒0.5-3次的波称为δ波,4-7次的波称为θ波,8-13次的波称为α波,14-30次的波称为β波。一般说来,频率慢的波其波幅常比较大,而频率快的波其波幅就比较小。例如,在成年人头上皮上引导时,δ波可有20-200μV,α波有20-100μV,而β波只有5-20μV(图10-46)。
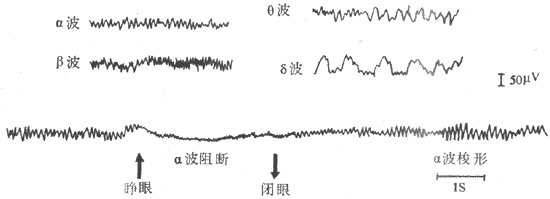
图10-46 正常脑电波各种波形
各种波都可在皮层的不同区域引得,但枕叶区域其α波活动比较显著,而β波在额叶与顶叶比较显著。有时,β波与α波同时在一个部位出现,而β波重合在α波的上面。人类α波在清醒、安静并闭眼时即出现。α波出现时,在枕叶部位最大,并可具有时大时小的变化;即波幅先由小逐渐变大,然后又由右面变小,接着又由小变大,如此反复,形成α波的梭形,每一梭形持续约1-2s。睁开眼睛或接受其他刺激时,α波立即消失而呈现快波,这一现象称为α波阻断,如果被试者又安静闭眼时,则α波又重现。在困倦时,一般可见θ波。成人清醒状态下,几乎是没有θ波的,但在睡眠期间皮层脑电图可出现δ波。如将睡者唤醒,δ波即转成快波。因此,一肌认为快波是新皮层处在紧张活动状态时的主要脑电活动表现,α波是皮层处在安静状态时的主要表现,慢波是睡眠状态下的主要表现。在幼儿时期,脑电波频率比成人慢,一般常见到θ波,到10岁后才出现明确的α波;在婴儿时期,脑电波频率更慢,常见到δ波。此外δ波在成年人极度疲劳时及麻醉状态下也可出现。
癫痫患者,脑电图可出现棘波、尖波、棘慢综合波等,棘波的时程在80ms以下,幅度为50-150μV。尖波的时程为80-200ms,幅度为100-200μV。棘慢综合波指的是棘波后跟随出现一个慢波,慢波时程达200-500ms;一般棘慢综合波出现时,多数为每秒3次左右(图10-47)。在皮层具有占位性病变(肿瘤等)的区域,即使病人外于清醒状态时,亦可引出θ波或δ波。因此,临床上可以借这些脑电波改变的特点,并结合临床资料,来诊断癫痫或探索肿瘤的所在部位。
图10-47脑电图正常波形与癫痫波形的对比
(二)脑电波形成的机制
脑电波以的波形是一种近似于正弦波的电位变化,而与神经干上见到的动作电位不一样。应用微电极记录皮层神经元细胞内电位变化,见到皮层表面出现类似α波节律的电位变化时,细胞内记录到的突触后电位变化也出现节律相一致的改变(图10-48)。由此认为,,此层表现的电位变化主要是由突触后电位变化形成的,也就是说由细胞体和树突的电位变化形成的。可以设想,单一神经元的突触后电位变化是不足以引起皮层表面的电位改变的;必须有大量的神经组织同时发生突触后电位变化,才能同步起来引起皮层表面再现电位改变。从皮层的神经元组成来看,锥体细胞的分布排列比较整齐,其顶树突互相平行并垂直于皮层表面,因此其电活动在同步时易于总和而形成强大的电场,从而改变皮层表面的电位。

图10-48脑电图与皮层神经元细胞内电位记录的关系
目前知道,大量皮层神经组织的放电活动同步总和必须依赖丘脑的功能。在动物实验中见到,当用中度麻醉时,即使没有其他感觉传入的刺激,皮层会出现每称8-12次的自发脑电活动。这种脑电活动的波幅亦时大时小,并可以皮层广泛的区域内引出,因此这咱脑电活动与人类脑电波中的α节律极相似。如果切断皮层与丘脑间的纤维联系,上述类似α波的电活动就大大减小。如用每秒8-12次节律性电刺激来刺激丘脑非特异投射系统的一此神经核(如髓板内核群),则皮层上会出现每称8-12次的节律性脑电变化。这种变化的波幅亦时大时小,同时在皮层的空间分布也是广泛的,因此,从频率、波幅形状以及空间分布上来看,刺激丘脑非特投射系统所获得的脑电变化,与上述类似α波的自发脑电活动相一致。由此认为,某引起自发脑电形成的同步机制,就是皮层与丘脑非特异投射系统之间的交互作用;一定的同步节律的丘脑非特特异投射系统的活动,促进了电层电活动的同步化。
如果用每秒60次的节律性电刺激来刺激丘脑非特异投射系统,则皮层上类似α波的自发脑电活动立即消失而转成快波。这可理解为高频刺激对同步化活动的扰乱,脑电出现了去同步化现象,快波的出现就是去同步化的结果。刺激脑干网状结构时引起的上行激动作用,一般也认为是其上行冲动扰乱了丘脑非特异投射系统与皮层之间同步化环节,脑电出现了激活状态,呈现了去同步化的快波。在人类脑电记录中所见到的α波阻断现象,事实上也是由同样机制引起的。
电生理研究观察到,当皮层癫痫病灶区出现棘波时,皮层内神经元出现爆发式短串冲动发放,频率可高达每秒200-900次;如将电极插入神经元细胞体内,则观察到当棘波出现时,细胞体出现大幅度去极化电位(可达30mV),去极化电位发展到一定程度后则爆发短串动作电位。由此认为,许多神经元同时出现大幅度的去极化电位,就使皮层表现出现电棘波;而神经元的爆发式短串冲动发放,也是由于大幅度去极化电位造成的,这种大幅度去极化电位,可能是大量同步的兴奋性突触后电位总和起来形成的,这是癫痫病例灶区神经元异常活动的表现。由于皮层肿瘤等占位性病变区本身不产生电活动变化,因此在该区域记录到的θ波或δ波,一般认为来源于其环周异常脑细胞的电活动。
(三)脑诱发电位
诱发电位是指感觉传入系统受刺激时,在中枢神经系统内引起的电位变化。受刺激的部位可以是感觉器官、感觉神经或感觉传导途径上的任何一点。但是广义地说,用其他刺激方法引起的中枢神经系统的电位变化,也可称为诱发电位。例如,直接刺激脊髓前根,冲动沿运动神经逆向传至脊髓前多角引起的电位变化,亦可称为诱发电位。
大脑皮层诱发电位一般是指感觉传入系统受刺激时,在皮层上某一局限区域引出的电位变化;由于皮层随时在活动着并产生自发脑电波,因此诱发电位时常出现在自发脑电波的背景之上。在动物皮层相应的感觉区表面引起的诱发电位可分为两部分,一为主反应,另一为后发放(图10-49)。主反应出现的潜伏期是稳定不变的,为先正后负的电位变化。后发放尾随主反应之后,为一系列正相的周期电位变化。皮层诱发电位是用以寻找感觉投射部位的重要方法,在研究皮层功能定位方面起着重要的作用。
图10-49家兔大脑皮层感觉运动区诱发电位
上线:诱发电位记录,向下为正,向上为负
下线:时间,50ms第一个向上小波为刺激桡浅神经记号,
间隔10ms后即出现先正后负的主反应,再
间隔100ms左右后,即相继出现正相波动的后发放
诱发电位也可在人体头颅外头皮上记录到。由于记录电极离中枢较远,颅骨的电阻很大,记录到的电位变化极微弱;而且诱发电位夹杂在自发脑电之间,电位很难分辨。运用电子计算机将电位变化叠加、平均起来,能够使诱发电位显示出来,这种方法记录到的电位称为平均诱发电位(averaged evoked potential)。平均诱发电位目前已成为研究人类的感觉功能、神经系统疾病、行为和心理活动的一种手段。临床常用的有体感诱发电位、听觉诱发电位和视觉诱发电位几种。现简述体感诱发电位的引导方法和波形;刺激电极安放在上肢正中神经经过的皮肤表面(也可放在下肢的某一部位),记录电极放在颅顶靠近中央后回的头皮表面,参考电极置于耳壳;记录到的标准波形如图10-50所示。图中的P9波起源于正中神经的第一级神经元;P11波可能起源于脑干或颈脊髓,因为丘脑以上中枢病变时,P11不受影响,而颈脊髓病变时P11消失;P13和P14波可能由脑干内侧丘系活动所产生;N20波是一个负波,一般认为它来源于丘脑向皮层的投射或皮层感觉区,因为在丘脑病变时可使N20波消失,而N20波以前的电波成分不受影响。因此,通过体感诱发电位的记录和分析,有助于对患者中枢损伤位置的诊断。
四、觉醒和睡眠
觉醒和睡眠都是生理活动所必要的过程,只有在觉醒状态下,人体才能进行劳动和其他活动;而通过睡眠,可以使人体的精力和体力得到恢复,于睡眠后保持良好的觉醒状态。成年人一般每天需要7-9小时,儿童需要睡眠的时间比成年人长,而老年需要睡眠的时间就比较短。与觉醒对比,睡眠时许多生理功能发生了变化,一般表现为:①嗅、视、听、触等感觉功能暂时减退;②骨骼肌反射运动和肌紧张减弱;③伴有一系列自主神经功能的改变。例如,血压下降、心率减慢、瞳孔缩小、尿量减少、体温下降、代谢率减低、呼吸变慢、胃液分泌可增多而唾液分泌减少、民汗功能增强等。
(一)觉醒状态的维持
动物实验中观察到,单纯在中脑肉状结构的头端加以破坏,而保留各种感觉上传的特异传导途径,动物即进入持久的昏睡状态;各种感觉刺激都不能唤醒动物,脑电波不能由同步化慢波转化成去同步化快波,虽然这时感觉传入冲动完全可以沿特异传导途径抵达大脑皮层。因此认为,觉醒状态的维持是脑干网状结构上行激动系统的作用。目前认为,脑干网状结构上行激动系统可能是乙酰胆碱递质系统,因此静脉注射阿托品能阻断脑干网状结构对脑电的唤醒作用。
进一步的研究观察到,动物在注入阿托品后,脑电呈现同步化慢波而不再出现快波,但动物在行为上并不表现睡眠。看来,觉醒状态的纤维比较复杂,脑电觉醒状态(呈现快波)与行为觉醒状态的维持有不同的机制。动物实验观察到,单纯破坏中脑黑质多巴胺递质系统后,则动物在行为上不能表现觉醒,对新异的刺激不能表现控究行为,但脑电仍可有快波出现。因此,行为觉醒的维持可能是黑质多巴胺递质系统的功能。动物实验还见到,破坏蓝斑上部(去甲肾上腺素递质系统)后,则动物电波快波明显减少;但如有感觉刺激传入冲动时,则动物仍能唤醒脑电呈现快波,不过和种唤醒作用很短暂,感觉传入刺激一停止,唤醒作用即终止。所以,蓝斑上部去甲肾上腺素递质系统与脑电觉醒的维持也有关系。其作用是持续的紧张性作用;而上行激动系统(乙酰胆碱递质系统)的作用是时相性作用,它调制去甲肾上腺素递质系统的脑电觉醒作用。
(二)睡眠的时相
通过对整个睡眠过程的仔细观察,发现睡眠具有两种不同的时相状态。其一是脑电波呈现同步化慢波的时相,其二是脑电波呈现去同步化的时相。前者是一般熟知的状态,其表现已在前文述及,常称为慢波睡眠(slow wave sleep,SWW)。后者的表现与慢波睡眠不同,称为异相睡眠(paradoxicalsleep,PS)或快波睡眠、快速眼球运动 (rapid eyemovements,REM)睡眠。异相睡眠期间,各种感觉功能进一步减退,以致锅醒阈提高;骨骼肌反射运动和肌紧张进一步减弱,肌肉几乎完全松驰;脑电波呈现去同步纶快波。这些表现是异相睡眠期间的基本表现。此外,在异相睡眠期间还会有间断性的阵发性表现,例如眼球出现快速运动、部分躯体抽动,在人类还观察到血压升高和心率加快,呼吸加书本而不规则。
慢波睡眠与异相睡眠是两个相互转化的时相。成年人睡眠一开始首先进入慢波睡眠,慢波睡眠持续约80-120分钟左右后,转入异相睡眠;异相睡眠持续约20-30分钟左右后,又转入慢波睡眠;以后又转入异相睡眠。整个睡眠期间,这种反复转化约4-5次,越接近睡眠后期,异相睡眠持续时间逐步延长。在成年人,慢波睡眠和异相睡眠均可直接转为觉醒状态;但觉醒状态只能进入慢波睡眠。而不能直接进入异相睡眠。在异相睡眠期间,如将其唤醒,被试者往往会报告他正在做梦。据统计,在191例被试者异相睡眠期间唤醒后,报告正在做梦的有152例,占80%左右;在160例被试者慢波睡眠期间唤醒后,报告正在做梦的只有11例,占7%左右。因此一般认为,做梦是异相睡眠的特征之一。
在人体中还观察到,垂体前叶生长激素的分泌与睡眠的不同时相有关。在觉醒状态下,生长激素分泌较小;进入慢波睡眠后,生长激素分泌明显升高;转入异相睡眠后,生长激素分泌又减少。看来,慢波睡眠对促进生长、促进体力恢复是有利的。
异相睡眠是睡眠过程中再现的生理现象,具有一定的生理意义。曾观察到,如几天内被试者在睡眠过程中一出现异相睡眠就将其唤醒,使异相睡眠及时阻断,则被试者会出现易激动等心理活动的扰乱。然后,又让被试者能自然睡眠而不予唤醒,开始几天异相睡眠增加,以补偿前阶段异相睡眠的不足;在这种情况下异相睡眠可直接出现在觉醒之后,而不需经过慢波睡眠阶段。由此认为异相睡眠是政党衙所必需的生理活动过程。动物脑灌流实验观察到,异相睡眠期间脑内蛋白质合成加快。因此认为,异相睡眠对于幼儿神经系统的成熟有密切关系;并认为异想睡眠期间有昨于建立新的突触联系而促进学习记忆活动。看来,异相睡眠对促进精力的恢复是有利的。但是,异相睡眠会出现间断性的阵发性表现,这可能与某些疾病在夜间发作有关,例如心绞痛、哮喘、阻塞性肺气肿缺氧发作等。有人报导,病人在夜间心绞痛发作前常先做梦,梦中情绪激动,伴有呼吸加快、血压升高、心率加快,以致心绞痛发作而觉醒。
(三)睡眠发生的机制
睡眠是由中枢内发生了一个主动过程而造成的,中枢内存在着产生睡眠的中枢,有人认为,在脑干尾端存在能引起睡眠和脑电波同步化的中枢。这一中枢向上传导可作用于大脑皮层(有人称之为上行抑制系统),并与上行激动系统的作用相对抗,从而调节着睡眠与觉醒的相互转化。
由于中枢神经递质研究的进展,已把睡眠的发生机制与不同的中枢递质系统功能联系了起来。慢波睡眠可能与脑干内5-羟色递质系统有关,异相睡眠可能与脑干内5-羟色胺和去甲肾上腺素递质系统有关。
(张镜如)
参考资料
1.徐丰彦,张镜如主编.人体生理学.北京:人民卫生出版社,1989
2.韩济生主编.神经科学纲要.北京:北京医科大学、中国协和医科大学联合出版社1993
3.潘家右,伍嘉宁. NO:中枢神经中一个新的重要信使物质.生理科学进展1993;24:293-297
4.包雅琳,郭 恒怡.M受体亚型研究的新进展。生理科学进展 1992;23:18-22
5.钱家庆.a –肾上腺素受体,生理科学进展1988;19:57-60
6.王崇铨,突触前受体,生理科学进展1989;20:144-147
7.宋亮年,中枢神经系统中受体亚型的多样性及其生物学意义。生理科学进展1992;23:13;17
8.柳川,神经生长因子的研究,生理科学进展1990;21:54-57
9.邱一华,王健鹤,神经递质和神经肽对免疫功能的影响,生理科学进展1992;22:21-24
10.苏正昌,等,免疫反应产物对神经及内分泌系统功能的影响,生理科学进展1992;23:327-331
11.金观源,突触的可塑性与学习、记忆机制,生理科学进展1989;20:148-153
12.BaileyCH,Kandel ER.Structural changes accompanying memory storage.Ann ReyPhysiol1993;55:397-426
13.Bredt DS.Snyder SH.Nitricoxide,a noval neuronal messenger,Neuron 1992;8:3-11
14.Goodman LS,Gilman A.ThePharmacological Basis of Therapeutics 8thed,Pergamon Press,NewYork,1990
15.Hellige JB.Hemisphericasymmetry.Ann Rev Psychol 1990;41:55-80
16.McNaughton BL.The mechanism ofexpression of long-term enhancement of hippocampal synpses.Ann Rev Physiol1993;55:375-369
17.Ochs S. Systems of materialtransport in nerve fibres (axoplasmic transport )related to nerve function andtrophic control.Ann NY Acad Sci 1974;228:202-223
18.Patton HD,Fuchs AF,HilleB,Scher AM,Steiner R.Textbook of Physiology 21sted,WB SaunderCO,Philadelphia,1989
19.Turek FW.Circadian neuralrhythms in mammals.Ann Rev Physiol 1985;47:49-64
20. Walsh K.Neuropsychology,Churchill Livingstone,Edinburgh,1978y
第十一章 内分泌
第一节 概述
内分泌系统是由内分泌腺和分解存在于某些组织器官中的内分泌细胞组成的一个体内信息传递系统,它与神经系统密切联系,相互配合,共同调节机体的各种功能活动,维持内环境相对稳定。
人体内主要的内分泌腺有垂体、甲状腺、甲状旁腺、肾上腺、胰岛、性腺、松果体和胸腺;散在于组织器官中的内分泌细胞比较广泛,如消化首粘膜、心、肾、肺、皮肤、胎盘等部位均存在于各种各样的内分泌细胞;此外,在中枢神经系统内,特别是下丘存在兼有内分泌功能的神经细胞。由内分泌腺或散在内分泌细胞所分泌的高效能的生物活性物质,经组织液或血液传递而发挥其调节作用,此种化学物质称为激素(hormone)。
随着内分泌研究的发展,关于激素传递方式的认识逐步深入。大多数激素经血液运输至远距离的靶细胞而发挥作用,这种方式称为远距分泌(telecring);某些激素可不经血液运输,仅由组织液扩散而作用于邻近细胞,这种方式称为旁分泌(paracrine);如果内分泌细胞所分泌的激素在局部扩散而又返回作用于该内分泌细胞而发挥反馈作用,这种方式称为自分泌(autocrine)。另外,下丘脑有许多具有内分泌功能的神经细胞,这类细胞既能产生和传导神经冲动,又能合成和释放激素,故称神经内分泌细胞,它们产生的激素称为神经激素(neurohormone)。神经激素可沿神经细胞轴突借轴浆流动运送至末梢而释放,这种方式称为神经分泌(neurocrine)(图11-1)。
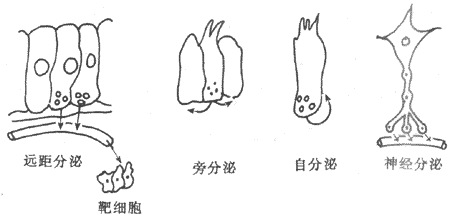
图11-1 激素的递送方式
一、激素的分类
激素的种类繁多,来源复杂,按其化学性质可分为两大类(表11-1):
(一)含氮激素
1.肽类和蛋白质激素 主要有下丘脑调节肽、神经垂体激素、腺垂体激素、胰岛素、甲状旁腺激素、降钙素以及胃肠激素等。
2.胺类激素 包括肾上腺素、去甲肾上腺素和甲状腺激素。
(二)类固醇(甾体)激素
类固醇激素是由肾上腺皮质和性腺分泌的激素,如皮质醇、醛固酮、雌激素、孕激素以及雄激素等。另外,胆固醇的衍生物枣1,25-二羟维生素D3也被作为激素看待。
此外,前列腺素广泛存在于许多组织之中,由花生四烯酸转化而成,主要在组织局部释放,可对局部功能活动进行调节,因此可将前列腺看作一组局部激素。
表11-1 主要激素及其化学性质
| 主要来源 | 激素 | 英文缩写 | 化学性质 |
| 下丘脑 | 促甲状腺激素释放激素 | TRH | 三肽 |
| 促性腺激素释放激素 | GnRH | 十肽 | |
| 生长素释放抑制激素(生长抑素) | GHRIH | 十四肽 | |
| 长征素释放激素 | GHRH | 四十四肽 | |
| 促肾上腺皮制裁激素释放激素 | CRH | 四十一肽 | |
| 促黑(素细胞)激素释放因子 | MRF | 肽 | |
| 促黑(素细胞)激素释放抑制因子 | MIF | 肽 | |
| 催乳素释放因子 | PRF | 肽 | |
| 催乳素释放抑制因子 | PIF | 多巴肽(?) | |
| 升压素(抗利尿激素) | VP(ADH) | 九肽 | |
| 催产素 | OXT | 九肽 | |
| 腺垂体 | 促肾上腺皮持激素 | ACTH | 三十九肽 |
| 促甲状素皮质激素 | TSH | 糖蛋白 | |
| 卵泡刺激素 | FSH | 糖蛋白 | |
| 黄体生长素(间接细胞刺激素) | LH(ICSH) | 糖 蛋白 | |
| 促黑(素细胞)激素 | MSH | 十三肽 | |
| 生长素 | GH | 蛋白质 | |
| 催乳素 | PRL | 蛋白质 | |
| 甲状腺 | 甲状腺素(四碘甲腺原氨酸) | T4 | 胺类 |
| 三碘甲腺原氨酸 | T3 | 胺类 | |
| 甲状腺C细胞 | 降钙素 | CT | 三十二肽 |
| 甲状旁腺 | 甲状旁腺激素 | PTH | 蛋白质 |
| 胰岛 | 胰岛素 | 蛋白质 | |
| 胰高血糖素 | 二十九肽 | ||
| 胰多肽 | 三十六肽 | ||
| 糖皮质激素(如皮质醇) | 类固醇 | ||
| 盐皮激素(如醛固酮) | 类固醇 | ||
| 髓质 | 肾上腺素 | E | 胺类 |
| 去甲肾上腺素 | NE | 胺类 | |
| 睾丸:间质细胞 | 睾酮 | T | 类固醇 |
| 支持细胞 | 抑制素 | 糖 蛋白 | |
| 卵巢、胎盘 | 雌二醇 | E2 | 类固醇 |
| 雌三醇 | E3 | 类固醇 | |
| 孕酮 | P | 类固醇 | |
| 胎盘 | 绒毛膜促性腺激素 | CG | 糖蛋白 |
| 消化道、脑 | 胃泌素 | 十七肽 | |
| 胆囊收缩素-促胰酶素 | CCK-PZ | 三十三肽 | |
| 促胰液素 | 二十七肽 | ||
| 心房 | 心房利尿钠肽 | ANP | 二十一、二十三肽 |
| 松果体 | 褪黑素 | 胺类 | |
| 胸腺 | 胸腺激素 | 肽类 |
二、激素作用的一般特性
激素虽然种类很多,作用复杂,但它们在对靶组织发挥调节作用的过程中,具有某些共同的特点。
(一)激素的信息传递使用
内分泌系统与神经系统一样,是机体的生物信息传递系统,但两者的信息传递形式有所不同。神经信息在神经纤维上传输时,以电信号为信息的携带者,在突触或神经-效应器接头外处,电信号要转变为化学信号,而内分泌系统的信息只是把化学的形式,即依靠激素在细胞与细胞之间进入信息传递。不论是哪种激素,它只能对靶细胞的生理化过程起加强或减弱的作用,调节其功能活动。例如,生长素促进生长发育,甲状腺激素增强代谢过程,胰岛素降低血糖等。在这些作用中,激素既不能添加成分,也不能提供能量,仅仅起着“信使”的作用,将生物信息传递给靶组织,发挥增强或减弱靶细胞内原有的生理化生化近程的作用。
(二)激素作用的相对特异性
激素释放进入血液被运送到全身各个部位,虽然他们与各处的组织、细胞有广泛接触,但有此激素只作用于某些器官、组织和细胞,这称为激素作用的特异性。被激素选择作用的器官、组织和细胞,分别称为靶器官、靶组织和靶细胞。有些激素专一地选择作用于某一内分泌腺体,称为激素的靶腺。激素作用的特异性与靶细胞上存在能与该激素发生特异性结合的受体有关。肽类和蛋白质激素的受体存在于靶细胞膜上,而类固醇激素与甲状腺激素的受体则位于细胞浆或细胞核内。激素与受体相互识别并发生特异性结合,经过细胞内复杂的反应,从而激发出一定的生理效应。有些激素作用的特异性很强,只作用于某一靶腺,如促甲状腺激素只作用于甲状腺,促肾上腺皮质激素只作用于肾上腺皮质,而垂体促性腺激素只作用于性腺等。有些激素没有特定的靶腺,其作用比较广泛,如生长素、甲状腺激素等,它们几乎对全身的组织细胞的代谢过程都发挥调节作用,但是,这些激素也是与细胞的相应受体结合而起作用的。
(三)激素的高效能生物放大作用
激素在血液中的浓度都很低,一般在纳摩尔(n mol/L),甚至在皮摩尔(p mol/L)数量级,虽然激素的含量甚微,但其作用显着,如1mg的甲状腺激素可使机体增加产热量约4200000 J(焦耳)。激素与受体结合后,在细胞内发生一系列酶促放大作用,一个接一个,逐级放大效果,形成一个效能极市制生物放电系统。据估计,一个分子的胰高血糖素使一个分子的腺苷酸环化酶激活后,通过cAMP-蛋白激酶。可激海参10000个分子的磷酸化酶。另外,一个分子的促甲状腺激素释放激素,可使腺垂体释放十万个分子的促甲状腺激素。0.1μg的促肾上腺皮质激素释放激素,可引起腺垂体释放1μg促肾上腺皮持激素,后者能引起肾上腺皮质分泌40μg糖皮质激素,放大了400倍。据此不难理解血中的激素浓度虽低,但其作用却非常明显,所以体液中激素浓度维持相对的稳定,对发挥激素的正常调节作用极为重要。
(四)激素间的相互作用
当多种激素共同参与某一生理活动的调节时,激素与激素之间往往存在着协同作用或拮抗作用,这对维持其功能活动的相对稳定起着重要作用。例如,生长素、肾上腺素、糖皮质激素及胰高血糖素,虽然使用的环节不同,但均能提高血糖,在升糖效应上有协同作用;相反;胰岛素则以降低血糖,与上述激素的升糖效应有拮抗作用。甲状旁腺激素与1,12-二羟维生素D3对血钙的调节是相辅相成的,而降钙素则有拮抗作用。激素之间的协同作用与拮抗作用的机制比较复杂,可以发生在受体水平,也可以发生在受体后信息传递过程,或者是细胞内酶促反应的某一环节。例如,甲状腺激素可使许多组织(如心、脑等)β-肾上腺素能受体增加,提高对儿茶酚胺的敏感性,增强其效应。孕酮与醛固醛在受体水平存在着拮抗作用,虽然孕酮与醛固酮受体的亲和性较小,但当孕酮浓度升高时,则可与醛固酮竞争同一受体,从而减弱醛固酮调节水盐代谢的作用。前列环素(PGI2)可使血小板内cAMP增多,从而抑制血小板聚集;相反,血栓素A2(TXA2)却能使血小板内cAMP减少,促进血小板的聚集。
另外,有的激素本身并不能直接对某些器官、组织或细胞产生生理效应,然而在它存在的条件下,可使另一种激素的作用明显增强,即对另一种激素有调节起支持作用。这种现象称为允许作用(permissive action)。糖皮质激素的允许作用是最明显的,它对心肌和血管平滑肌并元收缩作用,但是,必须有糖皮质激素有存在,儿茶酚胺才能很好地发挥对心血管的调节作用。关于允许作用的机制,至今尚未完全清楚。过去认为,允许作用是由于糖皮质激素抑制儿茶酚-O-甲基移位酶,使儿茶酚胺降解速率减慢,导致儿茶酚胺作用增强。现在通过对受体和受体水平的研究,也可以调节受体介导的细胞内住处传递过程,如影响腺苷酸环化酶的活性以及cAMP的生成等。
三、激素作用的机制
激素作为信息物质与靶细胞上的受体结合后,如何把信息传递到细胞内,并经过怎样的错综复杂的反应过程,最终产生细胞生物效应的机制,一直是内分泌学基础理论研究的重要领域。近一二十年来,随着分子生物学的发展,关于激素作用机制的研究,获得了迅速进展,不断丰富与完善了关于激素作用机制的理论学说。激素按其化学性质分为两在类枣含氮激素和类固醇激素,这两类激素有作用机制也完全不同,现分别叙述。
(一)含氮激素有作用机制枣第二信使学说
第二信使学说是Sutherland等于1965年提出来的。Sutherland学派在研究糖原酵解第一步所需限速酶枣磷酸化酶的活性时,发胰高血糖素与肾上腺素可使肝匀浆在APT、Mg2+与腺苷酸环化酶(adenylate cyclase,AC)的作用下产生一种新物质,这种物质具有激活磷酸体酶从而催化糖原酵解的作用。实验证明,它是环-磷腺苷(cyclic AMP,cAMP),在Mg2+存在的条件下,腺苷酸化酶促进ATPA转变为cAMP。CAMP在磷酸二酯酶(phosphodiesterase)的作用下,降解为5ˊAMP。随后,进一步发现cAMP之所以能激活磷酸化酶,是由于cAMP激活了另一种酶,即依赖cAMP的蛋白激酶(cAMP-dependent protein kinase,cAMP-PK,PKA)而完成的。
Sutherland综合这些资料提出第二信使学说,其主要内容包括:①激素是第一信使,它可与靶细胞膜上具有立体构型的专一性受体结合;②激素与受体结合后,激活漠上的腺苷酸环化酶系统;③在mg2+楔存在的条件下,腺苷酸环化酶促使ATP转变为cAMP,cAMP是第二信使,信息由第一信使传递给第二信使;④cAMP是使无活性的蛋白激酶(PKA)激活。PKA具有两个亚单位,即调节亚单位与催化亚单位。CAMP与PKA的调节亚单位结合,导致调节亚单位与催化亚脱离而使PKA激活,催化细胞内多种蛋白质发生磷酸化反应,包括一些蛋白发生磷酸化,从而引起靶细胞各种生理生化反应(图11-2)。
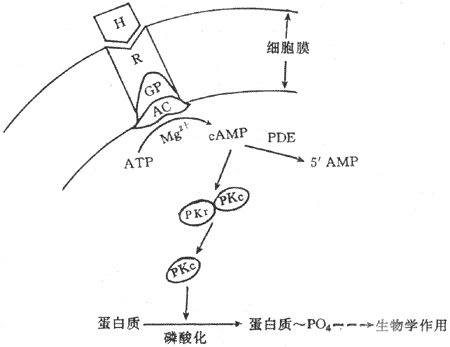
图11-2 含氧激素作用机制示意图
H:激素 R:受体 GP:G蛋白 AC:腺苷酸环化酶PDE:磷酸 二酯酶
RKr :蛋白激酶调节亚单位 PKc:蛋白激酶催化亚单位
以cAMP为第二信使学说的指出,推动了激素作用机制的研究工作迅速深入发展。近年来的研究资料表明,cAMP并不是唯一的第二信使,可能作为第二信使的化学物质还有cGMP、三磷酸肌醇、二酰甘油、Ca2+等。另外,关于细胞表现受体调节、腺苷酸环化酶活化机制、蛋白激酶C的作用等方面的研究都取得了很大进展,现概述如下:
1.激素与受体的相互作用 激素有膜受体多为糖蛋白,其结构一般分为三部分:细胞膜外区段、质膜部分和细胞膜内区段。细胞膜外区段含有许多糖基,是识别激素并与之结合的部位。激素分子和靶细胞受体的表现,均由许多不对称的功能基团构成极为复杂而又可变的立体构型。激素和受体可以相互诱导而改变本身的构型以适应对方的构型,这就为激素与受体发生专一性结合提供了物质基础。
激素与受体的结合力称为亲和力(affinity)。一般来说,由于相互结合是激素作用的第一步,所以亲和力与激素的生物学作用往往一致,但激素的类似物可与受体结合而不表现激素的作用,相反却阻断激素与受体相结合。实验证明,亲和力可以随生理条件的变化而发生改变,如动物性周期的不同阶段,卵巢颗粒细胞上的的卵泡刺激素(FSH)受体的亲和力是不相同的。某一激素与受体结合时,其邻近受体的亲和力也可出现增高或降低的现象。
受体除表现亲和力改变外,其数量也可发生变化。有人用淋巴细胞膜上胰岛素受体进行观察发现,如长期使用大剂量的胰岛素,将出现胰岛素受体数量减少,亲和力也降低;当把胰岛素的量降低后,受体的数量和亲和力可恢复正常。许多种激素(如促甲状腺激素、绒毛膜促性腺激素、黄体生成素、卵泡刺激素等)都会出现上述情况。这种激素使其特异性受体数量减少的现象,称为减衰调节或简称下调(down regulation0)。下调发生的机制可能与激素-受体复合物内移入胞有关。相反,有些激素(多在剂量较小时)也可使其特异性受体数量增多,称为上增调节或简称上调(up regulation),如催乳素、卵泡刺激素、血管紧张素等都可以出现上调现象。下调或上调现象说明,受体或上调现象说明,受体的合成与降解处于动态平衡之中,其数量是这一平衡的结果,它的多少与激素有量相适应,以调节靶细胞对激素有敏感性与反应强度。
2.G蛋白在信息传递中的作用激素受体与腺苷酸环化酶是细胞膜上两类分开的蛋白质。激素受体结合的部分在细胞膜的外表面,而腺苷环化酶在膜的胞浆面,在两者之间存在一种起耦联作用的调节蛋白——鸟苷酸结合蛋白(guanine nucleotide-bindingregulatory protein),简称G蛋白。G蛋白由α、β和γ三个亚单位组成,α亚单位上有鸟苷酸结合位点。当G蛋白上结合的鸟苷酸为GTP时则激活而发挥作用,但当G蛋白上的GTP水解为GDPA时则失去活性。当激素与受体结合时,活化的受体便与G蛋白的α亚单位结合,并促使其与β、γ亚单位脱离,才能对腺苷酸环化酶起激活或抑制作用。
G蛋白可分为兴奋型G蛋白(Gs)和抑制型G蛋白 Gi。Gs的作用是激活腺苷酸环化酶,从而使cAMP生成增多;Gi的作用则是抑制腺苷酸环化酶的活性,使cAMP生成减少。有人提出,细胞膜的激素受体也可分为兴奋型(Rs)与抑制型(Ri)两种,它们分别与兴奋性激素(Hs)或抑制性激素(Hi)发生结合,随后分别启动Gs或Gi,再通过激活或抑制腺苷酸环化酶使cAMP增加或减少而发挥作用。
3.三磷酸肌醇和二酰甘油为第二信使的信息传递系统许多含氮激素是以cAMP为第二信使调节细胞功能活动的,但有些含氮激素的作用信息并不以cAMP为媒介进行传递,如胰岛素、催产素、催乳素、某些下丘脑调节肽和生长因子等。实验证明,这些激素作用于膜受体后,往往引起细胞膜磷脂酰肌醇转变成为三磷酸肌醇(inositol-1,4,5,triphosphate,IP3)和二酰甘油(diacylglycerol,DG),并导致胞浆中Ca2+浓度增高。近年来,有人提出IP3和DG可能是第二信使的学说引起人们的重视,并且得到越来越我的实验证实。这一学说认为,在激素的作用下,可能通过G蛋白的介导,激活细胞膜内的磷脂酶C(phosphinositol-specificphospholipase C.PLC),它使由磷脂酰肌醇(PI)二次磷酸化生成的磷脂酰二磷肌醇(PIP2)分解,生成IP3和DG。DG生成后仍留在膜中,IP3则进入胞浆。在未受到激素作用时,细胞膜几乎不存在游离的DG,细胞内IP3的含量也极微,只有在细胞3受到相应激素作用时,才加速PIP2的降解,大量产生IP3和DG。IP3的作用是促使细胞内Ca2+贮存库释放Ca2+进入胞浆。细胞内Ca2+主要贮存在线粒体与内质网中。实验证明,IP3引起Ca2+的释放是来自内质网而不是线粒体,因为在内质网膜上有IP3受体,IP3与其特异性受体结合后,激活Ca2+通道,使Ca2+从内质网中进入胞浆。IP3诱发Ca2+动员最初发反应是引起暂短的内质网释放Ca2+,随后是由Ca2+释放诱发作用较长的细胞外Ca2+内流,导致胞浆中Ca2+浓度增加。Ca2+与细胞内的钙调蛋白(calmodulin,CaM)结合后,可激活蛋白酶,促进蛋白质磷酸化,从而调节细胞的功能活动。
DG的作用主要是它能特异性激活蛋白激酶C(protein kinase C,PKC)PKC的激活依赖于Ca2+的存在。激活的PKC与PKA一样可使多种蛋白质或酶发生磷酸化反应,进而调节细胞的生物效应。另外,DG的降解产物花生四烯酸是合成前列腺素的原料,花生四烯酸与前列腺素的过氧化物又参与鸟苷酸环化酶的激活,促进cGMP的生成。CGMP作为另一种可能的第二信使,通过激活蛋白激酶G(PKG)而改变细胞的功能(图11-3)。
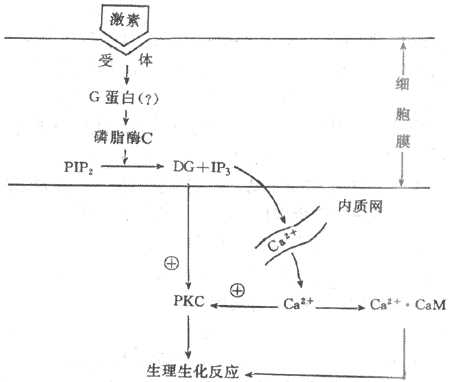
图11-3 磷脂酰肌醇信息传递系统示意图
PIP2:磷脂酰二磷肌醇 DG:二酰甘油 IP3:三磷酸肌醇PKC:蛋白激酶C CaM:钙调蛋白
(二)类固醇激素作用机制枣基因表达学说
因固醇激素的分子小(分子量仅为300左右)、呈脂溶性,因此可透过细胞膜进入细胞。在进入细胞之后,经过两个步骤影响基因表面而发挥作用,故把此种作用机制称为二步作用原理,或称为基因表达学说。
第一步是激素与胞浆受体结合,形成激素-胞浆受体复合物。在靶细胞将中存在着类固醇激素受体,它们是蛋白质,与相应激素结合特点是专一性强、亲和性大。例如,子宫组织胞浆的雌二醇受体能与17β-雌二醇结合,而不能与17α-雌二醇结合。激素与受体的亲和性大小与激素的作用强度是平行的。而且胞浆受体的含量也随靶器官的功能状态的变化而发生改变。当激素进入细胞内与胞浆受体结合后,受体蛋白发生构型变化,从而命名激素-胞浆受体复合物获得进入核内的能力,由胞浆转移至核内。第二步是与核内受体相互结合,形成激素-核受体复合物,从而激发DNA的转录过程,生成新的mRNA,诱导蛋白质合成,引起相应的生物效应(图11-4)。
图11-4 类固醇激素作用机制示意图
近年来由于基因工程技术的发展与应用,不少类固醇激素的核内受体的结构已经清楚。它们是特异地对转录起调节作用的蛋白,其活性受因固醇激素的控制。核受体主要有三个功能结构域:激素结合结构域、DNAA结构结构域和转录增强结构域。一旦激素与受体结合,受体的分子构象发生改变,暴露出隐蔽于分子内部的DNA结合结构域及转录增强结构域,使受体DNA结合,从而产生增强转录的效应。另外,政治家实验资料表明,在DNA结合结构域可能有一个特异序列的氨基酸片断,它起着介导激素受体复合物与染色质中特定的部位相结合,发挥核定位信号的作用。
甲状腺激素虽属含氮激素,但其作用机制却与类固醇激素相似,它可进入细胞内,但不经过与胞浆受体结合即进入核内,与核受体结合调节基因表达。
应该指出,含氮激素可作用于转录与翻译阶段而影响蛋白质的合成;反过来,类固醇激素也可以作用于细胞膜引起基因表达学说难以解释的某引起现象。
第二节 下丘脑的内分泌功能
下丘脑与神经垂体和腺垂体的联系非常密切,如视上核和室旁核的神经元轴突延伸终止于神经垂体,形成下丘脑-垂体束。在下丘脑与腺垂体之间通过垂体门脉系统发生功能联系。下丘脑的一些神经元既能分泌激素(神经激素),具有内分泌细胞的作用,又保持典型神经细胞的功能。它们可将从大脑或中枢神经系统其他部位传来的神经信息,转变为激素的信息,起着换能神经元的作用,从而以下丘脑为枢纽,把神经调节与体液调节紧密联系起来。所以,下丘脑与垂体一起组成下丘脑-垂体功能单位(图11-5)。
凡是能分泌神通肽或肽类激素的神经分泌细胞称为肽能神经元。下丘脑的肽能神经元主要丰硕盱视上核、室旁核与促垂体核团。促垂体区核团位于下丘脑的内侧基底部,主要包括正中隆起、弓状核、腹内侧核、视交叉上核以及室周核等,多属于小细胞肽能神经元,其轴突投射到正中隆起,轴突末梢与垂体门脉系统的第一级毛细血管风接触,可将下丘脑调节肽释放进入门脉系统,从而调节垂体的分泌活动。
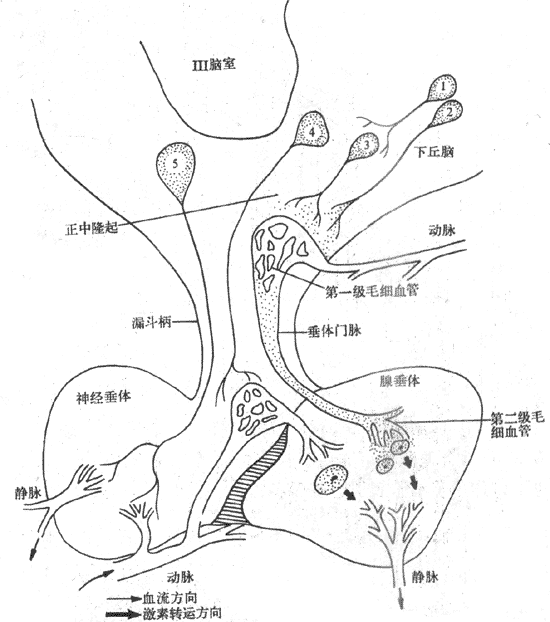
图11-5 下丘脑-垂体功能单位
1:单胺能神经元 2、3、4、5:为下丘脑各类肽能神经元
一、下丘脑调节肽
下丘脑促垂体区肽能神经元分泌的肽类激素,主要作用是调节腺垂体的活动,因此称为下丘脑调节肽(hypothalamus regulatory peptide,HRP)。近20多年来,从下丘脑组织提取肽类激素获得成功,并已能人工合成。1968年Guillemin实验室从30万只羊的下丘脑中成功地分离出几毫克的促甲状腺激素释放激素(TRH),并在一年后确定其化学结构为三肽。在这一生成成果鼓舞下,Schally实验室致力于促性腺激素释放激素(GnRH)的提取工作。1971年他们从16万头猪的下丘脑中提纯出GnRH,又经过6年的研究,阐明其化学结构为十肽。此后,生长素释放抑制激素 (GHRIH)、促肾上腺皮质激素释放激素(CRH)与生长素释放激素(GHRH)相继分离成功,并确定了化学结构,此外,还有四种对腺垂体催乳素和促黑激素的分泌起促进或抑制作用的激素,因尚未弄清其化学结构,所以暂称因子。
下丘脑调节肽除调节腺垂体功能外,它们几乎都具有垂体外作用,而且它们也不仅仅在下丘脑“促垂体区”产生,还可以大中枢神经系统其他部位及许多组织中找到它们踪迹,使人们更加广泛深入地研究他们的作用。
(一)促甲状腺激素释放激素
促甲状腺激素释放激素(thyrotropin-releasinghormone,TRH)是三肽,其化学结构为:
(焦)谷-组-脯-NH2
TRH主要作用于腺垂体促进促甲状腺激素(TSH)释放,血中T4和T3随TSH浓度上升而增加。给人和动物静脉注射TRH(1mg),1-2min内血浆TSH浓度便开始增加,10-20min达高峰,TSH的含量可增加20倍。腺垂体的促甲状腺激素细胞的膜上的TRH受体,与TRH结合后,通过Ca2+介导引起TSH释放,因此IP3-DG系统可能是TRH发挥作用的重要途径。TRH除了刺激腺垂体释放TSH外,也促进催乳互的释放,但TRH是否参与催乳素分泌的生理调节,尚不能肯定。
下丘脑存在大量的TRH神经元,它们主要分布于下丘脑中间基底部,如损毁下丘脑的这个区域则引起TRH分泌减少。TRH神经元合成的TRH通过轴浆运输至轴突末梢贮存,延伸到正中隆起初级毛细血管周围的轴突末梢在适当刺激作用下,释放TRH并进入垂体门脉系统运送到腺垂体,促进TRH释放。另外,在第三脑室周围尤其是底部排列有形如杯状的脑室膜细胞(tanycyte),其形态特点与典型的脑室膜细胞有所不同,其胞体细长,一端面向脑室腔,其边界上无纤毛而有突起,另一端则延伸至正中隆起的毛细血管周围。在这些细胞内含有大量的TRH与GnRH等肽类激素。下丘脑特别是室周核释放的TRH或GnRH进入第三脑室的脑脊液中,可被脑室膜细胞摄入,再转幸福至正中隆起附近释放,然后进入垂体门脉系统。
除了下丘脑有较多的TRH外,在下丘脑以外的中枢神经部位,如大脑和脊髓,也发现有TRH存在,其作用可能与神经信息传递有关。
(二)促性腺激素释放激素
促性腺激素释放激素(gonadotropin-releasinghormone,GnRH,LRH)是十肽激素,其化学结构为:
(焦)谷-组-色-丝-酪-甘-亮-精-脯-甘-NH2
GnRH促进性腺垂体合成与释放促性腺激素。当机体静脉注射100mgGnRH,10min后血中黄体生成素(LH)与卵泡刺激素(FSH)浓度明显增加,但以LH的增加更为显着。在体外腺垂体组织培养系统中加入GnRH,亦能引起LH与FSH分泌增加,如果先用GnRH抗血清处理后,再给予GnRH,则可减弱或消除GnRH的效应。
下丘脑释放GnRH的特脉冲式释放,因而造成血中LH与FSH浓度也呈现脉冲式波动。从恒河猴垂体门脉血管收集的血样测定GnRH含量,呈现阵发性时高时低的现象,每隔1-2h波动一次。在大鼠,GnRH每隔20-30min释放一次,如果给大鼠注射抗GnRH血清,则血中LH与FSH浓度的脉冲式波动消失,说明血中LH与FSH的脉冲式波动是由下丘脑GnRH脉冲式释放决定的。用青春期前的幼猴实验表明,破坏产生GnRH的弓状核后,连续滴注外源的GnRH并不能诱发青春期的出现,只有按照内源GnRH所表现的脉冲式频率和幅度滴注GnRH,才能使血中LH与FSH浓度呈现类似正常的脉冲式波动,从而激发青春期发育。看来,激素呈脉冲式释放对发挥其作用是十分重要的。
腺垂体的促性腺激素细胞的膜上有GnRH受体,GnRH与其受体结合后,可能是通过磷脂酰肌醇信息传递系统导致细胞内Ca2+浓度增加而发挥作用的。
在人的下丘脑,GnRH主要集中在弓状核、内侧视前区与室旁核。除下丘脑外,在脑的其他区域如间脑、边缘叶,以及松果体、卵巢、睾丸、胎盘等组织中,也存在着GnRH。GnRH对性腺的直接作用则是抑制性的,特别是药理剂理的GnRH,其抑制作用更为明显,对卵巢可抑制卵泡发育和排卵,使雌激素与孕激素生成减少;对睾丸则抑制精子的生成,使睾酮的分泌减低。
(三)生长抑素与生长素释放激素
1.生长抑素(生长素释放抑制素,growthhormone release-inlease-inhibiting hormone,GHRIH,或somatostatin)是由116个氨基酸的大分子肽裂解而来的十四肽,其分了结构呈环状,在第3位和第14位半胱氨酸之间有一个二硫键,其化学结构为:

生长抑素是作用比较广泛的一种神经激素,它的主要作用是抑制垂体生长素(GH)的基础分泌,也抑制腺垂体对多种刺激所引起的GH分泌反应,包括运动、进餐、应激、低血糖等。另外,生长抑素还可抑制LH、FSH、TSH、PRL及 ACTH的分泌。生长抑素与腺垂体生长素细胞的膜受体结合后,通过减少细胞内cAMP和 Ca2+而发挥作用。
除下丘脑外,其他部位如大脑皮层、纹状体、杏仁核、海马,以及脊髓、交感神经、胃肠、胰岛、肾、甲状腺与甲状旁腺等组织广泛存在生长抑素。在脑与胃肠又纯化出28个氨基酸组成的在GHRIH28,它是GHRIH14N端向外延伸而成。生长抑素的垂体外作用比较复杂,它在神经系统可能起递质或调质的作用;生长抑素对胃肠运动与消化道激素的分泌均有一定的抑制作用;它还抑制胰岛素、胰高血糖素、肾素、甲状旁腺激素以及降钙素的分泌。
2.生长素释放激素(growthhormone releasing hormone,GHRHA)由于下丘脑中GHRH的含量极少,致化学提取困难。1982年有人首先从一例患胰腺癌伴发肢端肥大症患者的癌组织中提取并纯化出一种44个氨基酸的肽,它在整体和离体实验均显示有促GH分泌的生物活性。1983年,从大鼠下丘脑中提纯了GHRH43,这种四十三肽对人的腺垂体也有很强有促GH分泌作用。近年用DNA重组扶得到GHRH40和GHRH44的基因,这些基因已被克隆化,并非酵母系统中传代和表达,为提供充足与兼价的GHRH开拓了可喜的前景。
产生GHRH的神经元主要分布在下丘脑弓状核及腹内侧核,它们的轴突投射到正中隆起,终止于垂体门脉初级毛细血管旁。GHRH呈脉冲式释放,从而导致腺垂体的GH分泌也呈现脉冲式。大鼠实验证明,注射GHRH抗体后,可消除血中GH浓度的脉冲式波动。一般认为,GHRH是GH分泌的经常性调节者,而GHRIH则是在应激刺激GH分泌过多时,才显著地发挥对GH分泌的抑制作用。GHRH与GHRIH相互配合,共同调节腺垂体GH的分泌。
在腺垂体生长素细胞的膜上有GHRH受体,GHRH与其受体结合后,通过增加内cAMP与Ca2+促进GH释放。
(四)促肾上腺皮质激素释放激素
促肾上腺皮质激素释放激素(corticotropin releasinghormone,CRH)为四十一肽,其主要作用是促进腺垂体合成与释放促肾上腺皮质激素(ACTH)。腺垂体中存在大分子的促阿片-黑素细胞皮质素原(pro-opiomelanocortin,POMC),简称阿黑皮素原。在CRHA作用下经酶分解了ACTH、溶脂激素(lipotropin,β-LPH)和少量的β-内啡肽。静脉注射CRH5-20min后,血中ACTH浓度增加5-20倍。
分泌CRH的神经元主要分布在下丘脑室旁核,其轴突多投射到正中隆起。在下丘脑以外部位,如杏仁核、海马、中脑,以及松果体、胃肠、胰腺、肾上腺、胎盘等处组织中,均发现有CRH存在。下丘脑CRH以脉冲式释放,并呈现昼夜周期节律,其释放量在6-8点钟达高峰,在0点最低。这与ACTH及皮质醇的分泌节律同步。机体遇到的应激刺激,如低血溏、失血、剧痛以及精神紧张等,作用于神经系统不同部位,最后将信息汇集于下丘脑CRH神经元,然后通过CRH引起垂体-肾上腺皮质系统反应。
CRH与腺垂体促肾上腺皮质激素细胞的膜上CRH受体结合,通过增加细胞内cAMP与Ca2+促进ACTH的释放。
(五)催乳素释放抑制因子与催乳素释放因子
下丘脑对腺垂体催乳素(PRL)的分泌有抑制和促进两种作用,但平时以抑制作用为主。首先在哺乳动物下丘脑提取液中,发现一种可抑制腺垂体释放PRL的物质,称为催乳素释放抑制因子(prolactinrelease-inhibiting factor,PIF)。随后,又在下丘脑提取液中发现还有一咱能促进腺垂体释放PRL的因子,称为催乳素释放因子(prolactin releasingfactor,PRF)。将下丘脑提取液中的TRH分离出去,仍具有PRF活性,说明下丘脑提取液中PRF活性不是来自TRH。PIF与PRF的化学结构尚不清楚,由于多巴肽可直接抑制腺垂体PRL分泌,注射多巴胺可使正常人或高催乳素血症患者血中的PRL明显下降,而且在下丘脑和垂体存在的多巴胺,因此有人进出多巴胺可能就是PIF的观点。
(六)促黑素细胞激素释放因子与抑制因
促黑素细胞激素释放因子(melanophore-stimulatinghormone releasing factor,MRF)(melanophore-stimulatinghormone release-inhibiting factor,MIF)可能是催产素裂解出来的两种小分子肽。MRF促进MSH的释放,而MIF则抑制MSH的释放。
二、调节下丘脑肽能神经元活动的递质
下丘脑能神经元与来自其他部位的神经纤维有广泛的突触联系,其神经递质比较复杂,可分为两大类:一类递质是肽类物质,如脑啡肽、β-内啡肽、神经降压素、P物质、血管活性肠肽及胆囊收缩素等;另一类递质是单胺类物质,主要有多巴胺(DA)、去甲肾上腺素(NE)与 5-羟色胺(5-HT)。
组织化学研究表明,三种单受类递质的浓度,以下丘脑“促垂体区”正中隆起附近最高。单胺能神经元可直接与释放下丘脑调节肽的肽能神经元发生突触联系,也可以通过多突触发生联系。单胺能神经元通过释放单胺类递质,调节肽能神经元的活动。下丘脑单受能神经元的活动不断受中枢神经系统其他部位的影响,所以它们对下丘脑调节肽分泌的调节作用比较复杂,现将一些研究结果列于表11-2。
表11-2 单胺类递质对几种下丘脑调节肽和相关激素分泌的影响
| TRH(TSH) | GnRH(LH、FSH) | GHRH(GH) | CRH(ACTH) | PRF(PRL) | |
| NE | ↑ | ↑ | ↑ | ↓ | ↓ |
| DA | ↓ | {4}↓(一) | ↑ | ↓ | ↓ |
| 5-HT | ↓ | ↓ | ↑ | ↑ | ↑ |
↑增加分泌 ↓减少分泌 (一)不变
近年来的研究表明,阿片肽对下丘脑调节肽的释放有明显的影响。例如,给人注射脑啡肽或β-内啡肽可抑制CRH的释放,从布使ACTH分泌减少,而纳洛酮则有促进CRH释放的作用;注射脑啡肽或β-内啡肽可刺激下丘脑释放TRH和GHRH,使腺垂体分泌TSH与GH增加,而对下丘脑的GnRH释放则明显的抑制作用。
第三节 垂体
垂体按其胚胎发育和功能、形态的不同,分为腺垂体和神经垂体两部分。腺垂体来自胚胎口凹的外胚层上皮,是由6种腺细胞组成的上皮细胞。神经垂体来自间脑底部的漏斗,主要由下丘脑-垂体束的无髓神经纤维和神经胶质细胞分化而成的神经垂体细胞组成。垂体以漏斗与下丘脑相连。由于在形成与功能上下丘脑与垂体的联系非常密切,可将它们看作一个功能单位。
腺垂体与神经垂体的内分泌功能迥然不同,现分别叙述。
一、腺垂体
腺垂体是体内最重要 的内分泌腺。它由不同的腺细胞分泌七种激素:由生长素细胞分泌生长素(GH);由促甲状腺激素细胞分泌促甲状腺激素(TSH);由促肾上腺皮质激素细胞分泌促肾上腺皮质激素(ACTH)与促黑(素细胞)激素(MSH);由促性腺激素细胞分泌卵泡刺激素(FSH)与共同体生成素(LH);由催乳素细胞分泌催乳素(PRL)。在腺垂体分泌的激素中,TSH、ACTH、FSH与LH均有各自的靶腺,分别形成:①下丘脑-垂体-甲状腺轴;②下丘脑-垂体-肾上腺皮质轴;③下丘脑-垂体-性腺轴。腺垂体的这些激素是通过调节靶腺的活动而发挥作用的,而GH、PRL与MSH则不通过靶腺,分别直接调节个体生长、乳腺发育与泌乳、黑素细胞活动等。所以,腺垂体激素的作用极为广泛而复杂。
(一)生长素
人生长素(human growth hormone,hGH)含有191个氨基酸,分子量为22000,其化学结构与会催乳素近似,故生长素有弱催乳素作用,而催乳素有弱生长素作用。不同种类动物的生长素,其化学结构与免疫性质等有较大差别,除猴的生长素外,其他动物的生长素对人无效。近年利用DNA重组技术可以大量生产hGH,供临床应用。人GH的化学结构见图11-6。
1.生长素的作用 GH的生理作用是促进物质代谢与生长发育,对机体各个器官与各种组织均有影响,尤其是骨骼、肌肉及内脏器官的作用更为显着,因此,GH也称为躯体刺激素(somatotropin)。
(1)促进生长作用:机体生长受多种激素的影响,而GH是起关键作用的调节因素。幼年动物摘除垂体后,生长即停止,如及时补充GH则可使其生长恢复。人幼年时期GH,将出现生长停滞,身材矮小,称为侏儒症;如GH过多则患巨人症。人成年后GH过多,由于长骨骨骺已经钙化,长骨不再生长,只能使软骨成分较多的手脚肢端短骨、面骨及其软组织生长异常,以致出现手足粗大、鼻大唇厚、下颌突出等症状,称为肢端肥大症。正常成年男子在空腹安静状态下,血浆中GH浓度不超过5μg/L,成年女子不超过10μg/L。而巨人症与肢端肥大症患者血中GH浓度可明显增高。
图11-6 人生长素的化学结构(黑柱处代表二硫键)
GH的促生长作用是由于它能促进骨、软骨、肌肉以及其他组织细胞分裂增殖,蛋白质合成增加,离体软骨培养实验发现,将GH加入到去垂体动物的软骨培养液中,对软骨的生长无效,而加入正常动物的血浆却有效,说明GH对软骨的生长并无直接作用,而在正常动物血浆中存在某种有促进生长作用的因子。实验研究证明,GH主要诱导肝产生一种具有促生长作用的肽类物质,称为生长介素(somatomedin,SM),因其化学结构与胰岛素看近似,所以又称为胰岛素样生长因子(insulin-like growth factor,IGF)。目前已分离出两种生长介素,即IGF-I和IGF-Ⅱ,它们分子组成的氨基酸有70%是相同的。IGF-I是含有70个氨基酸的多肽,GH的促生长作用主要是通过IGF-I作介导的。IGF-Ⅱ是含有67个氨基酸的多肽,它主要在胚胎期产生,对胎儿的生长起重要作用。血液中的IGF-I含量信号2于GH的水平,摘除垂体的大鼠血中IGF-I含量降低,注射GH后,血中IGF-I含量增加,并与GH的剂量呈依赖式。活动期肢端肥大症患者血中IGF-I含量明显增高而侏儒症患者血中IGF-I含量明显低于正常。给人注射GH,往往需要12-18h后,血中IGF-I含量才会升高,所以当血中GH浓度有急剧变化时,在一定时间内血中IGF-I的含量可维持相对稳定,在青春期,随着GH分泌增多,血中IGF-I的浓度也相应增加。
给幼年动物注射生长介素能明显刺激动物生长,身长增高,体重增加,IGF-Ⅱ比IGF-I的促生长作用更强。生长介素主要的作用是促进软骨生长,它除了可促进硫酸盐进入软髓组织外,还促进氨基酸进入软骨细胞,增强DNA、RNA和蛋白质的合成,促进软骨组织增殖与骨化,使长骨加长。
血中的生长介素,绝大部分与生长介素结合蛋白结合,被运送到全身各处除肝外,肌肉、肾、心与肺等组织也能产生生长介素,可能以旁分泌的方式,以局部起作用。
(2)促进代谢作用:GH可通过生长介素促进氨基酸进入细胞,加速蛋白质合成,包括软骨、骨、肌肉、肝、肾、心、肺、肠、脑以皮肤等组织的蛋白质合成增强;GH促进脂肪分解,增强脂肪酸氧化,抑制外周组织摄取与利用葡萄糖,减少葡萄糖的消耗,提高血糖水平。GH对脂肪与糖代谢的作用似乎与生长介素无关,机制尚不清楚。
近年研究证明,血中的生长介互可对GH分泌有负反馈调节作用。IGF-I能刺激下丘脑释放GHRIH,从而抑制GH的分泌。IGF-I还能直接抑制培养的腺垂体细胞GH的基础分泌和GHRH刺激的GH分泌,说明IGF-I可通过下丘脑和垂体两下水平对GH分泌进入负反馈调节。
除了上述的调控机制外,还有许多因素可以影响GH的分泌:
(1)睡眠的影响:人在觉醒状态下,GH分泌较少,进入慢波睡眠后,GH分泌明显增加,约在60min左右,血中GH浓度达到高峰。转入异相睡眠后,GH分泌又减少。看来,在慢波睡眠其GH分泌增多,对促进生长和体力恢复是有利的。50岁以后,GH这种分泌峰消失。
(2)代谢因素的影响:血中糖、氨基酸与脂肪酸均能影响GH的分泌,其中以低血糖对GH分泌的刺激作用最强。当静脉注射胰岛素使血糖降至500mg/L以下时,经30-60min,血中GH浓度增加2-10倍。相反,血糖升高可使GH浓度降低。有人认为,在血糖降低时,下丘脑GHRH神经元兴奋性提高,释放GHRH增多,GH分泌增加,可减少外周组织对葡萄糖的利用,而脑组织对葡萄糖的利用可基本不受影响。血中氨基酸与脂肪酸增多可引起GH分泌增加,有利于机体对这些物质的代谢与利用。
此外,运动、应激刺激、甲状腺激素、雌激素与睾酮无法能促进GH分泌。在青春其,血中雌激素或睾酮浓度增高,可明显地增加GH分泌,这是在期GH分泌较多的一个重要因素。
(二)催乳素
催乳素(prolactin,PRL)是含199个氨基酸并有三个二硫键的多肽,分子量为22000。在血中还存在着较大分子的PRL,可能是PRL的前体或几个PRLA分子的聚合体,成人血浆中的PRL浓度<20μg/L。
PRL的作用极为广泛,下面仅就其主要作用加以扼要说明。
1.对乳腺的作用 PRL引起并维持泌乳,故名催乳素。在女性青春期乳腺的发育中,雌激素、孕激素、生长素、皮质醇、胰岛素、甲状腺激素及PRL起着重要的作用。到妊娠期,PRL、雌激素与孕激素分泌增多,使乳腺组织进一步发育,具备泌乳能力却不泌乳,原因是此时血中雌激素与孕激素浓度过高,抑制PRL的泌乳作用。分娩后,血中的雌激素和孕激素浓度大大降低,PRL才能地发挥始动和维持泌乳的作用。在妊娠期PRL的分泌显著增加,可能与雌激素刺激垂体催乳素细胞的分泌活动有关。妇女授乳时,婴儿吸吮乳头反射性引起PRL大量分泌。
2.对性腺的作用 在哺乳类运物,PRL对卵巢的黄体功能有一定的作用,如啮齿类,PRL与LH配合,促进黄体形成并维持分泌孕激素,但大剂量的PRL又能使黄体溶解。PRL对人类的卵巢功能也有一定的影响,随着卵泡的发育成熟,卵泡内的PRL含量逐渐增加,并在次级留言簿包发育成为排卵前卵泡的过程中,在颗粒细胞上出现PRL受体,它是在FSH的刺激下形成的。PRL与其受体结合,可刺激LH受体生成,LH与其受体结合后,促进排卵、黄体生成及孕激素与雌激素的分泌。实验表明,小量的PRL对卵巢激素与孕激素的合成起允许作用,而大量的PRL则有抑制作用。临床上患闭经溢乳综合症的妇女,表现特征为闭经、溢乳与不孕,患者一般都存在无排卵与雌激素水平低落,而血中PRL浓度却异常增高。
男性在睾酮存在的条件下,PRL促进前列腺及精囊腺的生长,还可以增强LH对间质细胞的应用,使睾酮的合成增加。
PRL参与反激反应。 在应激状态下,血中PRL浓度升高,而且往往与ACTH和GH浓度的增高一出现,刺激停止数小时后才逐渐恢复到正常水平。看来,PRL可能与ACTH及GH一样,是应激反应中腺垂体分泌的三大激素之一。
腺垂体PRL的分泌受下丘脑PRF与PIF的双重控制,前者促进PRL分泌,而执行者则抑制其分泌。多巴胺通过下丘脑或直接对腺垂体PRL分泌有抑制作用。下丘脑的TRH能促进PRL的分泌。吸吮乳头的刺激引起传入神经冲动,经脊髓上传至下丘脑,使PRF神经元发生兴奋,PRF释放增多,促使腺垂体分泌PRL增加,这是一个典型的神经内分泌反射。
腺垂体分泌的促激素(TSH、ACTH、LH、FSH)在有关章节中叙述。
二、神经垂体
神经垂体不含腺体细胞,不能合成激素。所谓的神经垂体激素是指在下丘脑视上核、室旁核产生而贮存于神经垂体的升压素(抗利尿激素)与催产素,在适宜的刺激作用下,这两种激素由神经垂体释放进入血液循环。
升压素(vasopressin, VP或antidiuretic hormone,ADH)与催产素(oxytocin,OXT)在下丘脑的视上核与室旁核均可产生,但前者主要在视上核产生,而后者主要在室旁核产生。它们的化学结构都是九肽,催产素与升压素只是第3位与第8位的氨基酸残基有所不同(图11-7)。人升压素的第8位氨基酸为精氨酸,故称为精氨酸升压不比(argininevasopressin,AVP)。这两种激素已能人工合成。
图 11-7 催产素与升压素的化学结构
实验证明,升压素与催产素是在视上核和室旁核神经元的核蛋白体上先形成激素有前身物质(激素原),再裂解成神经垂体激素的,并与同时合成的神经垂体激素运载蛋白(neurophysin)形成复合物,包装于囊泡中,呈小颗粒状。在轴突内,囊泡以每天2-3mm的速度运送至神经垂体。在适宜刺激的作用下,视上核或室旁核发生兴奋,神经冲动将沿着下丘脑-垂体束传导至神经垂体中的神经末梢,使其发生去极化,导致Ca2+内流进入末梢内,促进末梢的分泌囊泡经出泡作用而将神经垂体激素与其运载蛋白一并释放进入血液。
神经垂体激素运载蛋白有两种:一种与催产素结合释放入血液的,称为运载蛋白I,由92个氨基酸组成;另一种与升压素结合的称为运载蛋白Ⅱ,由97个氨基酸组成,烟碱可使血浆中运载蛋白Ⅱ和升压素浓度同时升高,而雌激素可使血浆中运载蛋白I含量增加,而催产素浓度并不随之增加。
有资料表明,神经垂体激素不仅存在于下丘脑-垂体束系统内,而且在下丘脑正中隆起与第三脑室附近的神经元轴突中也有神经垂体激素。在大鼠和猴的垂体门脉血液中,检测出大量的升压素,其浓度远远主于外周血液中的浓度,而且注射大量的升压素能引起腺垂体ACTH分泌增加,提示神经垂体激素可能影响垂体的分泌活动。
(一)升压素(抗利尿激素)
血浆中升压素浓度为1.0-1.5ng/L,它在血浆中的半衰期仅为6-10mim。升压素的生理浓度很低,几乎没有收缩血管而致血压升高的作用,对正常血压调节没有重要性,但在失血情况下由于升压素释放较多,对维持血压有一定的作用。但是,升压素的抗利尿作用却十分明显,因此称为抗利尿激素较为适宜。
关于抗利尿激素的作用与分泌的调节,在第四章和第八章已有详细叙述。
(二)催产素
催产素具有促进乳汁排出一刺激子宫收缩的作用。
1.对乳腺的作用哺乳期乳腺不断分泌乳汁,贮存于腺泡中,当腺泡周围具有收缩性的肌上皮细胞时,腺泡压力增高,使乳汁从腺泡经输乳管由乳头射出。射乳是一典型的神经内分泌反射。乳头含有丰富的感觉神经末梢,吸吮乳头的感觉信息经传入神经传至下丘脑,使分泌催产素的神经元发生兴奋,神经冲动经下丘脑-垂体束传送到神经垂体,使贮存的催产素释放入血,并作用于乳腺中的肌上皮细胞使之产生收缩,引起乳汁排出,在射乳反射过程,血中抗利尿激素浓度毫无变化。在射乳反射的基础上,很容易建立条件反射,如母亲见到婴儿或听到其哭声均可引起条件反向性射乳。催产素除引起乳汁排出外,还有维持哺乳期乳腺不致萎缩的作用。
在射乳反射中,催乳素与催产素的分泌一同增加,而GnRH的释放减少。催乳素分泌增多促使GnRH分泌,对下一次射乳有利。GnRH释放减少引起腺垂体促性腺激素分泌减低,可导致哺乳期月经暂停。GnRH释放减少可能由于吸吮乳头刺激引起下丘脑多巴胺神经元兴奋,释放多巴胺,多巴胺可抑制GnRH的释放;也可能与下丘脑的β-内啡肽有关。它既可促进催乳素分泌,又可抑制GnRH的释放。
2.以子宫的作用催产素促进子宫肌收缩,但此种信息处理民子宫的功能状态有关。催产素对非孕子宫的作用较弱,而对妊娠子宫的作用较强,雌激素能增加子宫对催产素的敏感性,而孕激素则相反,催产素可使细胞外Ca2+进入子宫平滑肌细胞内,提高肌细胞内的Ca2+浓度,可能通过钙调蛋白的作用,并在蛋白激酶的参与下,诱发肌细胞收缩。研究表现,催产素虽然刺激子宫收缩,但它并不是发动分娩子宫进一步收缩。
由于催产素与抗利尿激素的化学结构相似,它们的生理作用有一定程度的交叉,例如,催产素对犬的抗利尿作用相当于抗利尿激素的1/200,而抗利尿激素对大鼠离体子宫的收缩作用为催产素的1/500左右。
第四节 甲状腺
甲状腺是人体内最大的内分泌腺,平均生理约为20-25g。甲状腺内含有许多大小不等的圆形或椭圆形腺泡。腺泡是由单层的上皮细胞围成,腺泡腔内充满胶质。胶质是腺泡上皮细胞的分泌物,主要成分为甲状腺球蛋白。腺泡上皮细胞是甲状腺激素的合成与释放的部位,而腺泡腔的胶质是激素有贮存库。腺泡上皮细胞的形态物质及胶质的量随甲状腺功能形态的不岢发生相应的变化。腺泡上皮细胞通常为立方形,当甲状腺受到刺激而功能活跃时,细胞变高呈低柱状,胶质减少;反之,细胞变低呈扁平形,而胶质增多。
在甲状腺腺泡之间和腺泡上皮细胞之间有滤泡旁细胞,又称C细胞,分泌降钙素。
一、甲状腺激素的合成与代谢
甲状腺激素主要有甲状腺素,又称甲碘甲腺原氨酸(thyroxine,3,5,3’,5’-tetraiodotyyronine,T4)和三碘甲腺原氨酸(3,5,3’-triiodothyronine,T3)两种,它们都是酷氨酸碘化物。另外,甲状腺也可合成极少量的逆-T3(3,3’,5’-T3或reverseT3,rT3),它不具有甲状腺激素有生物活性(图11-8)。
图11-8甲状腺激素有化学结构
甲状腺激素合成的原料有碘和甲状腺球蛋白,在甲状腺球蛋白的酪氨酸残基上发生碘化,并合成甲状腺激素。人每天从食物中大约摄碘100-200μɡ,占合身碘量的90%。因此,甲状腺与碘代谢的关系极为密切。
在胚胎期11-12周,胎儿甲状腺开始有合成甲状腺激素的能力,到13-14周在胎儿垂体促甲状腺激素的刺激下,甲状腺加强激素的分泌,这对胎儿脑的发育起着关键作用,因为母体的甲状腺激素进入胎儿体内的量很少。
甲状腺激素的合成过程包括三步:
(一)甲状腺腺泡聚碘
由肠吸收的碘,以I-形式存在于血液中,浓度为250μg/L,而μg/L内I-浓度比血液高20-25倍,加上甲状腺上皮细胞膜静息电位为-50mV,因此,I-从血液转运进入甲状腺上皮细胞内,必须逆着电化学梯度面进行主动转运,并消耗能量。在甲状腺腺泡上皮细胞在底面的膜上,可能存在I-转运蛋白,它依赖Na+-K+-ATP酶活动提供能量来完全I-的主动转运,因为用哇巴因抑制ATP酶,则聚碘作用立即发生障碍。有一些离子,如过氯酸盐的COO4-、硫氰桎卤的SCN-GnI-竞争转运机制,因此能抑制甲状腺的聚碘作用。摘除垂体可降低聚碘能力,而给予TSH则促进聚碘。用同位素(Na131I)示踪法观察甲状腺对放射性碘的摄取,在正常情况下有20%-30%的碘被甲状腺摄取,临床常用摄取放射性碘的能力来检查与判断甲状腺的功能状态。
(二)I-的活化
摄入腺泡上皮细胞的I-,在过氧化酶的作用下被活化,活化的部位在腺泡上皮细胞项端质膜微绒毛与腺泡腔交界处(图11-9)。活化过程的本质,尚未确定,可能是由I-变成I2或I。或是与过氧化酶形成某种复合物。
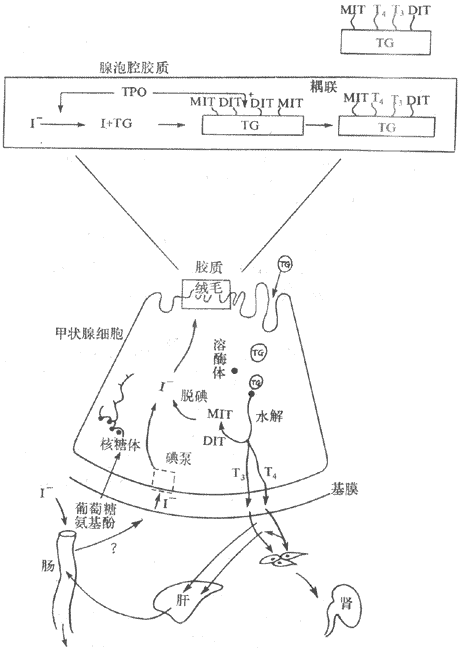
图11-9 甲状腺激素合成及代谢示意图
TPO:过氧化酶 TG:甲状球蛋白
I-的活化是碘得以取代酪氨酸残基上氢原子的先决条件。如先天缺乏过剩,I-不以活化,将使甲状腺激素有合成发生障碍。
(三)酷氨酸碘化与甲状腺激素的合成
在腺泡上皮细胞粗面内质网的核糖体上,可形成一种由四个肽链组成的大分子糖蛋白,即甲状腺球蛋白(thyroglobulin,TG),其分子量为670000,有3%的酪氨酸残基。碘化过程就是发生在甲状腺球蛋白的酪氨酸残基上,10%的酪氨酸残基可被碘化。放射自显影实验证明,注入放射性碘几分钟后,即可在甲状腺腺泡上皮细胞微绒毛与腺泡腔壁的上皮细胞残部,即能碘化甲状腺球蛋白,说明碘化过程发生在甲状腺腺泡上皮细胞微绒毛与腺泡交界处。
甲状腺球蛋白酪氨酸残基上的氢原子可被碘原子取代或碘化,首先生成一碘酪氨酸残基(MIT)和二碘酪氨残基(DIT),然后两个分子的DIT耦联生成四碘甲腺原氨酸(T4);一个分子的MIT与一个分子的DIT发生耦联,形成三碘甲腺原氨酸(T3),还能合成极少量的rT3(图11-9)
上述酪氨酸的碘化和碘化酪氨酸的耦联作用,都是在甲状腺球蛋白的分子上进行的,所在甲状腺球蛋白的分子上既含有酪氨酸、碘化酪氨酸,也常含有MIT、DIT和T4及T3。在一个甲状腺球蛋白分子上,T4与T3之比为20:1,这种比值常受碘含量的影响,当甲状腺内碘化活动增强时,DIT增多,T4含量也相应增加,在缺碘时,MIT增多,则T3含量明显增加。
甲状腺过氧化酶是由腺上皮细胞的核糖体生成的,它是一种含铁卟啉的蛋白质,分子量为60000-100000,在腺上皮顶缘的微绒毛处分布最多。实验证明,甲状腺过氧化酶的活性受TSH的调控,大鼠摘除垂体48h后,甲状腺过氧化酶活性消失,注入TSH后此酶活性再现。甲状腺过氧化酶的作用是促进碘活化、酪氨酸残基碘化及碘化酪氨酸的耦联等,所以,甲状腺过氧化酶晨甲状腺激素的合成过程中起关键作用,抑制此酶活性的药物,如硫尿嘧啶,便可抑制甲状腺激素的合成,可用于治疗甲状腺功能亢进。
(四)甲状腺激素有贮存、释放、运输与代谢
1.贮存 在甲状腺球蛋白上形成的甲状腺激素,在腺泡腔内以胶质的形式贮存。甲状腺激素有贮存有两个特点:一是贮存于细胞外(腺泡腔内);二是贮存的量很大,可供机体利用50-120天之久,在激素贮存的量上居首位,所以应用抗甲状腺药物时,用药时间需要较长才能奏效。
2.释放 当甲状腺受到TSH刺激后,腺泡细胞顶端即活跃起来,伸出伪足,将含有T4、T3及其他多种碘化酪酸残基的甲状腺球蛋白胶质小滴,通过吞饮作用,吞入腺细胞内(图11-9)。吞入的甲状腺球蛋白随即与溶酶体融合而形成吞噬体,并在溶酶体蛋白水解酶的作用下,将T4、T3以及MIT和DIT水解下来。甲状腺球蛋白分子较大,一般不易进入血液循环,而MIT和DIT的分子虽然较小,但很快受脱碘酶的作用而脱碘,脱下来的碘大部分贮存在甲状腺内,供重新利用合成激素,另一小部分从腺泡上皮细胞释出,进入血液。T4和T3对腺泡上皮细胞内的脱碘不敏感,可迅速进入血液。此外,尚有微量的rT3、MIT和DIT也可从甲状腺释放,进入血中。已经脱掉T4、T3、MIT和DIT的甲状腺球蛋白,则被溶酶体中的蛋白水解酶所水解。
由于甲状腺球蛋白分子上的T4数量远远超过T3,因此甲状腺分泌的激素主要是T4,约占总量的90%以上,T3的分泌量较少,但T3的生物活性比T4约大5倍
3.运输 T4与T3释放入血之后,以两种形式在血液中运输,一种是与血浆蛋白结合,另一种则呈游离状态,两者之间可互相转化,维持动态平衡。游离的甲状腺激素在血液中含量甚少,然而正是这些游离的激素才能进入细胞发挥作用,结合型的甲状腺激素是没有生物活性的。能与甲状腺激素结合的血浆蛋白有三种:甲状腺素结合球蛋白(thyroxine-binding globulin,TBG)、甲状腺素结合前白蛋白(thyroxine-bindingprealbumin,TBPA)与白蛋白。它们可与T4和T3发生不同程度的结合。血液中T4有99.8%是与蛋白质结合,其余10%与白蛋白结合。血中T4与TBG的结合受TBG含量与T4含量变化的影响,TBG在血浆听浓度为10mg/L,可以结合T4100-260μg。T3与各种蛋白的亲和力小得多,主要与TBG结合,但也只有T4结合量的3%。所以,T3主要以游离形式存在。正常成年人血清T4浓度为51-142nmol/L,T3浓度为1.2-3.4nmol/L。
4.代谢 血浆T4半衰期为7天,半衰期为1.5天,20%的T4与T3在肝内降解,也葡萄糖醛酸或硫酸结合后,经胆汁排入小肠,在小肠内重吸收极少,绝大部分被小肠液进一步分解,随粪排出。其余80%的T4在外周组织脱碘酶(5’-脱碘酶或5-脱碘酶)的作用下,产生T3(占45%)与rT3(占55%)。T4脱碘变成T3是T3的主要来源,血液中的T3有75%来自T4,其余来自甲状腺;rT3仅有少量由甲状腺分泌,绝大部分是在组织内由T4脱碘而来。由于T3的作用比T4大5倍,所以脱碘酶的活性将影响T4在组织内发挥作用,如T4浓度减少可使T4转化为T3增加,而使rT3减少。另外妊娠、饥饿、应激、代谢紊乱、肝疾病、肾功能衰竭等均会使T4转化为rT3增多。T3或rT3可再经脱碘变成二碘、一碘以及不含碘的甲状腺氨酸。另外,还有少量的T4与T3在肝和肾组织脱氨基和羧基,分别形成四碘甲状腺醋酸与在三碘甲状腺醋酸,并随尿排出体外。
二、甲状腺激素的生物学作用
T4与T3都具有生理作用。由于T4在外周组织中可转化为T3,而且T3的活性较大,曾使人认为T4可能是T3激素原,T4只有通过T3才有作用。目前认为,T4不仅可作为T3的激素原,而且其本身也具有激素作用,约占全部甲状腺激素作用的35%左右。临床观察发现,部分甲状腺功能低下患者的血中T3浓度强;另外,实验证明,在甲状腺激素作用的细胞核受体上,既存在T3结合位点,也有T4结合位点,T3或T4与其结合位点的亲和力是不同的,T3比T4高10倍。这些资料提示,T4本身也具有激素作用。
甲状腺激素的主要作用是促进物质与能量代谢,促进生长和发育过程。机体未完全分化与已分化的组织,对甲状腺激素的反应可以不同,而成年后,不同的组织对甲状腺的敏感性也有差别。甲状腺激素除了与核受体结合,影响转录过程外,在核糖体、线粒体、以及细胞膜上也发现了它的结合位点,可能对转录后的过程、线粒体的生物氧化作用以及膜的转运功能均有影响,所以,甲状腺激素的作用机制十分复杂。
(一)对代谢的影响
1.产热效应 甲状腺激素可提高绝大多数组织有耗氧率,增加产热量。有人估计,1mgT4可使组织产热增加,提高基础代谢率28%。给动物注射甲状腺激素后,需要经过一段较长时间的潜伏期才能出现生热作用。T4为24-48h,而T3为18-36h,T3的生热作用比T4强3-5倍,但持续时间较短。给动物注射T4或T3后,取出各种组织进入离体实验表明,心、肝、骨骼肌和肾等组织耗氧率明显增加,但另一些组织,如脑、肺、性腺、脾、淋巴结和皮肤等组织的耗氧率则不受影响。在胚胎期胎儿大脑组织可受甲状腺激素的作用而增加耗氧率,但出生后,大脑组织就失去了这种反应能力。
近年的研究表明,动物注射甲状腺激素后,心、肝、肾和骨骼肌等组织出现产热效应时,Na+-K+-ATP酶活性明显升高,如用哇巴因抑制此酶活性,则甲状腺激素的产热效应可完全被消除。又如,甲状腺功能低下的大鼠,血中甲状腺激素含量下降,其肾组织细胞膜Na+-K+-ATP酶活性减弱,若给予T4,酶的活性可恢复甚至增加,由此看来,甲状腺激素的产热作用与Na+-K+-ATP酶的关系十分密切。另外,有 人认为,甲状腺激素也能促进脂肪酸氧化,产生大量的热能。
甲状腺功能亢进时,产热量增加,基础代谢率升主患者喜凉怕热,极易出汗;而甲状腺功能低下时,产热量减少,基础代谢率降低,患者喜热恶寒,两种情况无法不能适应环境温度的变化。
2.对蛋白质、糖 和脂肪代谢的影响
(1)蛋白质代谢:T4或T3作用于核受体,刺激DNA转录过程,促进mRNA形成,加速蛋白质与各种酶的生成。肌肉、肝与肾的蛋白质合成明显增加,细胞数量增多,体积增大,尿氮减少,表现为正氮平衡。甲状腺激素分泌不足时,蛋白质合成减少,肌肉收缩无力,但组织间的粘蛋白增多,可结合大量的正离子和水分子,引起粘液性水肿(myxedema)。甲状腺分泌过多时,则加速蛋白质分解,特别是促进骨骼蛋白质分解,使肌酐含量降低,肌肉收缩元力,尿酸含量增加,并可促进骨的蛋白质分解,从而导致血钙升高和骨质疏松,尿钙的排出量增加。
(2)糖代谢:甲状腺激素促进小肠粘膜对糖的吸收,增强糖原分解,抑制糖原合成,并能增强肾上腺素、胰高血糖素、皮质醇和生长素的生糖作用,因此,甲状腺激素有升主血糖的趋势;但是,由于T4与T3还可加强外周组织对糖的利用,也有降低血糖的作用。甲状腺功能亢进时,血糖常升高,有时出现糖尿。
(3)脂肪代谢:甲状腺激素促进脂肪酸氧化,增强儿茶酚胺与胰高血糖素对脂肪的分解作用。T4与T3既促进胆固醇的合成,又可通过肝加速胆固醇的降解,而且分解的速度超过合成。所以,甲状腺功能亢进患者血中胆固醇含量低于正常。
甲状腺功能亢进时,由于蛋白质、糖和脂肪的分解代谢增强,所以患者常感饥饿,食欲旺盛,且有明显消瘦。
(二)对生成与发育的影响
甲状腺激素具有促进组织分化、生长与发育成熟的作用。切除甲状腺的蝌蚪,生长与发育停滞,不能变态成蛙,若及时给予甲状腺激素,又可恢复生长发育,包括长出肢体、尾巴消失,躯体长大,发育成蛙。在人类和哺乳动物,甲状腺激素是维持正常生长也发育不可缺少的激素,特别是对骨和脑的发育尢为重要。甲状腺功能低下的儿童,表现为以智力迟钝生身体矮小为特征的呆小症(又称克汀病)。在胚胎期缺碘造成甲状腺激素合成不足,或出生后甲状腺功能低下,脑的发育明显障碍,脑各部位的神经细胞变小,轴突、树突与髓鞘均减少,胶质细胞数量也减少。神经组织内的蛋白质、磷脂以及各种重要的酶与递质的含量都减低。甲状腺激素刺激骨化中心发育,软骨骨化,促进长骨和牙齿的生长。值得提出的是,在胚胎期胎儿骨的生长并不必需甲状腺激素,所以患先天性甲状腺发育不全的胎儿,出生后身长可以基本正常,但脑的发育已经受到程度不同的影响。在出生后数周至3-4个月后,就会表现出明显的智力迟钝和长骨生长停滞。所以,在缺碘地区预防呆小症的发生,应在妊娠期注意补充碘,治疗呆小症必须抓时机,应在生后三个月以前补给甲状腺激素,过迟难以奏效。
(三)对神经系统的影响
甲状腺激素不但影响中枢系统的发育,对已分化成熟的神经系统活动也有作用。甲状腺功能亢进时,中枢神经系统的兴奋性增高主要表现为注意力不易集中、过敏疑虑多愁善感、喜怒失常、烦躁不安、睡眠不好而且多梦幻,以及肌肉纤颤等。相反,甲状腺功能低下时,中枢神经系统兴奋性降低,出现记忆力减退,说话和行动迟缓,淡漠无怀与终日思睡状态。
甲状腺激素除了影响中枢神经系统活动外,也能兴奋交感神经系统,其作用机制还不十分清楚。
另外,甲状腺激素对心脏的活动有明显影响。T4与T3可使心率增快,心缩力增强,心输出量与心作功增加。甲状腺功能亢进患者心动过速,心肌可因过度耗竭而致心力衰竭。离体培养的心细胞实验表明,甲状腺激素可直接作用于心肌,T3能增加心肌细胞膜上β受体的数量,促进肾上腺素刺激心肌细胞内cAMP的生成。甲状腺激素促进心肌细胞肌质网释放Ca2+,从而激活与心肌收缩有关的蛋白质,增强收缩力。
三、甲状腺功能的调节
甲状腺功能活动主要受下丘脑与垂体的调节。下丘脑、垂体和甲状腺三个水平紧密联系,组成下丘脑-垂体-甲状腺轴。此外,甲状腺还可进行一定程度的自身调节。
(一)下丘脑-腺垂体对甲状腺的调节
腺垂体分泌的促甲状腺激素(thyroid stimulatinghormone,TSH)是调节甲状腺功能的主要激素。TSH是一种糖原白激素,分子量为28000,由α和β两个亚单位组成,α亚单位有96个氨基酸残基,其氨基酸顺序与LH、FSH和hCG的α亚单位相似;β亚单位有110个氨基酸残基,其顺序与以上三种激素有β亚单位完全不同。TSH的生物活性主要决定于β亚单位,但水解下来的单独β来只有微弱的活性,只有α亚单位与β亚单位结合在一起共同作用,才能显出全部活性。
血清中TSH浓度为2-11mU/L,半衰期约60min。腺垂体TSHA呈脉冲式释放,每2-4h出现一次波动,在脉冲式释放的基础上,还有日周期变化,血中TSH浓度清晨高而午后低。
TSH的作用是促进甲状腺激素有合成与释放。给予TSH最早出现的效果是甲状腺球蛋白水解与T4、T3的释放。给TSH数分钟内,甲状腺腺泡上皮细胞靠吞饮把胶质小滴吞入细胞内,加速T4与T3的释放,随后增强碘的摄取和甲状腺激素的合成。TSH还能促进腺泡上皮细胞的葡萄糖氧化,尤其经已糖化旁路,可提供过氧化酶作用所需要的还能型辅酶Ⅱ(NADPH)。TSH的长期效应是刺激甲状腺细胞增生,腺体增大,这是由于TSH刺激腺泡上皮细胞核酸与蛋白质合成增强的结果。切除垂体之后,血中TSH迅速消失,甲状腺发生萎缩,甲状腺激素分泌明显减少。
在甲状腺腺泡上皮细胞存在TSH受体,它是含有750个氨基酸残基的膜蛋白,分子量为85000。TSH与其受体结合后,通过G蛋白激活腺苷酸环化酶,使cAMP生成增多,进而促进甲状腺激素的释放与合成。TSH还可通过磷脂酰肌醇系统刺激甲状腺激素的释放与合成。
有些甲状腺功能亢进患者,血中可出现一些免疫球蛋白物质,其中之一是人类刺激甲状腺免疫球蛋白(human thyroid-stmulating immunoglobulin,HTSI),其化学结构与TSH相似,它可与TSH竞争甲状腺细胞腺上的受体刺激甲状腺,这可能是引起甲状腺功能亢进的原因之一。
腺垂体TSH分泌受下丘脑TRH的控制。下丘脑TRH神经元接受神经系统其他部位传来的信息影响,把环境因素与TRH神经元活动联系起来,然后TRH神经元释放TRH,作用于腺垂体。例如,寒冷刺激的信息到达中枢神经中枢神经系统,一方面传入下丘脑体温调节中枢,同时还与该中枢接近的TRH神经元发生联系,促进TRH释放增多,进而使腺垂体TRH分泌增加。在这一过程中,去甲上腺素趣了重要的递质作用,它能增强TRH神经元释放TRH,如阻断去甲肾上腺素的合成,则机体对寒冷刺激引起的这一适应性反应大大减弱。另外,下丘脑还可通过生长抑素减少或停止TRH的合成与释放。例如,应激刺激也可通过单胺能神经元影响生长抑素的释放,如外科手术与严重创伤将引起生长抑素的释放,从而使腺垂体分泌的TRH减少,T4与T3的分泌水平降低,减少机体的代谢消耗,有利于创伤修复过程。
(二)甲状腺激素的反馈调节
血中游离的T4与T3浓度的升降,对腺垂体TSH的分泌起着经常性反馈调节作用。当血中游离的T4与T3浓度增高时,抑制TSH分泌。实验表明,甲状腺激素抑制TSH分泌的作用,是由于甲状腺激素刺激腺垂体促甲状腺激素细胞产生一种抑制性蛋白,它使TSH的合成与释放减少,并降低腺垂体对TRH的反应性。由于这种抑制作用需要通过新的蛋白质合成,所以需要几小时后方能出现效果,而且可被放线菌D与放线菌酮所阻断。T4与T3比较,T3对腺垂体TSH分泌的抑制作用较强,血中T4与T3对腺垂体这种反馈作用与TRH的刺激作用,相互拮抗,相互影响,对腺垂体TSH的分泌起着决定性作用。
关于甲状腺激素对下丘脑是否有反馈调节作用,实验结果很不一致,尚难有定论。
另外,有引起激素也可影响腺垂体分泌TSH,如雌激素可增强腺垂体对TRH的反应,从而使TSH分泌增加,而生长素与糖皮质激素则对TSH的分泌有抑制作用。
图11-10 甲状腺激素分泌的调节示意图
⊕表示促进或刺激 (一)表示抑制
(三)甲状腺的自身调节
除了下丘脑-垂体对甲状腺进行调节以及甲状腺激素的反馈调节外,甲状腺本身还具有适应碘的供应变化,调节自身对碘的摄取以及合成与释放甲状腺激素的能力;在缺乏TSh 或TSH浓度不变的情况下,这种调节仍能发生,称为自身调节。它是一个有限度的缓慢的调节 系统。血碘浓度增加时,最初T4与T3的合成有所增加,但碘量超过一定限度后,T4与T3的合成在维持一高水平之后,旋即明显下降,当血碘浓度超过1mmol/L时,甲状腺摄碘能力开始下降,若血碘浓度达到10mmol/L时,甲状腺聚碘作用完全消失,即过量的碘可产生抗甲状腺效应,称为Wolff-Chaikoff效应。过量的碘抑制碘转运的机制,尚不十分清楚。如果在持续加大碘量的情况下,则抑制T4与T3合成的现象就会消失,激素的合成再次增加,出现对高碘含量的适应。相反,当血碘含量不足时,甲状腺将出现碘转运机制增强,并加强甲状腺激素的合成。
(四)自主神经对甲状腺活动的影响
荧光与电镜检查证明,交感神经直接支配甲状腺腺泡,电刺激一侧的交感神经,可使该侧甲状腺激素合成增加;相反,支配甲状腺的胆碱能纤维对甲状腺激素的分泌则是抑制性的。
第五节 甲状旁腺和甲状腺C细胞
甲状旁腺分泌的甲状腺激素(parathyroid hormone,PTH)与甲状腺C细胞分泌的降钙素(calcitonin,CT)以及1,25-二羟维生素D3共同调节钙磷代谢,控制血浆中钙和磷的水平。
一、甲状旁腺激素
PTH是甲状旁腺主细胞分泌的含有84个氨基酸的直链肽,分子量为9000,其生物活性决定于N端的第1-27个氨基酸残基。在甲状旁腺主细胞内先合成一个含有115个氨基酸的前甲状旁腺激素原(prepro-PTH),以后脱掉N端二十五肽,生成九十肽的甲状旁腺激素原(pro-PTH),再脱去6个氨基酸,变成PTH(图11-11)。

图11-11 人甲状旁腺激素的化学结构
在甲状旁腺主细胞内,部分PTH分子可以第33位与第40位氨基酸残基之间裂解,形成两个片断,可与PTH一同进入血中。正常人血浆PTh 浓度为10-50ng/L,半衰期为20-30min。PTH主要在肝水解灭活,代谢产物经肾排出体外。
近年从鳞状上皮癌伴发高血钙的患者癌组织中,分离出一种在化学结构上类似PTH的肽,称为甲状旁腺激素相关(parathyroid hormone related peptide,PTHrp),并进一步发现正常组织如皮肤、乳腺以及胎儿甲状旁腺中也存在这种肽。PTHrp与PTH从来源上是同族的,尤其两者的N端1-13位氨基酸残其完全相同,PTHrp也具有PTH活性。
(一)甲状旁腺激素的生物学作用
PTH是调节血钙水平的最重要激素,它有升高血钙和降低血磷含量的作用。将动物的甲状旁腺摘除后,血钙浓度逐渐降低,而血磷含量则逐渐升高,直至动物死亡。在人类,由于外科切除甲状腺时不慎,误将甲状旁腺摘除,可引起严重的低血钙。钙离子对维持神经和肌肉组织正常兴奋性起重要作用,血钙浓度降低时,神经和肌肉的兴奋性异常增高,可发生低血钙性手足搐搦,严重时可引起呼吸肌痉挛而造成窒息。
PTH对靶器官的作用是通过cAMP系统而实现的。
1.对骨的作用 骨是体内最大的钙贮存库,PTH动员骨钙入血,使血钙浓度升高,其作用包括快速效应与延缓效应两个时相。
(1)快速效应:在PTH作用后数分钟即可发生,是将位于骨和骨细胞之间的骨液中的钙转运至血液中,骨细胞和成骨细胞在骨内形成一个膜系统,全部覆盖了骨表面和腔隙的表面,在骨质与细胞外液之间形成一层可通透性屏障。在骨膜与骨质之间含有少量骨液,骨液中含有Ca2+(只有细胞外流人的1/3)。PTH能迅速提高骨细胞膜对Ca2+的通透性,使骨液中的钙进入细胞,进而使骨细胞膜上的钙泵活动增强,将Ca2+转运到细胞外液中(图11-12)。
图11-12 PTH对骨钙转运作用示意图
(2)延缓效应:在PTH作用后2-14h出现,通常在几天甚至几周后达高峰,这一效应是通过刺激破骨细胞活动增强而实验的。PTH既加强已有的破骨细胞的溶骨活动,又促进破骨细胞的生成。破骨细胞向周围骨组织伸出绒毛样突起,释放蛋白水解酶与乳酸,使骨组织溶解,钙与磷大量入轿,使血钙浓度长时间升主。PTH的两个效应相互配合,不但能对血钙急切需要作出迅速应答,而且能使血钙长时间维持在一定水平。
2.对肾的作用 PTH促进远球小管对钙的重吸收,使尿钙减少,血钙升高,同时还抑制近球不管对磷的重吸收,增加尿磷酸盐的排出,使血磷降低。
此外,PTH对肾的另一重要作用是激活α-羟化酶,使25-羟维生素D3(25-OH-D3)转变为有活性的1,25-二羟维生素D3(1,25-(OH)2-D3)。
3.1,25-(OH)2-D3的生成与作用体内的VD3(维生素D3)主要由皮肤中7-脱氢胆固醇经日光中些外线照射转化而来,也可由动物性食物中获取。VD3无生物活性,它首先需在肝羟化成25-OH-D3,然后在肾又进一步转化成1,25-(OH)2-D3,其作用为:①促进小肠粘膜上皮细胞对钙的吸收,这是由于1,25-(OH)2-D3进入小肠粘膜细胞内,与胞浆受体结合后进入细胞核,促进转录过程,生成一咱与钙有很高亲和力的钙结合蛋白(calcium-binding protein),参与钙的转运而促进钙的吸收。PTH在增强钙的吸收的同时也促进磷的吸收;②对骨钙动员和骨盐沉积有作用,一方面促进钙、磷的吸收,增加血钙、血磷含量,刺激成骨细胞的活动,从而促进骨盐沉积和骨的形成。另一方面,当血钙浓度降低时,又能提高破骨细胞的活性,动员骨钙入血,使血钙浓度升高。另外,1,25-(OH)2-D3能增强PTH对骨的作用,在缺乏1,25-(OH)2-D3时,PTH的作用明显减弱。
(二)甲状旁腺激素分泌的调节
PTH的分泌主要受血浆钙浓度变化的调节。血浆钙浓度轻微下降时,就可使甲状旁腺分泌PTH迅速增加,血钙浓度降低可直接刺激甲状旁腺细胞释放PTH,PTH动员骨钙入轿,增强肾重吸收钙,结果使已降低了血钙浓度迅速回升。相反,血钙浓度升高时,PTH分泌减少。长时间的高血钙,可使甲状旁腺发生萎缩,而长时间的低血钙,则可使甲状旁腺增生。
PTH的分泌还受其他一些因素的影响,如血磷升高可使血钙降低而刺激PTH的分泌。血Mg2+浓度很低时,可使PTH分泌减少。另外,生长抑素也能抑制PTH的分泌。
二、降钙素
降钙素是含有一个二硫键的三十二肽,分子量为3400。正常人血清中降钙素浓度为10-20ng/L,血浆半衰期小于1h,主要在肾降解并排出,降钙素整个分子皆为激素活性所必需,其化学结构如图11-13。
图11-13 人降钙素的化学结构
(一)降钙素的生物学作用
降钙素的主要作用是降低血钙和血磷,其主要靶器官是骨,对肾也有一定的作用。
1.对骨的作用 降钙素抑制破骨细胞活动,减弱溶骨过程,这一反应发生很快,大剂量的降钙素在15min内便可使破骨细胞活动减弱70%。在给降钙素1h左右,出现成骨细胞活动增强,持续几天之久。这样,降钙素减弱溶骨过程,增强成骨过程,使骨组织释放的钙磷减少,钙磷沉积增加,因而血钙与血磷含量下降。
成人降钙素对血钙的调节作用较小,因为降钙素引起的血钙浓度下降,可强烈地刺激PTH的。PTH的作用完全可以超过降钙素的效应。另外,成人的破骨细胞每天只能向细胞外液提供0.8g钙,因此,抑制破骨细胞的活动对血钙的影响是很小的。然而,儿童骨的更新速度很快,破骨细胞活动每天可向细胞外液提供5g以上的钙,相当于细胞外液总钙量的5-10倍,因此,降钙素对儿童血钙的调节则十分明显。
2.对肾的作用 降钙素能抑制肾小管对钙、磷、钠及氯的重吸收,使这些离子从尿中排出增多。
(二)降钙素分泌的调节
降钙素的分泌主要受血钙浓度的调节。当血钙浓度升高时,降钙素的分泌亦随之增加,降钙素与PTH对血钙的作用相反,区同调节血钙浓度的相对稳定。比较降钙素与PTH对血钙的调节作用,有两个主要的差别:①降钙素分泌启动较快,在1h内即可达到高峰,而PTH分泌则需几个小时;②降钙素只对血钙水平产生短期调节作用,其作用很快被有力的PTH作用所克服,后者对血钙浓度发挥长期调节作用,由于降钙素的作用快速而短暂,所以,对高钙饮食引起的血钙升高回复到正常水平起着重要作用。进食可刺激降钙素的分泌。这可能与几种胃肠激素如胃泌素、促胰液素以及胰高血糖素的分泌有关,它们都有促进降钙素分泌的作用,其中以胃泌素的作用最强。
现将PTH,1,25-(OH)2-D3和降钙素对血钙的调节作用及其相互关系总结于图11-14。
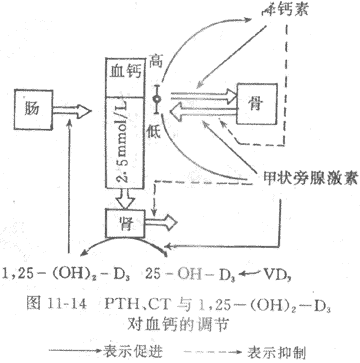
图11-14 PTH、CT与1,25-(OH)2-D3对血钙的调节
→表示促进→表示抑制
第六节 肾上腺
肾上腺包括中央部的髓质和周围部的皮质两个部分,两者在发生、结构与功能上均不相同,实际上是两种内分泌腺。
一、肾上腺皮质
肾上腺皮质分泌的皮质激素分为三类,即盐皮质激素、糖皮质激素和性激素。各类皮质激素是由肾上腺皮质不同层上皮细胞所分泌的,球状带细胞分泌盐皮质激素,主要是醛固醇(aldosterone);束状带细胞分泌糖皮质激素,主要是皮质醇(cortisol);网状带细胞主要分泌性激素,如脱氢雄酮(dehydroepiandrosterone)和雌二醇(estradiol)也能分泌少量的糖皮质激素。肾上腺皮质激素属于类固醇(甾体)激素,其基本结构为环戊烷多氢菲。盐皮质激素与糖皮质激素是21个碳原子的类固醇,雄激素含有19个碳原子,雌激素含有18个碳原子(图11-15)。
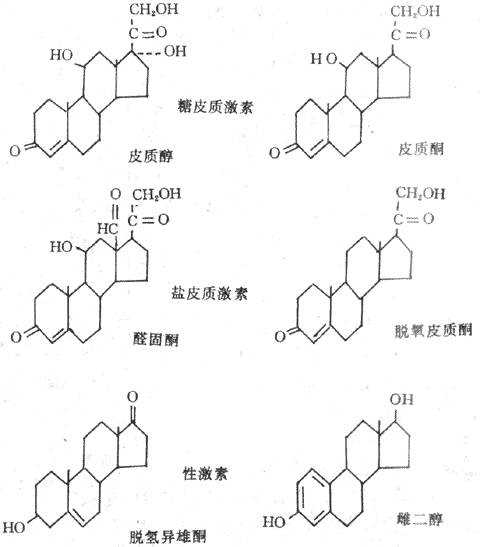
图11-15 几种主要的肾上腺皮质激素有化学结构
胆固醇是合成肾上腺皮质激素的原料,主要来自血液。在皮质细胞的线粒体内膜或内质网中所含的裂解酶与羟化酶等酶系的作用下,使胆固醇先变成孕烯酮,然后再进一步转变为各种皮质激素。由于肾上腺皮制裁各层细胞存在的酶系不同,所以合成皮质激素亦不相同(图11-16)。
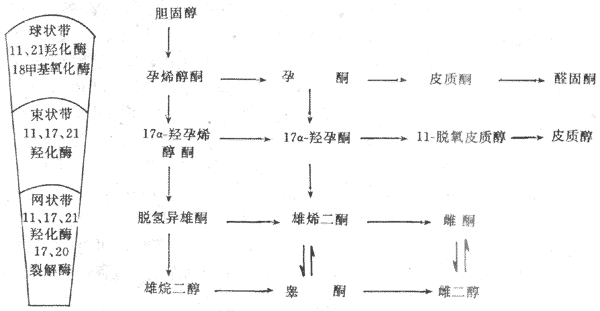
图11-16 肾上腺皮质激素合成的主要步骤
皮质醇进入血液后,75%-80%与血中皮质类固醇结合球蛋白(corticosteroid-binding globulin,CBG)或称为皮质激素运载蛋白结合,15%与血浆白蛋白结合,5%-10%的皮质醇是游离的。结合型与游离型皮质醇可以相互转化,维持动态平衡。游离的皮质醇能进入靶细胞发挥其作用。CBG是肝产生的α2球蛋白,分子量为52000,血浆中CBG浓度为30-50mg/L。CBG与皮质醇有较强有亲和力,每一分子的CBG仅有一个结合位点,只能结合一个分子的皮质醇。每100ml血浆CBG能结合20μg皮质醇。可见,CBG在运载皮质醇方面起着重要作用,。醛固醇与血浆白蛋白及CBG的结合能力很弱,主要以游离状态存在和运输。
皮质醇在血浆中半衰其为70min,醛固醇为20min。它们都在肝中降解,皮质醇首先是在C4GN C5间的双键加氢还原,形成双氢皮质醇,随后,C3上的酮基变成羟基产生四氢皮质醇,与葡萄糖醛酸或硫酸结合,随尿排出体外。四氢皮质醇是皮质醇的主要代谢产物,点尿中皮质醇代谢物排出量的45%-50%。四氢皮质醇和皮五醇在C20酮基变为痉基生成皮五醇,占尿中排出量的20%左右。由于四氢皮质醇和皮五醇在C17上均有羟基,故称为17-羟类固醇。另外,C1`7上脱去侧链,生成17-氧类固醇,占尿中排出题的10%左右。醛固醇基本上循类似途径被处理。
肾上腺皮质网状带分泌的性激素以脱氢异雄酮为主,它是一种17-氧类固醇,睾酮的代谢产物也是17-氧类固醇。因此,男子尿中17-氧类固醇的来源有睾丸分泌的睾酮和肾上腺皮质分泌的皮质醇及雄激素。
(二)肾上腺皮质激素的生物学作用
动物摘除双侧肾上腺后,如不适当处理,一二周即死去,如仅切除肾上腺髓质,动物可以存活较长时间,说明肾上腺皮质是维持生命所必需的。分析动物死亡的原因,主要有两个方面:其一是机体水盐损失严重,导致血压降低,终于因循环衰竭而死,这主要是缺乏盐皮质激素所致;其二是糖、蛋白质、脂肪等物质代谢发生严重紊乱,对各种有害刺激的抵抗力降低,导致功能活动失常,这是由于缺乏糖皮质激素的缘故。若及时补充肾上腺皮质激素,动物的生命可以维持。
1.糖皮质激素 人体血浆中糖皮质激素主要为皮质醇,其次为皮质酮,但皮质酮的含量仅为皮质醇的1/20-1/10。
(1)对物质代谢的影响:糖皮质激素对糖、蛋白质和脂肪代谢均有作用。①糖代谢;糖皮质激素是调节机体糖代谢的重要激素之一,它促进糖异生,升高血糖,这是由于它促进蛋白质分解,有较多的氨基酸进入肝,同时增强肝内与糖异生有关酶的活性,致使糖异生过程大大加强。此外,糖皮质激素又有抗胰岛素作用,促进血糖升主。如果糖皮质激素分泌过多(或服用此类激素药物过多)可引起血糖升高,甚至出现糖尿;相反,肾上腺皮质功能低下患者(如阿多数狄森病),则可出现低血糖;②蛋白质代谢:糖皮质激素促进肝外组织,特别是肌肉组织蛋白质分解,加速氨基酸转移至肝生成肝糖原。糖皮质激素分泌过多时,由于蛋白质分解增强,合成减少,将出现肌肉消瘦、骨质疏松、皮肤变薄、淋巴组织萎缩等;③脂肪代谢:糖皮质激素促进脂肪分解,增强脂肪酸在肝内氧化过程,有利于糖异生作用。肾上腺皮质功能亢进时,糖皮质激素对身体不同部位的脂肪作用不同,四肢脂肪组织分解增强,而腹、面、肩及背有脂肪合成有所增加,以致呈现面圆、背厚、躯干部发胖而四肢消瘦的特殊体形。
(2)对水盐代谢的影响:皮质醇有较弱的贮钠排钾作用,即对肾远由小管及集合管重吸收和排出钾有轻微的促进作用。此外,皮质醇还可以降低肾小球入球血管阻力,增加肾小球血浆流量而使肾小球滤过率增加,有利于水的排出。皮质醇对水负荷时水的快速排出有一定的作用,肾上腺皮质功能不足患者,排水能力明显降低,严重时可出现“水中毒”,如补充适量的糖皮质激素即可得到缓解,而补充盐皮质激素则无效。有资料指出,在缺乏皮质醇时,ADH释放增多,集合管对水的重吸收增加。
(3)对血细胞的影响:糖皮质激素可使血中红细胞、血小板和中性粒细胞的数量增加,而使淋巴细胞和嗜酸性粒细胞减少,其原因各有不同。红细胞和血小板的增加,是由于骨髓造血功能增强;中性粒细胞的增加,可能是由于附着在小血管壁边缘的中性粒细胞进入血液循环增多所致;至于淋巴细胞减少,可能是糖皮质激素使淋巴细胞DNA合成过程减弱,抑制胸腺与淋巴组织的细胞分裂。此外,糖皮质激素还能促进淋巴细胞与渚酸性粒细胞破坏。
(4)对循环系统的影响:糖皮质激素对维持正常血压是必需的,这是由于:①糖皮质激素能增强血管平滑肌对儿茶酚胺的敏感性(允许作用),这可能由于糖皮质激素能啬血管平滑肌细胞膜上的儿茶酚胺受体数量以及调节受体介导的细胞内的信息传递过程;②糖皮质激素能抑制具有血管舒张作用的前列腺素的合成;③糖皮质激素能降低毛细血管的通透性,有利于维持血容量。肾上腺皮质功能低下时,血管平滑肌对儿茶酚胺的反应性降低,毛细血管扩张,通透性增加,血压下降,补充皮质醇后可恢复。
另外,离体实验证明,糖皮质激素可增强心肌的收缩力,但在整体条件下对心脏的作用并不明显。
(5)在应激反应中的作用:当机体受到各种有害刺激,如缺氧、创伤、手术、饥饿、疼痛、寒冷以及精神紧张和焦虑不安等。血中ACTH浓度立即增加,糖皮质激素也相应增多。能引起ACTH与糖皮质激素分泌增加的各种刺激称为应激刺激,而产生的反应称为应激(stress)。在这一反应中,除垂体-肾上腺皮质系统参加外,交感-肾上腺髓质系统也参加,所以,在应激反应中,血中儿茶酚胺含量也相应增加。切珍重肾上腺髓质的动物,可以抵抗应激而不产生严重后果,而当去掉肾上腺皮质时,则机体应激反应减弱,对有害刺激的抵抗力大大降低,严重时可危及生命。应激反应可能从以下几个方面调节机体的适应能力:
①减少应激刺激引起的一些物质(缓激肽、蛋白水解酶及前列腺素等)的产生量及其不良作用;②使能量代放运转以糖代谢为中心,保持葡萄糖对重要器官(如脑和心)的供应;③在维持血压方面起允许作用,增强儿茶酚胺对血管的调节作用。应该指出,在应激反应中,除了ACTH、糖皮质激素与儿茶酚胺的分泌增加外,β-内啡肽、生长素、催乳素、抗利尿激素、胰主血压素及醛固醇等均可增加,说明应激反应是多种激素参与并使机体抵抗力增强的非特异性反应。
糖皮质激素的作用广泛而复杂,以上仅简述了它们的主要作用。此外,还有多方面的作用,如促进胎儿肺表面活性物质的合成,增强骨骼肌的收缩力,提高胃腺细胞对迷走神经与胃泌素的反应性,增加胃酸与胃蛋白酶原的分泌,抑制骨的形成而促进其分解等。临床上使用大剂量的糖皮质激素及其类似物,可用于抗炎、抗过敏、抗毒和抗休克。
2.盐皮质激素主要为醛固醇,对水盐代谢的作用最强,其次为脱氧皮质醇(表11-3)。
表11-3 几种肾上腺皮质激素对糖代谢作用的比较
| 激素 | 对糖代谢作用 | 保钠排钾作用 |
| 皮质醇 | 1.0 | 1.0 |
| 可地松 | 0.8 | 0.8 |
| 皮质酮 | 0.5 | 1.5 |
| 醛固酮 | 0.25 | 500 |
| 脱氧皮质酮 | 0.01 | 30 |
表中数字代表皮质激素的相对效力,以皮质醇的效力为1.0,即醛固酮的保钠排钾作用为皮质醇的500倍
醛固酮是调节机体水盐代谢的重要激素,它促进肾远曲小管及集合管重吸收钠、水和排出钾,即保钠、保水和排钾作用。当醛固酮分泌过多时,将使钠和水贮留,引起高血钠、高血压和血钾降低。相反,醛固酮缺乏时则钠与水的排出过多,血钠减少,血压降低,而尿钾排出减少,血钾升高。关于醛固酮对肾的作用及其机制,可参阅第八章,另外,盐皮质激素与糖皮质激素一样,以增强血管平滑肌对儿茶酚胺的敏感性,且作用比糖皮质激素更强。
(三)肾上原皮质激素分泌的调节
1.糖皮质激素分泌的调节肾上腺皮质分泌皮质激素的束状带及网状带,处于腺垂体保肾上腺皮质激素(adrenocortiotropin,ACTH)的经常性控制之下,无论是糖皮质激素的基础分泌,还是在应激状态下的分泌,都受ACTH的调控,切除动物的垂体后,束状带与网状带萎缩,糖皮质激素有分泌显着减少,如及时补充ACTH,可使已发生萎缩的束状带与网状带基本恢复,糖皮质激素有分泌回升。
(1)ACTH:ACTH是一个含39个氨基酸的多肽,分子量为4500,其化学结构示于图11-17。
图11-17 人ACTH的化学结构
ACTH分子上的1-24位氨基酸为生物活性所必需的,25-39位氨基酸可保护激素,减慢降解,延长作用时间。各种动物的ACTH前24位氨基酸均相同,因此,从动物(牛、羊、猪等)腺垂体提到的ACTH对人有效。目前,ACTH已能人工合成。在垂体,ACTH是由阿黑皮素原(POMC)经酶分散而来,同时产生β-MSH。ACTH再经酶分解生成α-MSH,ACTH的第4-10位氨基酸与α-MSH第4-10位氨基酸和β-MSH第11-17位氨基酸相同,这部分氨基酸是产生MSH活性最小单位,因此ACTH也具有促黑素细胞产生黑色素的作用。
ACTH的分泌呈现日节律波动,入睡后ACTH分泌逐渐减少,午夜最低,随后又逐渐增多,至觉醒起床前进入分泌高峰,白天维持在较低水平,入睡时再减少。由于ACTH分泌的日节律波动,促糖皮质激素的分泌也出现相应的波动。ACTH分泌的这种日节律波动,是由下丘脑CRH节律性释放所决定的。
ACTH 不但刺激糖皮质激素的分泌,也刺激束状带与网状带细胞的生长发育,关于ACTH的作用机制已基本清楚。在束状还与网状带细胞膜上存在ACTH特异性受体,在Ca2+存在的条件下,ACTH与膜受体结合,激活腺苷酸环化酶,通过cAMP激活蛋白激酶,蛋白激酶起三项重要作用;①使核糖蛋白磷酸化,促进mRNA形成一种特殊蛋白质,使胆固醇得以进入线粒体,并经侧链解形成孕烯醇酮,以进一步合成糖皮质激素;②使磷酸化酶活化,促进糖原分解,产生ATP,提供能量,另外还通过戊糖旁路产生还原型辅酶Ⅱ(NADPH),以利胆固醇的羟化过程;③使胆固醇酯活化,促进其转变为胆固醇,提供激素合成的原料。在ACTH促进肾上腺皮质细胞合成糖皮质激素的同时,束状带细胞膜对葡萄糖与胆固醇的转运机制增强,使较多的葡萄糖与胆固醇进入细胞内(图11-18)。
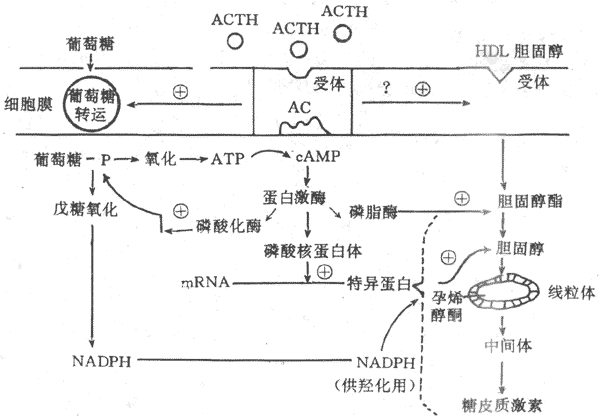
图11-18 ACTH作用机制示意图
HDL:高密度脂蛋白 AC:腺苷酸环化酶
(2)ACTH分泌的调节:ACTH调节糖皮质激素的分泌,而ACTH的分泌受下丘脑CRH的控制又与糖皮质激素有反馈调节。下丘脑CRH神经元和其他下丘脑调节肽神经元一样,又受脑内神经递质的调控。应激刺激作用于神经系统的不同部位,最后通过神经递质,将信息汇集于CRH神经元,然后借CRH控制腺垂体的促肾上腺皮质激素细胞分泌ACTH。此外,当血中糖皮质激素浓度升高时,可使腺垂体释放ACTH减少,ACTH的合成也受到抑制,腺垂体对CRH的反应也性减弱。糖皮质激素的负反馈调节主要作用于垂体,也可作用于下丘脑,这后一种反馈称为长反馈。ACTH还可反馈抑制CRH神经元,称为短反馈。至于是否存在CRH对CRH神经地的超短反馈,尚不能肯定。
综上所述,下丘脑、垂体和肾上腺皮质组成一个密切联系、协调统一的功能活动轴,从而维持血中糖皮质激素浓度的相对稳定和在不同状态下的适应性变化(图11-19)。
2.盐皮质激素分泌的调节 醛固酮的分泌主要受肾素-血管紧张素系统的调节。另外,血K+,血Na+浓度可以直接作用于球状带,影响醛固酮的分泌(详见第四章与第八章)。
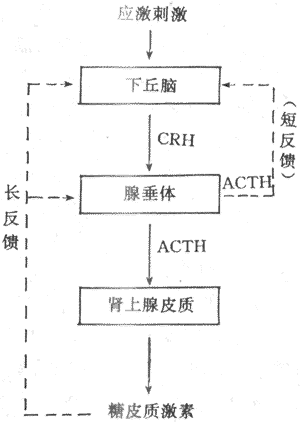
图11-19 糖皮质激素分泌的调节示意图
实线表示促进 点线表示抑制
在正常情况下,ACTH对醛固酮的分泌并无调节作用,但切除垂体后,在应激醛固酮的分泌反应减弱,提示在应激情况下,ACTH对醛固酮的分泌可能起到一定的支持作用。
二、肾上腺髓质
肾上腺髓质嗜铬细胞分泌肾上腺素(epinephrine,E)和去甲肾上腺素(norepinephrine,NE)都是儿茶酚胺激素。
(一)髓质激素的合成与代谢
髓质激素的合成与交感神经节后纤维合成去甲肾上腺素的过程基本一致,不同的是在嗜铬细胞胞浆中存在大量的苯乙醇胺氮位甲基移位酶(phenylethanolamine-N-methyltransferase,PNMT),可使去甲肾上腺素甲基化而成肾上腺素。合成髓质激素有原料分为酪氨酸,其合成过程为:酪氨酸→多巴→多巴胺→去甲肾上腺素→肾上腺素,各个步骤分别在特异酶,如酷氨酸羟化酶、多巴脱羟酶、多巴胺β-羟化酶及PNMT的作用下,最后生成肾上腺素。
肾上腺素与去甲肾上腺素一起贮存在髓质细胞的囊泡里内,以待释放。髓质中肾上腺素与去甲肾上腺素的比例大约为4:1,以肾上腺素为主。在血液中去甲肾上腺素除由髓质分泌外,主要来自肾上腺素能神经纤维末梢,而血中肾上腺素主要来自肾上腺髓质。
在体内的肾上腺素与去甲肾上腺素通过单胺氧化酶(monoamineoxidase,MAO)与儿茶酚-O-甲基移位酶(catechol-O-methyltransferase,COMT)的作用而灭活。
(二)髓质激素的生物学作用
髓质与交感神经系统组成交感-肾上腺髓质系统,或称交感-肾上腺系统,所以,髓质激素的作用与交感神经紧密联系,难以分开。生理学家Cannon最早全面研究了交感-肾上腺髓质系统的作用,曾提出应急学说(emergency reactionhypothesis),认为机体遭遇特殊情况时,包括畏惧、剧痛、失血、脱水、乏氧、暴冷暴热以及剧烈运动等,这一系统将立即调动起来,儿茶酚胺(去肾上腺素、肾上腺素)的分泌量大大增加。儿茶酚胺作用于中枢神经系统,提高其兴奋性,使机体处于警觉状态,反应灵敏;呼吸加强加快,肺通气量增加;心跳加快,心缩力增强,心输出量增加。血压升高,血液循环加快,内脏血管收缩,骨骼肌血管舒张同时血流量增多,全身血液重新分配,以利于应急时重要器官得到更多的血液供应;肝糖原分解增加,血糖升高,脂肪分解加强,血中游离脂肪酸增多,葡萄糖与脂肪酸氧化过程增强,以适应在应急情况下对能量的需要。总之,上述一切变化都是在紧急情况下,通过交感-肾上腺髓质系统发生的适应性反应,称之为应急反应。实际上,引起应急反应的各种刺激,也是引起应激反应的刺激,当机体受到应激刺激时,同时引起应急反应与应激反应,两者相辅相成,共同维持机体的适应能力。
(三)髓质激素分泌的调节
1.交感神经 髓质受交感神经胆碱能节前纤维支配,交感神经兴奋时,节前纤维末梢释放乙酰胆碱,作用于髓质嗜铬细胞上的N型受体,引起肾上腺素与去甲肾上腺素的释放。若交感神经兴奋时间较长,则合成儿茶酚胺所需要的酪氨酸羟化酶、多巴胺β-羟化酶以及PNMT的活性均增强,从而促进儿茶酚胺的合成。
2.ACTH与糖皮质激素动物摘除垂体后,髓质中酪氨酸氢化酶、多巴胺β-羟化酶与PNMT的活性降低,而补充ACTH则能使这种酶的活性恢复,如给予糖皮质激素可使多巴胺β-羟化酶与PNMT活性恢复,而对酪酸羟化酶未见明显影响,提示ACTH促进髓质合成儿茶酚胺的作用,主要通过糖皮质激素,也可能有直接作用。肾上腺皮质的血液经髓质后才流回循环,这一解剖特点有利于糖皮质激素直接进入髓质,调节儿茶酚胺的合成。
3.自身反馈调节去甲肾上腺素或多巴胺在髓质细胞内的量增加到一定数量时,可抑制酪氨酸羟化酶。同样,肾上腺素合成增多时,也能抑制PNMT的作用,当肾上腺素与去甲肾上腺素从细胞内释入血液后,胞浆内含量减少,解除了上述的负反馈抑制,儿茶酚胺的合成随即增加(图11-20)。
图11-20 肾上腺髓质激素生物合成示意图
PNMT:苯乙醇胺氮位甲基移位酶
第七节 胰岛
人类的胰岛细胞按其染色和形态学特点,主要分为A细胞、B细胞、D细胞及PP细胞。A细胞约占胰胰岛细胞的20%,分泌胰主血糖素(glucagon);B细胞占胰岛细胞的60%-70%,分泌胰岛素(insulin);D细胞占胰岛细胞的10%,分泌生成抑素;PP细胞数量很少,分泌胰多肽(pancreatic polyeptide)。
一、胰岛素
胰岛素是含有51个氨基酸的小分子蛋白质,分子量为6000,胰岛素分子有靠两个二硫键结合的A链(21个氨基酸)与B链(30个氨基酸),如果二硫键被打开则失去活性(图11-21)。B细胞先合成一个大分子的前胰岛素原,以后加工成八十六肽的胰岛素原,再经水解成为胰岛素与连接肽(C肽)。
图11-21 人胰岛素的化学结构
胰岛素与C肽共同释入血中,也有少量的胰岛素原进入血液,但其生物活性只有胰岛素的3%-5%,而C肽无胰岛素活性。由于C肽是在胰岛素合成过程产生的,其数量与胰岛素的分泌量有平行关系,因此测定血中C肽含量可反映B细胞的分泌功能。正常人空腹状态下血清胰岛素浓度为35-145pmol/L。胰岛素在血中的半衰期只有5min,主要在肝灭活,肌肉与肾等组织也能使胰岛素失活。
1965年,我国生化学家首先人工合成了具有高度生物活性的胰岛素,成为人类历史上第一次人工合成生命物质(蛋白质)的创举。
(一)胰岛素的生物学作用
胰岛素是促进合成代谢、调节血糖稳定的主要激素。
1.对糖代谢的调节 胰岛素促进组织、细胞对葡萄糖的摄取和利用,加速葡萄糖合成为糖原,贮存于肝和肌肉中,并抑制糖异生,促进葡萄糖转变为脂肪酸,贮存于脂肪组织,导致血糖水平下降。
胰岛素缺乏时,血糖浓度升高,如超过肾糖阈,尿中将出现糖,引起糖尿病。
2.对脂肪代谢的调节 胰岛素促进肝合成脂肪酸,然后转运到脂肪细胞贮存。在胰岛素的作用下,脂肪细胞也能合成少量的脂肪酸。胰岛素还促进葡萄糖进入脂肪细胞,除了用于合成脂肪酸外,还可转化为α-磷酸甘油,脂肪酸与α-磷酸甘油形成甘油三酯,贮存于脂肪细胞中,同时,胰岛素还抑制脂肪酶的活性,减少脂肪的分解。
胰岛素缺乏时,出现脂肪代谢紊乱,脂肪分解增强,血脂升高,加速脂肪酸在肝内氧化,生成大量酮体,由于糖氧化过程发和障碍,不能很好处理酮体,以致引起酮血症与酸中毒。
3.对蛋白质代谢的调节 胰岛素促进蛋白质合成过程,其作用可在蛋白质合成的各个环节上:①促进氨基酸通过膜的转运进入细胞;②可使细胞核的复制和转录过程加快,增加DNA和RNA的生成;③作用于核糖体,加速翻译过程,促进蛋白质合成;另外,胰岛素还可抑制蛋白质分解和肝糖异生。
由于胰岛素能增强蛋白质的合成过程,所以,它对机体的生长也有促进作用,但胰岛素单独作用时,对生长的促进作用并不很强,只有与生长素共同作用时,才能发挥明显的效应。
近年的研究表明,几乎体内所有细胞的膜上都有胰岛素受体。胰岛素受体已纯化成功,并阐明了其化学结构。胰岛素受体是由两个α亚单位和两个β亚单位构成的四聚体,α亚单位由719个氨基酸组成,完全裸露在细胞膜外,是受体结合胰岛素的主要部位。α与α亚单位、α与β亚单位之间靠二硫键结合。β亚单位由620个氨基酸残基组成,分为三个结构域:N端194个氨基酸残基伸出膜外;中间是含有23个氨基酸残基的跨膜结构域;C端伸向膜内侧为蛋白激酶结构域。胰岛素受体本身具有酪氨酸蛋白激酶活性,胰岛素与受体结合可激活该酶,使受体内的酪氨酸残基发生磷酸化,这对跨膜信息传递、调节细胞的功能起着十分重要的作用。关于胰岛素与受体结合启动的一系列反应,相当复杂,尚不十分清楚。
(二)胰岛素分泌的调节
1.血糖的作用 血糖浓度是调节胰岛素分泌的最重要因素,当血糖浓度升高时,胰岛素分泌明显增加,从而促进血糖降低。当血糖浓度下降至正常水平时,胰岛素分泌也迅速恢复到基础水平。在持续高血糖的刺激下,胰岛素的分泌可分为三个阶段:血糖升高5min内,胰岛素的分泌可增加约10倍,主要来源于B细胞贮存的激素释放,因此持续时间不长,5-10min后胰岛素的分泌便下降50%;血糖升高15min后,出现胰岛素分泌的第二次增多,在2-3h达高峰,并持续较长的时间,分泌速率也远大于第一相,这主要是激活了B细胞胰岛素合成酶系,促进了合成与释放;倘若高血糖持续一周左右,胰岛素的分泌可进一步增加,这是由于长时间的高血糖刺激B细胞增生布引起的。
2.氨基酸和脂肪酸的作用 许多氨基酸都有刺激胰岛素分泌的作用,其中以精氨酸和赖氨酸的作用最强。在血糖浓度正常时,血中氨基酸含量增加,只能对胰岛素的分泌有轻微的刺激作用,但如果在血糖升高的情况下,过量的氨基酸则可使血糖引起的胰岛素分泌加倍增多。务右脂肪酸和酮体大量增加时,也可促进胰岛素分泌。
3.激素的作用 影响胰岛素分泌的激素主要有:①胃肠激素,如胃泌素、促胰液素、胆囊收缩素和抑胃肽都有促胰岛素分泌的作用,但前三者是在药理剂量时才有促胰岛素分泌作用,不像是一引起生理刺激物,只有抑胃肽(GIP)或称依赖葡萄糖的促胰岛素多肽(glucose-dependentinsulin-stimulating polypeptide)才可能对胰岛素的分泌起调节作用。GIP是由十二指肠和空肠粘膜分泌的,由43个氨基酸组成的直链多肽。实验证明,GIP刺激胰岛素分泌的作用具有依赖葡萄糖的特性。口服葡萄糖引起的高血糖和GIP的分泌是平行的,这种平行关系的绘双导致胰岛素的迅速而明显的分泌,超过了静脉注射葡萄糖所引起的胰岛素分泌反应,。有人给大鼠口吸取葡萄糖并注射GIP抗血清,结果使血中葡萄浓度升高,而胰岛素水平却没有明显升高,因此可以认为,在肠内吸收葡萄糖期间,GIP是小肠粘膜分泌的一种主要的肠促胰岛素因子。除了葡萄糖外,小肠吸收氨基酸、脂肪酸及盐酸等也能刺激GIP的释放。有人将胃肠激素与胰岛素分泌之间的关系称为“肠-胰岛轴”,这一调节作用具有重要的生理意义,使食物尚在肠道中时,胰岛素的分泌便已增多,为即将从小肠吸收的糖、氨基酸和脂肪酸的利用做好准备;②生长素、皮质醇、甲状腺激素以及胰高血糖素告示可通过升高血糖浓度而间接刺激胰岛素分泌,因此长期大剂量应用这些激素,有可能使B细胞衰竭而导致糖尿病;③胰岛D细胞分泌的生长抑至少可通过旁分泌作用,抑制胰岛素和胰高血糖的分泌,而胰高血糖素也可直接刺激B细胞分泌胰岛素(图11-22)。
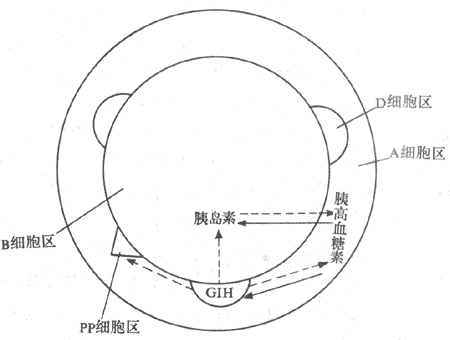
图11-22 胰岛细胞的分布及其分泌激素之间的相互影响
→表示促进 ----→表示抑制 GIH:生长抑素
4.神经调节 胰岛受迷走神经与交感神经支配。刺激迷起神经,可通过乙酰胆碱作用于M受体,直接促进胰岛素的分泌;迷走神经还可通过刺激胃肠激素的释放,间接促进胰岛素的分泌。交感神经兴奋时,则通过去甲肾上腺素作用于α2受体,抑制胰岛素的分泌。
二、胰高血糖素
人胰高血糖是由29个氨基酸组成的直链多肽,分子量为3485,它也是由一个大分子的前体裂解而来。胰高血糖在血清中的浓度为50-100ng/L,在血浆中的半衰期为5-10min,主要在肝灭活,肾也有降解作用。
(一)胰高血糖的主要作用
与胰岛素的作用相反,胰高血糖素是一种促进分解代谢的激素。胰高血糖素具有很强的促进糖原分解和糖异生作用,使血糖明显升高,1mol/L的激素可使3×106mol/L的葡萄糖迅速从糖原分解出来。胰高血糖素通过cAMP-PK系统,激活肝细胞的磷酸化酶,加速糖原分解。糖异生增强是因为激素加速氨基酸进入肝细胞,并激活糖异生过程有关的酶系。胰高血糖素还可激活脂肪酶,促进脂肪分解,同时又能加强脂肪酸氧化,使酮体生成增多。胰高血糖素产生上述代谢效应的靶器官是肝,切除肝或阻断肝血流,这些作用便消失。
另外,胰高血糖素可促进胰岛素和胰岛生长抑素的分泌。药理剂量的胰高血糖素可使心肌细胞内cAMp 含量增加,心肌收缩增强。
(二)胰高血糖素分泌的调节
影响胰高血糖素分泌的因素很多,血糖浓度是重要的因素。血糖降低时,胰高血糖素胰分泌增加;血糖升高时,则胰高血糖素分泌减少。氨基酸的作用与葡萄糖相反,能促进胰高血糖素的分泌。蛋白餐或静脉注入各种氨基酸均可使胰高血糖素分泌增多。血中氨基酸增多一方面促进胰岛素释放,可使血糖降低,另一方面还能同时刺激胰高血糖素分泌,这对防止低血糖有一定的生理意义。
胰岛素可通过降低血糖间接刺激胰高血糖素的分泌,但B细胞分泌的胰岛不比和D细胞分泌的生长抑素可直接作用于邻近的A细胞,抑制胰高血糖素的分泌(图11-22)。
胰岛素与胰高血糖素是一对作用相反的激素,它们都与血糖水平之间构成负反馈调节环路。因此,当机体外于不同的功能状态时,血中胰岛素与胰高血糖素的摩尔比值(I/G)也是不同的。一般在隔夜空腹条件下,I/G比值为2.3,但当饥饿或长时间运动时,比例可降至0.5以下。比例变小是由于胰岛素分泌减少与胰高血糖素分泌增多所致,这有利于糖原分解和糖异生,维持血糖水平,适应心、脑对葡萄糖的需要,并有利于脂肪分解,增强脂肪酸氧化供能。相反,在摄食或糖负荷后,比值可升至10以上,这是由于胰岛素分泌增加而胰高血糖素分泌减少所致。在这种情况下,胰岛不比的作用占优势。
第八节 松果体其他
一、松果体
松果体细胞是由神经细胞演变而来的,它分泌的激素主要有褪黑素和肽类激素。来自颈上交感神经节后神经末梢与松果体细胞形成突触联系,通过释放去甲上腺素控制松果体细胞的活动。
(一)褪黑素
1959年Lerner从牛松果体提取物中分离出一种能使青蛙皮肤褪色的物质,并命名为褪色素(melatonin),其化学结构为5-甲氧基-N-乙酰色胺。在松果体内羟化酶、脱羟酶、乙酰移位酶及甲基移位酶的作用下,色氨酸转变为褪色素。
松果体褪色素的分泌出现在明显的昼夜节律变化,白天分泌减少,而黑夜分泌增加。实验证明,大鼠在持续光照下,松果体重量变轻,细胞变小,合成褪色素的酶系活性明显降低,因而褪色素合成减少。反之,致盲大鼠或大鼠持续在黑暗环境中,将使松果体合成褪色素的酶系活发生增强,褪色素的合成随之增加。摘除动物的眼球或切断支配松果体的交感神经,则褪色素分泌的昼夜节律不再出现,说明光-暗对松果体活动的影响与视觉和交感神经有关。刺激交感神经可使松果体活动增强,而β-肾上腺素能受体阻断剂可阻断交感神经对松果体的刺激作用。如毁损视交叉上核,褪色素的昼夜节律性分泌消失。所以视交叉上核被认为是控制褪色素分泌的昼夜节律中枢,在黑暗条件下,视交叉上核即发出冲动传到颈上交感神经节,其节后纤维末梢释放去甲肾上腺素,与松果体细胞膜上的β-肾上腺素能受体结合,激活腺苷酸环化酶,通过cAMP-PK系统,增强褪色素合成酶系的活性,从而导致褪色素合成增加,在光刺激下,视网膜的传入冲动可抑制交感神经的活动,使褪色素合成减少。
褪色素对下丘脑-垂体-性腺轴与下丘脑-垂体-甲状腺活动均有抑制作用。切除幼年动物的松果体,出现性早熟,性腺与甲状腺的重量增加,功能活动增强。远在一个世纪之前,人们就发出某些性早熟男孩是因松果体肿瘤所致,因此认为松果体在青春期有抗性腺功能作用。正常妇女血中褪色素在有经周期的排卵前夕最低,随后在黄体期逐渐升高,月经来潮时达到顶峰,提示妇女朋经周期的节律与松果体的节律关系密切。
(二)肽类激素
松果体能合成GnRH、TRH及8精-(氨酸)催产素等肽类激素。在多种哺乳动物(鼠、牛、羊、猪等)的松果体内GnRH比同种动物下丘脑所含的GnRH量高4-10倍。有人认为,松果体是GnRH和TRH的补充来源。
二、胸腺
胸腺能分泌多种肽类物质,如胸腺素(thymosin)、胸腺生长素(thymopoietin)等,它们促进T细胞分化成熟。
三、前列腺素
前列腺素(prostaglandin,PG)是广泛存在于动物和人体内的一组重要的组织激素。PG的化学结构一般是具有五元环和两条侧链的二十碳不饱和脂肪酸。根据其分子结构的不同,可把PG分为A、B、D、E、F、H、I等型。
细胞膜的磷脂化在磷脂酶A2的作用下,生成PG的前体枣花生四烯酸。花生四烯酸在环氧化酶的催化下,形成不稳定的环内过氧化物枣PGG2,随后又转变为PGH2。PGH2在异构酶或还原酶的作用下,分别形成PGE2或PGF2α。PGG2与PGH2又可前列素合成酶的作用下,转变为前列环素(PGI2),在血栓烷合成酶的作用下变成血栓烷A2(TXA2)(图11-23)
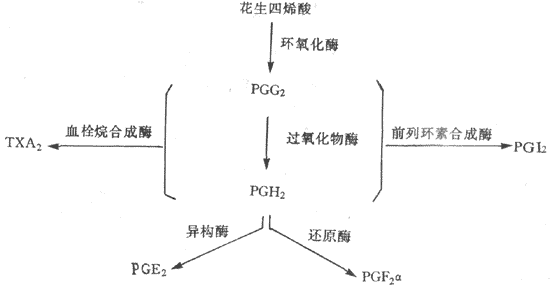
图11-23 体内主要前列腺素的合成途径
另外,花生四烯酸在脂氧化酶的作用下,形成5-氢过氧酸,进而被代谢生成白三烯。
PG在体内代谢极快,除PGI2外,经过肺和肝被迅速降解灭活,在血浆中的半衰期公为1-2min。一般认为,PG不属于循环激素,而是在组织局部产生和释放,并对局部功能进行调节的组织激素。
PG的生物学作用极为广泛而复杂,几乎对机体各个系统的功能活动均有影响。例如,由血小板产生的TXA2,能使血小板聚集,还有能使血管收缩的作用。相反,由血管内膜产生PHG2,能抑制血小板聚集,并有舒张血管的作用。PGE2有明显的抑制胃酸分泌的作用,它可能是胃液分泌的负反馈抑制物,PGE2可增加肾血流量,促进排钠利尿。此外,PG对体温调节、神经系统、以及内分泌与生殖均有影响。
参考资料
1.程治平主编 内分泌生理学 北京:人民卫生出版社,1984
2.邓洁英 生长介素 中华内分泌代谢杂志 1988;4:167-171
3.杨义力 糖皮质激素受体的结构和功能生理科学进展1989;20:133-136
4.谭景莹 甾体激素对基因表达的调节作用,生理科学进展1989;20:223-237
5.蔡子微,李健群 甲状腺激素对细胞核RNA合成的影响生理科学进展 1989;20:363-365
6.马青年 近年内分泌学基础研究值得注意的几个方面生理通讯 1992;11:3棗5
7.朱岩,邱学才 糖皮质激素“允许作用”的可能机制生理科学进展1993;24:263棗226
8.Fukayma s etal.Human parthyroid hirmone(PTH)-related protein and human PTH:comparativebiological activities on human bone cells and bone resorption.Endocrinology1988;123:2841_2848
9.Samuels HH et al.Regulation ofgene expression by thyroid hormone.Ann Rev Physiol 1989;51:623_639
10.Burnstein KL,CidlowskiJA,Regulation of gene expression by glucocorticoids.Ann Rev Physiol1989;51:683_639
11.Winer LM et al. Basal plasmagrowth hormone levels in man: new evidence for rhythmicity of growth hormonesecretion. J Clin Endocrinol Metab 1990;70:1678-11686
12.Emiliano C et al.Human growthhormone and human aging.Endocrinol Rev 1993;14:20-39
第十二章 生殖
生物体生长发育到一定阶段后,能够产生与自己相似的子代个体,这种功能称为生殖(reproduction),任何生物个体的寿命都是有限的,必然要衰老、死亡。一切生物都是通过产生新个体来延续种系的,所以生殖是动物绵延和繁殖种系的重要生命活动。在高等动物,生殖是通过两性生殖器官的活动来实现的,生殖过程包括生殖细胞(精子和卵子)的形成过程,交配和受精过程以及胚胎发育等重要环节。
第一节 男性生殖
男性主要生殖器官为睾丸,此外还有附睾、输精管、精囊腺、前列腺、尿道球腺、阴茎等附属性器官。
一、睾丸的功能
(一)睾丸的生精作用
睾丸由曲细精管与间质细胞组成。曲线精管上皮又由生精细胞和支持细胞构成。原始的生精细胞为精原细胞,肾贴于曲细精管的基膜上,从青春期开始,精原细胞分阶段发育形成精子,精子生成的过程为:精原细胞→初级精母细胞→次级精母细胞→精子细胞→精子。在曲细精管管壁中,各种不同发育阶段的生精细胞是顺次排列的即由基膜至管腔,分别为精原细胞、初级精母细胞、次级精母细胞、精子细胞、分化中的精子,直至成熟精子脱离支持细胞进入管腔,从精原细胞发育成为精子约需二个半月。
支持细胞为各级生殖细胞提供营养,并起着保持与支持作用。为生精细胞的分化发育提供合适的微环境,支持细胞形成的血睾屏障防止生精细胞的抗原物质进入血液循环而引起免疫反应。
精子生成需要了适宜的温度,阴囊内温度较腹腔内温度低2℃左右,适于精子的生成。在胚发育期间,由于某种原因睾丸不降入阴囊而停留在腹腔内或腹股沟内,称隐睾症,则曲细精管不能正常发育,也无精子产生。如果对发育成熟的动物睾丸进入加温处理,或施行实验性隐睾术,则可观察到生精细胞退化萎缩。
新生的精子释入曲线精管管腔内,本身并没有运动能力,而是靠小管外周肌样细胞的收缩和管腔液的移动运送至附睾内。在附睾内精子进一步成熟,并获得运动能力。附睾内可贮存小量的精子,大量的精子则贮存于输畏精管及其壶腹部。而性活动中,通过输精管的蠕动把精子运送至尿道。精子与附睾、精囊腺、前列腺和尿道球腺的分泌物混合形成精液,在性高潮时射出体外。正常男子每次射出精液约3-6ml,每毫升精液约含二千万到四亿个精子,少于二千万精子,不易使卵子受精。
(二)睾丸的内分泌作用
1.雄激素睾丸间质细胞分泌雄激素,主要为睾酮(testosterone,T)。
(1)睾酮的合成与代谢 :睾酮是C-19类固醇激素。在间质细胞内,胆固醇经羟体、侧链裂解形成孕烯醇酮,再经17-羟化并脱去侧链,形成睾酮。睾酮在其靶器官(如附睾和前列腺)内,被5α-还原酶还为为双氢睾酮,再与靶细胞内的受体结合而发挥作用。睾酮也可以芳香化酶作用下转变为雌二醇。
正常男性在20-50岁,睾丸每日约分泌4-9mg睾酮,血浆睾酮浓度为22.7±4.3nmol/L。50岁以上随年龄增长,睾酮的分泌量逐渐减少。
血液中97%-99%的睾酮与血浆蛋白结合,只有1%-3%睾酮是游离的。在血浆中存在一咱与睾酮有很高亲和力的蛋白质,是β球蛋白,分子量为44000-80000。约有30%睾酮与这种球蛋白结合,它也可结合雌激素,故将这咱球蛋白称为性激素结合球蛋白(sex hormone-binding globulin,SHBG)约68%睾酮与血浆白蛋白结合。睾酮主要在肝被灭活,以17-氧类固醇结合型由尿排出,少量经粪便排出。
(2)睾酮的生理作用:主要有以下方面作用:①维持生精作用,睾酮自间质细胞分泌后,可经支持细胞进入曲细精管,睾酮可直接或先转变为活性更强的双氢睾酮,与生精细胞的雄激素受体结合,促进精子的生成。支持细胞在FSH的作用下,可产生一咱对睾酮和双氢睾酮亲和性很强的蛋白质,称为雄激素结合蛋白(androgenbinding protein,ABP),ABP与 睾酮或双氢睾酮结合后,转运至曲线精管,提高雄激素在曲细精管的局部浓度,有利于生精过程;②刺激生殖器官的生长发育,促进男性副性征出现并维持其正常状态;③维持正常的性欲;④促进蛋白质合成,特别是肌肉和生殖器官的蛋白质合成,同时还能促进骨骼生长与钙磷沉积和红细胞生成等。
2.抑制素(inhibin)是睾丸支持细胞分泌的糖蛋白激素,由α和β两个亚单位组成,分子量为31000-32000。抑制素对腺垂体的FSH分泌有很强的抑制作用,而同样生理剂量的抑制素对LH分泌却无明显影响。
二、睾丸功能的调节
睾丸曲细精管的生精过程和间质细胞的睾酮分泌均受下丘脑-垂体的调节。下丘脑分泌的GnRH经垂体门脉到达腺垂体,促进腺垂体促性腺激素细胞合成和分泌卵泡细胞刺激素(follicle-stimulatinghormone, FSH)和黄体生成素(luteinizing hormone, LH)。LH主要作用于间质细胞,而FSH主要作用于生精细胞与支持细胞。动物实验证明,幼年动物摘除垂体后,导致睾丸及附性器官不能发育成熟,呈幼稚状态。如果把成年雄发生动物垂体摘除后,睾丸发生萎缩,生精细胞和间质细胞发生退变,数量减少,生精过程停止,睾酮分泌减少,附性器官也发生萎缩。如果给拉除垂体的动物及早补充垂体促性腺激素,则上述现象可以避免或逆转。毁损下丘脑GnRH神经元所在部位,或下丘脑病变涉及这些区域,将使睾丸萎缩,功能丧失。
睾丸间质细胞膜上存在LH受体。LH与间质细胞膜上的LH受体结合,激活腺苷酸环化酶,促进细胞内cAMP的生成。CAMP再激活依赖cAMP的蛋白激酶,促进蛋白质磷酸化过程,产生碗酸蛋白,从而使胆固醇进入线粒体内合成睾酮,所以LH又称间质细胞刺激素interstitial cell stimulatinghormone,ICSH)。当轿中睾酮达到一定浓度后,便可作用于下丘脑和垂体,抑制GnRH分泌,进而抑制LH的分泌,产生负反馈调节作用,可使血中睾酮浓度稳定在一定水平。
LH与FSH对生精过程都有调节作用,LH的作用是通过睾酮实现的。生精过程受FSH与睾酮的双重控制。大鼠实验表明,FSH起着始动生精的作用,而睾酮则有维持生精的效应。支持细胞膜上存在FSH受体,FSH与受体结合后,经cAMP-蛋白激酶系统,促进支持细胞蛋白质合成,这些蛋白质中,可能有启动精子生成的成分。在FSH作用下,促进支持细胞分泌ABP,ABP与睾酮和双氢睾酮结合转运至曲细精管内,提高曲细精管内雄激素的局部浓度有利于生精过程。实验证明,FSH能刺激支持细胞分泌抑制素,而抑制素对腺垂体的FSH分泌有负反馈调节作用。此外,FSH还可激活支持细胞内的芳香化酶,促进睾酮转变为雌二醇,雌二醇对睾丸的活动也有调节作用,它可降低腺垂体对GnRH的反应性,并可能作用于间质细胞,在局部调节睾酮的分泌。
综上所述,一方面下丘脑-垂体调节睾丸的功能;另一方面睾丸分泌的激素又能反馈调节下丘脑和垂体的分泌活动。下丘脑、垂体、睾丸在功能上密切联系,互相影响,上下统一,称为下丘脑-垂体-睾丸轴(图12-1)。此外,睾丸支持细胞与间质细胞之间,还能以旁分泌的方式进行局部调节。
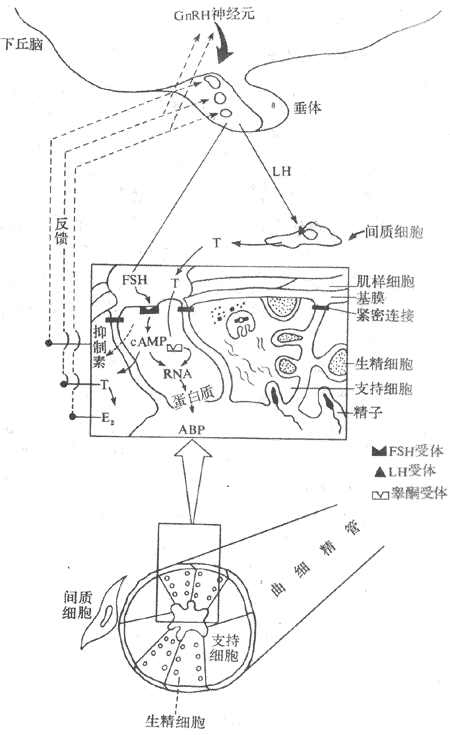
图12-1 下丘脑-垂体-睾丸轴活动的调节
T:睾酮 E2:雌二醇 ABP:雄激素结合蛋白
第二节 女性生殖
女性的主要生殖器官是卵巢,此还还有输卵管、子宫、阴道及外阴等附属性器官。卵巢的功能是产生卵子和分泌激素。
一、月经与排卵及激素调节
在青春期前,卵巢激素的分泌量虽然不大,但由于下丘脑GnRH神经元对卵巢激素反馈抑制作用的敏感性较高,而且GnRH神经元尚未发育成熟,所以GnRH的分泌很少,腺垂体FSH与LH分泌以及卵巢的功能也相应处于低水平状态。至青春期,下丘脑GnRH神经元发育成熟,对卵巢激素的反馈抑制作用的敏感性也明显降低,GnRH的分泌增加,FSH和LH分泌也之时差加,卵巢功能开始活跃,呈现周期性变化,表现为卵泡的生长发育、排卵与黄体形成,周而复始,在卵巢甾体激素周期性分泌的影响睛,子宫内膜发生周期性剥落,产生流血现象,称为月经(menstruation),所以女性生殖周期称为月经周期(menstrual cycle)。哺乳动物也有类似周期,称为动性周期。
卵巢与子宫的周期性变化,是在下丘脑-垂体-卵巢轴的调控下完成的。卵巢的周期性变化是月经周期形成的基础。习惯上将卵巢周期分为卵泡期与黄体期两个阶段。
(一)卵泡期
卵泡的生成发育从原始卵泡开始。人每次月经周期通常只有一个原始卵泡在激素的调控下发育成熟,原始卵泡经初级卵泡与次级卵泡期,最后发育为排卵前卵泡(成熟卵泡)。原始卵泡发育到初级卵泡的早期,不受垂体促性腺激素的控制,其发育取决于卵泡本身的内存因素。到初级卵泡发育晚期,颗粒细胞上出现了FSH受体,内膜细胞上出现了LH受体。到次级卵泡期,颗粒细胞上出现了FSH受体数量进一步增加,FSH在雌激素的协高作用下,诱导颗粒细胞出现LH受体,并随着卵泡发育成熟,颗粒细胞与内膜细胞上的LH受体不断增加。从初级卵泡发育阶段开始,卵泡接受垂体促性腺激素的控制,促使其发育成熟。
卵泡期开始时,血中雌激素与孕激素浓度均处于低水平,对垂体FSH与LH分泌的反馈抑制作用较弱,血中FSH表现逐渐增高的趋势,一二日后LH也有所增加。近年发现,卵泡液中存在一种促进FSH分泌的蛋白质,称为促FSH释放蛋白(FSH-releasing protein),可能对FSH的增加起一定的作用。生长发育的卵泡颗粒细胞上,除FSH受体增加外,还出现胰岛素样生长因子(IGF)及上皮生长因子(EGF)等与细胞增殖有关因子的受体,在FSH与各生长因子的作用下,颗粒细胞明显发育与分化,并产生芳香化酶,可将内膜产生并弥散转运至颗粒细胞的雄激素(主要为雄烯二酮)转变为雌激素。LH与内膜细胞上的LH受体结合,通过cAMP-蛋白激酶系统,使胆固醇转变为雄激素。内膜细胞产生雄激素,而在颗粒细胞转变为雌激素,称为雌激素分泌的双重细胞学说(图12-2)。
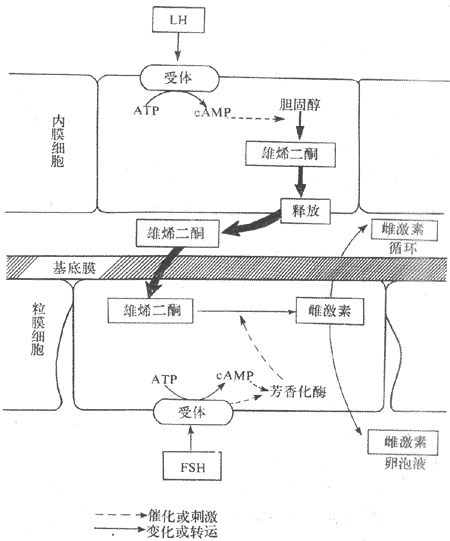
图12-2 雌激素分泌的双重细胞学说示意图
排卵前一周左右,卵泡分泌的雌激素明显增多,血中的浓度迅速上升,与此同时,血中FSH的水平有所下降,这是因为:①雌激素选择地对FSH分泌产生抑制作用;②颗粒细胞产生的卵泡抑制素也对FSH分泌发挥抑制作用。值得指出的是FSH浓度暂时处于低水平,但雌激素浓度并不因此而减少,却反而持续增加,其原因是雌激素可加强内膜细胞的分化与生长,可使LH受体数量增加,从而加强合成雄激素及转变为雌激素的过程。
至排卵前一天左右,血中雌激素浓度达到顶峰,在其作用下,下丘脑增强GnRH分泌,GnRH经垂体门脉转运至腺垂体,刺激LH与FSH的分泌,以LH的分泌增加最为明显,形成LH高峰(LH surge)。若事先用抗激素血清处理动物,则LH峰不再出现。毁损恒河猴下丘视前区后,LH峰也消失。所以,LH峰是由雌激素高峰所诱导出现的,雌激素这种促进LH大量分泌的作用,称为雌激素的正反馈效应。
在LH峰出现之前,卵母细胞已基本发育成熟,但由于包围卵母细胞的颗粒细胞分泌一种卵母细胞成熟抑制因子(oocyte maturation inhibitor,OMI),使卵母细胞成熟分裂中断于前期。LH峰出现的瞬间,高浓度的LH立即抵消OMI的抑制作用,卵母细胞恢复中断了的成熟分裂。成熟卵泡向效巢表面突出,形成透明的卵泡小斑。LH在孕酮的配合下,使卵泡壁溶解酶(如纤溶酶与原酶等)活性增加,导致卵泡壁溶化和松懈。此外LH又可使卵泡分泌前列腺素,后者促使卵泡壁肌样细胞收缩,于是卵细胞与附着的透明带,放射冠从破裂的卵泡壁处被排出,定过程称为排卵(ovulation)。排出的卵子旋即被输卵管伞捕获被抑制,而注射外源LH或HCG(见后文)则能诱发排卵,可见LH峰是控制排卵发生的关键性因素。
在人类,每个月经周期起初有15-20个原始卵泡同时开始生长发育,但是通常只有1个卵泡发育成为优势卵泡,最后发育成熟并排卵,其它卵泡均先后退化并形成闭锁卵泡,其原因尚不十分清楚。
在卵泡期中,子宫内膜也发生相应的变化,主要表现为内膜增厚,腺体增多并变长,此期长为增生期。
(二)黄体期(排卵后期)
卵细胞排出后残余的卵泡壁内陷,血管破裂,血液进入腔内凝固,形成血体。血液被吸收后,大量新生血管长入,血体转变为一个血管丰富的内分泌腺细胞团,外观呈黄色,故称为黄体(corpus luteum)。在LH作用下,颗粒细胞与内膜细胞分别转化为粒黄体细胞与膜黄体细胞。LH通过cAMP-蛋白激酶系统,促使黄体细胞分泌大量的孕激素与雌激素,血中孕酮与雌二醇浓度因而明显升高。在月经期中,雌激素发生二次升高,第一次升高发生在卵泡期,第二次升高发生在黄体期,但第二次升高的程度稍低于第一次。在黄体期,料主水平的雌激素有增加黄体细胞上LH受体的作用,故有利于LH促进孕酮的合成,使孕酮维持于高水平。孕酮和雌激素浓度增加,将使下丘脑与腺垂体受到抑制, GnRh 释放减少。FSH与 LH在血中浓度相应下降。
在黄体期,子宫内膜在雌激素作用的基础上又接受孕激素的刺激,内膜细胞体积增大,糖原含量增加腺管由直变弯,分泌含糖原的粘液,故称分泌期。在子宫的分泌期,一切为妊娠作好准备,“迎接”受精卵子。若不受孕,黄体的寿命为12-15天,黄体即退化,血中孕激素与雌激素浓度明显下降,子宫内膜血管发生痉挛性收缩,随后出现子宫内膜脱落与流血,出现月经。雌激素与孕激素分泌减少,使腺垂体FSH与LH的分泌又开始增加,重复另一周期。如怀孕,胎盘分泌绒毛膜促性腺激素(CG),使黄体功能继续维持一定时间,适应妊娠的需要。(图12-3)。
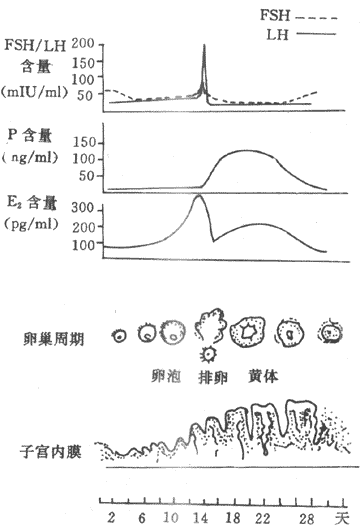
图12-3 月经周期血中FSH、LH、E2及P含量的变化
E2:雌二醇 P:孕酮
二、卵巢的内分泌功能
卵巢分泌的雌激素主要为雌二醇(estradiol,E2),孕激素主要为孕酮(progesterone,P)。此外,卵巢还分泌少量的雄激素。雌二醇是C-18类固醇激素,孕酮是C-21类固醇激素。
血中70%雌二醇与SHBG结合,25%与血浆白蛋白结合,其余为游离型;孕酮在血中约有48%与CBG结合,约有50%与血浆白蛋白结合,其余为游离型,雌二醇和孕酮主要在肝降解,雌三醇是雌二醇主要代谢产物,而孕二醇是孕酮的主要降解产物。这些代谢产物与葡萄糖醛酸或硫酸结合,随尿排出体外。
下面简述雌激素和孕激素的生理作用。
(一)雌激素
雌激素主要的作用是促进女性生死器官的发主育和副性征的出现,并维持在正常状态。此外,雌激素对代谢也有明显的影响
1.对生殖器官的作用 雌激素与卵巢、输卵管、子宫以及阴道粘膜上靶细胞受体结合,引起细胞DNA、RNA和蛋白质合成增加,促进细胞分裂与生长,从而使上述这些靶器官生长发育,并维持其正常功能。如在青春期前雌激素过少,则生殖器官不能正常发育;雌激素过多,则会出现早熟现象。
(1) 卵巢:雌激素除了通过反馈调节经下丘脑-垂全间接影响卵巢活动外,对 卵巢 也有直接作用。实验证明,卵巢组织培养时,雌激素可加速其生长,雌激素可协同FSH促进卵泡发育。FSH在雌激素的协同下,诱发并增加卵泡上LH受体,从而使卵泡对LH的敏感性增加。可见,排卵前的雌激素高峰一方面通过正反馈诱导LH峰的出现;另一方面协同FSH使卵泡上的LH受体增加,有利于LH与期受体结合,并诱发排卵。因此,雌激素是卵泡发育成熟并抑卵不可缺少的调节因素。
(2)输卵管:雌激素促进输卵管上皮细胞增生,分泌细胞、纤毛细胞与平滑肌细胞活动增强促进输卵管运动,有利于精子与卵子的运行。
(3)子宫:雌激素促进子宫发育,内膜发生增生期的变化。雌激素也促进子宫肌的增生,使肌细胞内肌纤蛋白和肌凝蛋白的含量增加。在雌激素的作用下,子宫肌的兴奋性增高,提高子宫肌对催产素的敏感性。
在雌激素的作用下,子宫颈分泌大量清亮、稀薄的粘液,其中的粘蛋白沿宫纵行排列,有利于精子穿行。
(4)阴道:雌激素可使阴道粘膜基底细胞分裂周期缩短,上皮细胞增生,糖原含量增加,表浅细胞角化,粘膜增厚并出现皱折。糖原分解使阴道呈酸性(pH4-5),利于阴道乳酸菌的生长,从而排斥其它微生物的繁殖,所以雌激素能增强阴道的抵抗力,随着人月经周期与动物动性周期的变化,阴道涂片所显示的粘膜脱落上皮细胞类型,也出现周期性变化。在雌激素的作用下,涂片以表浅细胞为主,雌激素水平愈高,则表浅细胞的角化程度愈明显。
2.对乳腺和副性征的影响 雌激素刺激乳腺导管和结缔组织增生,促进乳腺发育,并使全身脂肪和毛发分布具有女性特征,音调较主,骨盆宽大,臀部肥厚。
3.对代谢的作用 雌激素对代放的作用比较广泛,主要有:①雌激素刺激成骨细胞的活动,而抑制破骨细胞的活动,加速骨的生长,促进钙盐沉积,并能促进骨骺软骨的愈合,因而在青春
■[此处缺少一些内容]■
孕酮使宫颈粘液减少而变稠,粘蛋白分子弯曲,交织成网,使精子难以通过。
2.乳腺在雌激素作用的基础上,孕激素主要促进乳腺腺泡发育,并在妊娠后为泌乳作用好准备。
3.产热作用女性基础体温在排卵前先出现短暂降低,而在排卵后升高0.5℃左右,并在黄体期一直维持在此水平上,临床上常将这一基础体温的双相变化,作为判定排卵的标志之一。妇女在绝经或卵巢摘除后,这种双相的体温变化消失,如果注射孕酮则可引起基础体温长高,因此认为基础体温的升高与孕酮有关。
(三)雄激素
女子体内有少量的雄激素,是由卵泡内膜细胞和肾上腺皮质网状带细胞产竹。适量的雄激素配合雌激素可刺激阴毛及腋毛的生长,女子雄激素过多时,可引起男性化与多毛症。雄激素能增强女子的性欲,维持性快感,这可能由于它促进阴蒂的发育并提高其敏感性,或是由于它对中枢神经系统的作用。
三、妊娠
妊娠是新个体产生的过程,包括受精、着床、妊娠的维持、胎儿的生长以及分娩。
(一)受精
精子与卵子在输卵管壶腹部相遇而受精,精子与卵子相融合时称为受精卵。每一个精子和卵子各含23个染色体,受精卵则含有23对染色体。因此具有父母双方的遗传特性。
射入阴道的精子进入输卵管与卵子相遇的过程比较复杂。精子有运动不完全依靠本身的运动,宫颈、子宫和输卵管对精子的运动都起到一定的作用。精液射入阴道后穹窿后,很快(约1min)就变成胶冻样物质,使精液不易流出体外,并有暂进保持精子免受酸性阴道液的破坏作用。但是,阴道内的精子绝大部分被阴道内的酶杀伤失去活力,存活的精子随后又遇到宫颈粘液的拦截。月经中期在雌激素的作用下,宫颈粘液清亮、稀薄,其中的粘液蛋白纵行排列成行,有利于精子的穿行,而黄体期的孕激素的作用下,宫颈粘液变得粘稠,粘液蛋白卷曲,交织成网,使精子能以通过。总之,宫颈作为精子在女生生殖道内要通过的第一个关口,它在排卵时,为精子的穿行提供了最优越的条件。一部分精子靠本身的运动及射精后引起的子宫收缩,进入子宫腔内。精液中含有很高浓度的前列腺素,可刺激子宫发生收缩,收缩后的松驰造成宫腔内负压,可把精子吸入宫腔。精子进入输卵管后,在其中的运行主要受输卵管蠕动的影响。月经中期在雌激素的作用下,输卵管的蠕动由子宫向卵巢方向移行,推动精子由峡部运动至壶腹部。黄体期分泌的大量孕酮能抑制输卵管的蠕动。一次射精虽能排出数以亿计的精子,但最后能到达受精部位的只有15-15个精子,到达的时间约在性交后30-90min。精子在女性生殖道内的受精能力大约只能保持48h。
大多数哺乳动物和人类,精子必须在雌性生殖道内停留一段时间,方能获得使卵子受精的能力,称为精子获能(capacitation)。精子经过在附睾中的发育,已经具备了受精能力,但在附睾与精浆中存在去获能因子。它使精子的受精能力受到了抑制。当精子进入雌性生殖道内后,能解除去获能因子对精子的抑制,从而使其恢复受精能力。获能的主要场所是子宫,其次是输卵管,宫颈也可能有使精子获能的作用。
精子与卵子在输卵管壶腹部相遇后尚不能立即结合,顶体外膜与精子头部的细胞膜首先融合,继之破裂,形成许多小孔,释放出顶体膜,以溶解卵子外围的放射冠及透明带,这一过程称为顶体反应。顶体酶包含多种蛋白水解酶,如放射冠穿透酶可使放射冠的颗粒细胞松解,脱离卵细胞外围。颗粒细胞脱落后,在透明带周围仍残存一层放射冠基质,可在透明质酸酶的作用下,这些基质被水解,使透明带暴露出来。透明带为糖蛋白,在顶体蛋白酶的作用下,使透明带发生部分水解,促进精子能突破透明带的一个局限区到达并进入卵细胞内,在一个精子穿越透明带后,精子与卵细胞接触,激发卵细胞发生反应,主要是位于卵细胞周边部的皮质颗粒包膜与卵细胞膜逐渐融合、破裂,并向卵周隙释放其内容物,有人认为,释放物作用于透明带,使其变质,或其种物质起封锁透明带的作用,使其他精子难以再穿越透明带进入卵细胞内。精子进入卵细胞后立即激发卵细胞完成第二次成熟分裂,并形成第二极体。进入卵细胞的精子,其尾部迅速退化,细胞核膨大形成雄性原核,随即与雌性原核融合,形成一个具有46染色体的受精卵。
受精卵在输卵管的蠕动和纤毛的作用下,逐渐运行至子宫腔。受精卵在运行途中,一面移动,一面进行细胞分裂,经胚球和桑椹期阶段,发育为胚泡(blastocyst)。在受精后第四五天,桑椹胚或早期胚泡进入子宫腔,桑椹胚在子宫腔内继续分裂变成胚泡。胚泡在子宫腔内停留2-3天,胚泡外面的透明带变薄,胚泡可以直接从子宫内膜分泌的液体中吸收营养。
(二)着床
着床是胚泡植入子宫内膜的过程,经过定位、粘着和穿透三个阶段。着床成功的关键在于胚泡与子宫内膜的同步发育与相互配合。胚泡的分化与到达子宫的时间必需与子宫内膜发育程度相一致。胚泡过早或过迟到达子宫腔,将使着床纺明显降低,甚至不能着床。在着床过程中,胚泡不断地发出信息,使母体能识别妊娠发生相应的变化。胚泡可产生多种激素和化学物质,如绒毛膜促性腺激素,它能刺激卵巢黄体转变为妊娠黄体,继续分泌妊娠需要的孕激素。近年发出,受精24h的的受精卵便可产生早孕因子(early pregnancy factor),它能抑制母体淋巴细胞的功能,使胚泡免遭母体排斥。检测早孕因子可进行超早期妊娠诊断。
子宫仅在一个极短的关键时期内允许胚泡着床,此时期为子宫的敏感期或接受期。在此时期内,子宫内膜受到雌激素与孕激素的协同作用,可能分泌某些物质,激活胚泡着床。引起子宫内膜着床反应的机制尚不十分清楚,可能与胚泡子宫内膜产生某种激肽,释放组胺,或与胚泡分泌的蛋白水解酶和产生的CO2有关。胚泡产生的CO2扩散到子宫内膜,再进入子宫的微血管,在胚泡附近形成一个CO2梯度场。CO2可使滋养层细胞和子宫内膜上皮细胞表面的粘蛋白粘性增高,在着床时有胚泡粘着并植入,此外,CO2还能刺激子宫内膜的基质发生蜕膜反应。
(三)妊娠的维持及激素调节
正常妊娠的维持有赖于垂体、卵巢和胎盘分泌的各种激素相互配合,在受精与着床之前,在腺垂体促性腺激素的控制下,卵巢黄体分泌大量的孕激素与雌激素,导致子宫内膜发生分泌期的变化,以适应妊娠的需要。如未受孕,黄体按时退缩,妥激素与雌激素分泌减少,引起子宫内膜剥脱流血;如果受孕,在受精后第六天左右,胚泡滋养层细胞便开始分泌绒毛膜促性腺激素,以的逐渐增多,刺激卵巢黄体变为妊娠黄体,继续分泌孕激素和雌激素。胎盘形成后,胎盘成为妊娠期一个重要的内分泌器官,大量分泌蛋白质激素、肽类激素和类固醇激素。
1.人绒毛膜促性腺激素(human chorionic gonadotropin,HCG) HCG是由胎盘绒毛组织的合体滋养层细胞分泌的一种糖蛋白激素,分子量为45000-50000。HCG分子由α亚单位与β亚单位组成。其α亚单位氨基酸的数量与序列几乎与LH相同,其β亚单位的氨基酸也有很大部分与LH相同,但在β亚单位的羧基端约有30个氨基酸是独特的。因此,HCG与LH的生物学作用与免疫特性基本相似。
卵子受精后第六天左右,胚泡形成滋养层细胞,开始分泌HCG,但其量甚少。妊娠早期形成绒毛组织后,由合体滋养层细胞分泌大量的HCG,而且分泌量增长很快,至妊娠8-10周,HCG的分泌达到高峰,随后下降,在妊娠20周左右降至较低水平,并一直维持至妊娠末。如无胎盘残留,于产后四天轿中HCG消失。在妊娠过程中,尿中HCG含量的动态变化与血液相似。因为HCG在妊娠早期即出现,所以检测母体血中或尿中的HCG,可作为诊断早孕的准确指标。
在早孕期,HCG刺激卵巢黄体转变成妊娠黄体,妊娠黄体的寿命只有10周左右,以后便发生退缩,与此同时胎盘分泌孕激素和雌激素,逐渐接替了妊娠黄体的作用。
2.其他蛋白质激素和肽类激素胎盘还可分泌人绒毛膜生长素、绒毛膜促甲状腺激素、促肾上腺皮质激素GnRH,以及β-内啡肽等。
人绒毛生长素(human chorionicsomatomammotropin,HCS)为合体滋养层细胞分泌的单链多肽,含191个氨基酸残基,其中96%与人生长素相同,因此具有生长素的作用,可调节母体与胎儿的糖、脂肪与蛋白质代谢,促进胎儿生长。最初发现HC时,证明它对动物有很强的催乳作用,故命名为人胎盘催乳不比(humanplacental lactogen,HPL)。后来的研究证明,HPL对人几乎没有催乳作用,而主要是促进胎儿生长,因此在国际会议上将其定名为HCS。
3.类固醇激素胎盘本身不能独立产生类固醇激素,需要从母体或胎儿得到前身物质,再加工制成孕激素与雌激素。
(1)孕激素:由胎盘合体滋养层细胞分泌,胎盘不能将醋酸盐转变为胆固醇,而能将自母体进入胎盘的胆固醇变为孕烯醇酮,然后再转变为孕酮。胎儿肾上腺虽能合成孕烯醇酮,但由于缺乏3β-醇甾脱氢酶,故不能将孕烯醇酮转变为孕酮,而胎盘此种酶的活性很强,能把来自胎儿和母体的孕烯醇酮转变为孕酮。
在妊娠期间,母体血中孕酮浓度随着孕期的增长而稳步上升,在妊娠10周以后,由胎盘代替卵巢持续分泌孕酮,血中孕酮迅速增加,至妊娠足月时达高峰,平时浓度可达600nmol/L。
(2)雌激素:由母体和胎儿肾上腺产生的脱氢异雄酮硫酸盐,进入胎盘最后转变为雌酮和雌二醇,但生成量极少。胎盘分泌的雌激素主要为雌三醇,其合成的途径是,胎儿肾上腺的脱氢异雄硫酸盐先在胎儿肝中羟化,形成16α-痉脱氢异雄酮硫酸盐,然后随血液进入胎盘,在胎盘内脱去硫酸基,成为16α-羟脱氢异雄酮,再经芳香化酶的作用,转化为雌三醇。由此可见,雌三醇的生成是胎儿、胎盘共同参与制造的,故把两者称为胎儿-胎盘单位。检测母体血中雌三醇的含量多少,可用来判断胎儿是否存活。
(四)分娩与授乳
有关分娩与授乳的主要生理内容已在讨论催乳素与催产素时有所述及,不再重复。
(马青年)
参考资料
1.徐丰彦,张镜如主编 《人生生理学》 北京:人民卫生出版社,1989
2.王淑贞主编 《实用妇产科学》 北京 人民卫生出版社,1987
3.马青年,等,《实用男性学》 天津,天津科学技术出版社,1988
4.曹泳清,陈幼珍,《兔胚泡肽粘着作用的研究》 生殖医学杂志,1998;2:55
5.Eli YA etal.SmC/Insulin-like growth factor andgranulosa cells.Endocrinology1988;122:194-202
6.Conley AJ,FordSP.Direct luteotrophic effect of oewtradiol-17β on pig corpora lutea.J ReprodFert 1989;87:125-131
生理学索引
索引
A
| 暗适应 | Dark adaptation | 285 |
| 暗视觉 | Scotopic vision | 280 |
B
| 被动转运 | Passive transport | 13 |
| 白细胞 | Leukocyte | 62 |
| 背侧呼吸组 | dorsal respiratory group | 174 |
| 搏功 | Stroke work | 90 |
| 变力作用 | Inotropic action | 132 |
| 变时作用 | Chronotropic action | 132 |
| 变传导作用 | Dromtropic action | 132 |
| 比顺应性 | Specific compliance | 156 |
| 补吸气量 | Inspiratoryreserve volume | 159 |
| 补呼气量 | Expiratory reserve volume | 159 |
| 不完全强直收缩 | Incomplete tetanus | 50 |
| 不感蒸发(不显汗) | Insensible perpiration | 232 |
| 编码(作用) | Encoding | 269 |
| 边缘系统 | Babinski’s sign | 336 |
| 巴彬斯基征 | Limbic system | 356 |
| α波阻断 | α-boock | 364 |
C
| 出胞 | Exocytosis | 312 |
| 出血时间 | Bleeding time | 68 |
| 刺激 | Stimulus | 25 |
| 超常期 | Supranormal period | 26 |
| 超极化 | Hyperpolarization | 28 |
| 超速驱动压抑 | Overdrive suppression | 108 |
| 初长度 | Initial length | 46 |
| 促红细胞生成素 | Erythropoietin,EPO | 61 |
| 促胰液素 | Secretin | 204 |
| 促甲状腺激素 | Thyroid stimulating hormone,TSH | 382 |
| 促甲状腺激素释放激素 | Thyrotropin-releaxing hormone,TRH | 378 |
| 促性腺激素释放激素 | Gonadotropin-releasing hormone,GnRH | 379 |
| 促肾上腺皮质激素释放激素 | Corticotropin-releasing hormone,CRH | 380 |
| 促肾上腺皮质激素 | Adrenocorticotropin,ACTH | 382 |
| 促阿片-黑素细胞皮质素原 | Pro-opiomelanocrtin,POMC | 380 |
| (阿黑皮素原) | ||
| 长度-张力曲线 | Length-tension curve | 46,93 |
| 长时性记忆 | Long term memory | 359 |
| 长时程增强 | Long-term potentiation | 361 |
| 充盈期(相) | filling period(phase) | 88 |
| 充盈压 | Filling pressure | 92 |
| 层流 | Laminar flow | 116 |
| 湍流 | Turbulence | 116,117 |
| 潮气量 | Tidal volume | 159 |
| 肠抑胃素 | Enterogastrone | 202 |
| 肠-胃反射 | Enterogastric reflex | 202 |
| 肠致活酶 | Enterokinase | 208 |
| 产热 | Production of heat | 230 |
| 传导 | Conduction | 97,231 |
| 传递蛋白 | Transducin | 281 |
| 传入侧支性抑制 | Afferent collateral inhibition | 322 |
| 窗孔结构 | Fenestration | 244 |
| 重吸收 | Reabsoption | 128 |
| 操作条件反射 | Operate conditioned reflex | 358 |
| 催产素 | Oxytocin,OXT | 385 |
| 催乳素 | Prolactin,PRL | 382 |
| 雌二醇 | Estradiol,E2 | 398 |
D
| 单纯扩散 | Simple diffusion | 11 |
| 单收缩 | Single twitch | 49 |
| 单位平滑肌 | Single-unit smooth muscle | 52 |
| 单核细胞 | Monocyte | 62,65 |
| 电-化学势 | Electro-chemical potential | 14 |
| 电压门控通道 | Voltage-gated channel | 19 |
| 电压钳 | Voltage clamp | 32 |
| 电紧张性扩布 | Electrotonic propagation | 37 |
| 电突触 | Electric synapse | 306 |
| 第二信使 | Second messenger | 21,373 |
| 低常期 | Subnormal period | 26 |
| 动作电位 | Action potential | 25,27 |
| 动脉血压 | Arterial blood pressure | J118 |
| 动脉脉搏 | Atrerial pulse | 118 |
| 动-静脉吻合支 | Arteriovenous anastomosis | 126 |
| 动毛 | Kinocilium | 294 |
| 等长收缩 | Isometric contraction | 46 |
| 等张收缩 | Isotonic contraction | 49 |
| 等容收缩期 | Isovolumic systole | 86 |
| 等容舒张期 | Isovolumic diastole | 88 |
| 多单位平滑肌 | Multi-unit smooth muscle | 52 |
| 定向祖细胞 | Committed progenitors | 58 |
| 定比重吸收 | Constant fraction reabsorption | 257 |
| 代偿性间歇 | Compensatory pause | 107 |
| 代谢 | Metabolism | 218 |
| 端压 | End pressrure | 118 |
| 胆囊收缩素 | Cholecystokinin(CCK) | 205 |
| 胆盐 | Bile salt | 206 |
| 胆盐的肠肝循环 | Enterohepatic circulation of bile salt | 207 |
| 对流 | Convection | 231 |
| 毒蕈碱受体 | Muscarinic receptor | 313 |
| 短时性记忆 | Short term memory | 359 |
| 蛋白激酶C | Protein kinase C,PKC | 375 |
E
| 二酰油 | Diacyl glycerol | 22 |
| 二氧化碳解离曲线 | Carbon dioxide dissociation curve | 171,172 |
F
| 反馈 | Feedback | 324 |
| 反射弧 | Rdflex arc | 3,317 |
| 锋电位 | Spike potential | 29 |
| 房室延搁 | Atrioventricular delay | 110 |
| 非自律细胞 | Non-rhythmic cell | 97 |
| 非蛋白呼吸商 | Non-protein respiratory quotient,NPRQ | 221 |
| 非突触性化学传递 | Non-synaptic chemical transmission | 305 |
| 非特异投射系统 | Nonspecific projection system | 327 |
| 非联合型学习 | Non-associative learning | 358 |
| 复活 | Reactivqtion | 106 |
| 肺循环 | Pulmonary circulation | 145 |
| 肺通气 | Pulmonary ventilation | 150 |
| 肺内压 | Intrapulmonary pressure | 154 |
| 肺泡表面活性物质 | Alveolar surfactant | 156 |
| 肺活量 | Vital capacity | 160 |
| 肺总量 | Total lung capacity | 160 |
| 肺泡通气量 | Alveolar ventilation | 160,161 |
| 肺扩散容量 | Pulmonary diffusion capacity | 165 |
| 肺牵张反射 | Pulmonary stretch reflex | 175 |
| 腹式呼吸 | Abdominal breathing | 152 |
| 腹侧呼吸组 | Ventral respiratory group | 174 |
| 分节运动 | Segmentation contraction | 208 |
| 放能反应 | Exergonic reaction | 218 |
| 辐射原则 | Divergence principle | 318 |
| 辐射 | Radiation | 231 |
| 发汗 | Sweating(perspiration) | 232 |
| 翻正反射 | Righting reflex | 341 |
| 防御反应区 | Defense zone | 355 |
G
| 钙调蛋白 | Calmodulin | 51,375 |
| 肝素 | Heparin | 72 |
| 工作(心肌)细胞 | Working(cardiac)cell | 97 |
| 冠脉循环 | Coronary circulation | 143 |
| 功能余气量 | Functional residual volume | 159 |
| 管球反馈 | Tubuloglomerular feedback | 258 |
| 光适应 | Light adaptation | 285 |
| 感觉柱 | Sensory column | 329 |
| 感觉失语症 | Xensory aphasia | 362 |
| 感觉区 | Sensory area | 331 |
| 睾酮 | Testosterone,T | 411 |
H
| 化学门控通道 | Chmically gated channel | 18 |
| 化学感受性反射 | Chemoreceptor rdflex | 137 |
| 化学感受器 | Chemoreceptor | 177 |
| 后电位 | Afterpotential | 29 |
| 后负荷 | Afterload | 46 |
| 后放 | Atfterdischarge | 321 |
| 河豚毒 | Tetrodotoxin | 34 |
| 横桥 | Cross bridge | 43 |
| 横桥联接 | Cross bridge attachment | 94 |
| 滑行学说 | Sliding theory | 42 |
| 红细胞 | Erythrocyte | 57,59 |
| 缓冲神经 | Buffer neerves | 136 |
| 呼吸 | Respiration | 150 |
| 呼气 | Expiration | 152 |
| 呼吸运动 | Respiratory movement | 152 |
| 呼吸困难 | Dysp(o)nea | 152 |
| 呼吸调节中枢 | Pneumotaxic center | 173 |
| 呼吸热量计 | Respiration calorimeter | 219 |
| 呼吸商 | Respiratory quotient | 220 |
| 回漏 | Back-leak | 247 |
| 回返性抑制 | Recurrent inhibition | 322 |
| 壶腹 | Ampulla | 295 |
| 海马环路 | Hippocmpal circuit | 361 |
| 黄体生成素 | Luteinizing hormone,LH | 382 |
| (间质细胞刺激素) | (interstitial cell stimulating hormone,ICSH) | |
| 环-磷腺苷 | Cyclic AMP,cAMP | 373 |
J
| 继发性主动转运 | Secondary active transport | 15 |
| 绝对不应期 | Absolute refractory period | 26 |
| 极化 | Polarization | 28 |
| 局部兴奋 | Local excitation | 36 |
| 局部神经元回路 | Local neuronal circuit | 306 |
| 局部回路神经元 | Local circuit neuron | 306 |
| 肌原纤维 | Myofibril | 41 |
| 肌小节 | Sarcomere | 41 |
| 肌凝蛋白(肌球蛋白) | Myosin | 44 |
| 肌纤蛋白(肌动蛋白) | Actin | 43 |
| 肌钙蛋白(原宁蛋白) | Tropnin | 44 |
| 肌紧张 | Muscle tonus | 336 |
| 肌梭 | Muscle spindle | 37 |
| 集落刺激因子 | Colonystimulating factor,CSF | 62 |
| 浆细胞 | Plasma cell | 63 |
| 巨核细胞 | Megakaryocytes | 66 |
| 交叉配血试验 | Cross-match test | 83 |
| 交换血管 | Exchange vessel | 116 |
| 交互性突触 | Reciprocal synapse | 306 |
| 静息心指数 | Resting cardiac index | 89 |
| 静息张力 | Resting tension | 92 |
| 静脉回心血量 | Venous return | 122 |
| 静毛 | Stereocilia | 294 |
| 激肽释放酶-激肽系统 | Kallikrein-kinin system | 140 |
| 激素 | Hormone | 370 |
| 解剖无效腔 | Anatomical dead space | 161 |
| 基本电节律 | Basic electrical rhythm | 185 |
| 基础代谢 | Basal metabolic rate | 226 |
| 基础代谢率 | Basal metabolic rate | 226 |
| 假饲 | Sham-feeding | 198 |
| 假怒 | Sham rage | 355 |
| 直接测热法 | Direct calorimetry | 219 |
| 菊粉 | Inulin | 263 |
| 简化眼 | Reduced eye | 274 |
| 聚合原则 | Convergence principle | 318 |
| 脊休克 | Spinal shock | 335 |
| 甲状腺素(四碘甲腺原氨酸) | Thyroxin,3,5,3’,5’-tetraiodlthyronine,T4 | 387 |
| 甲状腺球蛋白 | Thyroglobulin,TG | 389 |
| 甲状腺素结合球蛋白 | Thyroxine-binding globulin,TBG | 390 |
| 甲状旁腺激素 | Parathyroid hormone,PTH | 394 |
| 降钙素 | Calcitomin,CT | 396 |
| 焦度 | Diopter | 273 |
K
| 跨膜信号传递 | Transmembrane signaling | 17 |
| 跨膜信号转换 | Transmembrane signal transduction | 18 |
| 跨膜静息电位 | Transmembrane reating potential | 28 |
| 快速(即早)基因 | Immediate-early gene | 24 |
| 快速射血期(相) | Rapid ejection period(phase) | 87 |
| 快痛 | Fast pain | 332 |
| 空间性总和 | Spitial summation | 37 |
| 抗凝物质 | Anticoagulant | 68 |
| 抗利尿激素 | Antidiuretic hormone | 259 |
| 可感蒸发 | Sensible perspiration | 232 |
L
| 酪氨酸蛋白激酶受体 | Tyrosine kinase receptor | 23 |
| 量子式释放 | Quantal release | 39 |
| 粒细胞 | Granulocyte | 62 |
| 淋巴细胞 | Lymphocyte | 62,65 |
| 淋巴液 | Lymph | 130 |
| 冷敏神经元 | Cold-sensitive neuron | 235 |
| 滤过 | Filtration | 129 |
| 滤过裂孔隙 | Filtration slit membrane | 242 |
| 联合型学习 | Associative learning | 358 |
| 裂脑 | Split brain | 363 |
| 卵泡刺激素 | Follicle-stimulating hormone,FSH | 382 |
M
| 膜片钳 | Patch clamp | 30,34 |
| 每搏输出量 | Stroke volume | 89 |
| 每分功(心脏) | Minute work(heart) | 90 |
| 每分输出量 | Minute volume(heart) | 89 |
| 每分通气量 | Minute ventilation volume | 160 |
| 脉搏压 | Pulse pressure | 119 |
| 毛细血管前阻力 | Precapillary resistance | 115 |
| 毛细血管后阻力 | Postcapillary resistance | 116 |
| 毛细血管前括约肌 | Precapillary sphincter | 116 |
| 毛细血管 | Capillary | 116 |
| 慢痛 | Slow pain | 332 |
| 慢波睡眠 | Slow wave sleep | 368 |
N
| 内环境 | Internal environment | 3,54 |
| 内因子 | Inintrinsic factor | 60 |
| 内源性激活途径 | Intrinsic route | 69 |
| 内皮舒张因子 | Endothelium-derived relaxing factor(EDRF) | 139 |
| 内皮缩血管因子 | Endothlium-derived vasoconstrictor factor(EDCF) | 139 |
| 内皮素 | Endothelin | 139 |
| 内脏痛 | Visceral pain | 33 |
| 钠泵 | Sodium pump | 14 |
| 鸟苷酸结合蛋白(G-蛋白) | Guanine nucleotide-binding protein(G-protein) | 22 |
| 凝血因子 | Blood clotting factors | 68 |
| 凝血酶 | Thrombin | 71 |
| 凝集原 | Agglutinogen | 78 |
| 凝集素 | Agglutinin | 78 |
| 粘滞度 | Viscosity | 57 |
| 粘液 | Mucus | 196 |
| 粘液-碳酸氢盐屏障 | Mucus-bicarbonate barrier | 196 |
| 脑缺血反应 | Brain ischemia response | 137 |
| 脑循环 | Cerbral circulation | 146 |
| 脑脊液 | Cerebrospinal fluid | 147 |
| 脑电图 | Electroencephalogram,EEG | 364 |
| 能量代谢 | Energy metabolism | 218 |
| 逆向转运 | Antiport | 247 |
| 逆向转运体(交换体) | Antiporter(exchanger) | 247 |
| 逆行性遗忘症 | Retrograde amnesia | 361 |
| 囊斑 | Macula | 396 |
O
| 呕吐 | vomiting | 202 |
P
| 坪(平台) | plateau | 99 |
| 泊隶叶定律 | Poiseuilli’s law | 117 |
| 平静呼吸 | Eupn(o)ea (quiet breathing) | 152 |
| 排便 | Defecation | 211 |
| 排尿 | Micturition | 269 |
| 排尿欲 | Micturition desire | 266 |
| 排卵 | Ovulation | 415 |
| 皮肤温度 | Skin temperature | 231 |
| 皮质醇 | Cortisol | 398 |
| 皮质类固醇结合球蛋白 | Corticosteroid-binding globulin,CBG | 398 |
| 旁锥体系 | Parapyramidal system | 347 |
| 旁分泌 | Paracrine | 4 |
Q
| 前馈 | Feed-forward | 5 |
| 前负荷 | Preload | 46 |
| 前体细胞 | Precursors | 59 |
| 前裂腺素 | Prostaglandin,PG | 140,409 |
| 去极化(除极) | Depolarzation | 28 |
| 去大脑僵直 | Decerebrate rigidity | 339 |
| 去皮层僵直 | Decorticate rigidity | 340 |
| 去甲肾上腺素 | Norepinephrine,NE | 309,403 |
| “全或无”现象 | “all or none”phenomenon | 30 |
| 期前收缩(期外收缩) | Premature systole(extrasystole) | 107 |
| 起搏电流 | Pacemaker current | 102 |
| 气胸 | Pneumothorax | 154 |
| 气道阻力 | Airway resistance | 158 |
| 气体交换 | Gas eschange | 161 |
| 气体扩散速率 | Diffusion rate of gas | 162 |
| 球管平衡 | Glomerulotubular balance | 257 |
| 醛固酮诱导蛋白 | Aldosterone-induced protein | 260 |
| 醛固酮 | Aldosterone | 260,397 |
| 曲张体 | Varicosity | 52 |
| 牵涉痛 | Referred pain | 333 |
| 牵张反射 | Stretch reflex | 336 |
| 屈肌反射 | Flexor reflex | 336 |
R
| 人白细胞抗原 | Human leudocyte antigen,HLA | 82 |
| 人绒毛膜促性腺激素 | Human chorionic gonadotropin,HCG | 420 |
| 入胞 | Endocytosis | 16 |
| 容量血管 | Capacitance vessel | 116 |
| 容量感受器 | Volume receptor | 136 |
| 蠕动 | Peristalsis | 193,209 |
| 绒毛 | Vullus | 419 |
| 绒毛收缩素 | Villikinin | 214 |
| 乳糜微粒 | Chylomicron | 216 |
| 热价 | Thermal equivalent | 220 |
| 热喘呼吸 | Panting | 232 |
| 热敏神经元 | Warm-sensitive neuron | 235 |
| 闰绍细胞 | Renshaw cell | 308 |
S
| 上行运动神经元麻痹 | Upper motorneuron paralysis | 348 |
| 上行抑制系统 | Ascenging inhibitory system | 369 |
| 神经垂体激素运载蛋白 | Neurophysin | 385 |
| 神经调节 | Neuroregulation | 3.130 |
| 神经递质 | Neurotransmitter | 307 |
| 神经激素 | Neurohormone | 370 |
| 神经分泌 | Neurocrine | 370 |
| 三磷酸肌醇 | Inositol triphosphate | 22 |
| 三碘甲腺原氨酸 | 3,5,3’-triiodothyronine,T3 | 387 |
| α受体 | α-receptor | 314 |
| β受体 | β-receptor | 314 |
| 时间性总和 | Tempoal summation | 37 |
| 时间肺活量 | Timed vital capacity | 160 |
| 四乙基铵 | Tetraethylammonium | 34 |
| 输血 | Blood transfusion | 77 |
| 嗜酸性粒细胞 | Eosnophils | 64 |
| 嗜碱性粒细胞 | Basophils | 64 |
| 射血分数 | Ejection fraction | 90 |
| 射血期(相) | Ejection period(phase) | 87 |
| 收缩能力(肌) | Contractility | 49 |
| 收缩(心缩期) | Systole | 86 |
| 收缩压 | Systolic pressure | 120 |
| 舒张压 | Diastolic pressure | 120 |
| 舒张(心舒)期 | Diastole | 88 |
| 舒血管纤维 | Vasodilator fiber | 132 |
| 水肿 | Edema | 129 |
| 水利尿 | Water diuresis | 260 |
| 缩血管神经纤维 | Vasoconstrictor fiber | 132 |
| 肾素-血管紧张素系统 | Renin-angiotensin sysem | 137 |
| 肾血流量 | Renal blood flow | 241 |
| 肾小球滤过率 | Renin-angiotensin-aldosterone system | 241.242.262 |
| 肾血浆流量 | Renal plasma flow | 246 |
| 肾素-血管紧张素-醛固酮系统 | Renin-angiotensig-aldosterone system | 260 |
| 肾上腺素 | Epinephrine,E | 403 |
| 渗透浓度 | Osmolality | 253 |
| 渗透性利尿 | Osmotic diuresis | 260 |
| 顺应性 | Compliance | 155 |
| 顺向性遗忘症 | Anterograde ameesia | 361 |
| 深吸气量 | Inspiratory capacity | 159 |
| 生理无效腔 | Physiological dead space | 161 |
| 生长抑素 | Growth hormone release-inhibiting hormone GHRIH | 380 |
| 生物节律 | Biorhythm | 355 |
| 生长素 | Growth hormone,GH | 382 |
| 生长介素 | Somatomedin,SM | 383 |
| 生长素释放激素 | Growth hormone releasing hormone,GHRH | 380 |
| 生殖 | Reproduction | 411 |
| 升压素 | Vasopressin,VP | 385 |
| 食团 | Bolus | 192 |
| 食管-胃括约肌 | Esophageal-gastric sphincter | 193 |
| 食糜 | Chyme | 201 |
| 食物的特殊动力作用 | Specific dynamic action | 225 |
| 散热 | Loss of heat | 230 |
| 适宜刺激 | Adequate stimulus | 268 |
| 适应(感觉器) | Adaptation | 271 |
| 视敏度 | Visual acuity | 274 |
| 视杆细胞 | Rod | 278 |
| 视锥细胞 | Cone | 278 |
| 视紫红质 | Rhodopsin | 279 |
| 视蛋白 | opsin | 279 |
| 视黄醛 | Retinal | 279 |
| 视野 | Visual field | 286 |
| 视网膜电图 | Elecrtoretinogram | 286 |
| 权突-树突型抑制 | Dendrodendrtic inhibition | 324 |
| 梭内肌纤维 | Intrafusal fiber | 337 |
| 梭外肌纤维 | Extrafuxla fiber | 337 |
| 失活 | Inactivation | 33 |
| 失写症 | Agraphia | 362 |
| 失读症 | Alexia | 362 |
| 失用症 | Apraxia | 363 |
T
| 褪黑素 | melatonin | 408 |
| 体液调节 | Humoral regulation | 233,353 |
| 体温 | Body temperature | 218,228 |
| 体表面积 | Body surface area | 226 |
| 通透性 | Permeability | 11 |
| 通气/血流比值 | Ventlation-perfusion ratio | 164 |
| 通血毛细血管 | Preferential channel | 125 |
| 吞饮 | Pinocytosis | 193 |
| 跳跃式传导 | Saltatory conduction | 38 |
| 调定点 | Set point | 353 |
| 调质 | Modulator | 311 |
| 条件反射 | Conditioned reflex | 3 |
| 碳酸酐酶 | Carbonic angydrase | 195 |
| 突触小体 | Synaptic knob,synaptic bouton | 303.304 |
| 突触前受体 | Presynaptic receptor | 315 |
| 突触后抑制 | Postsynaptic inhibition | 321 |
| 突触前抑制 | Presynaptic inhibition | 321 |
| 特异投射系统 | Specific projection system | 327 |
W
| 稳态 | homeostasis | 3,55 |
| 完全强直收缩 | Complete tetanus | 50 |
| 外源性激活途径 | Extrinsic route | 69 |
| 外周阻力 | Peripheral resistance | 119 |
| 外周静脉压 | Peripheral venous pressure | 123 |
| 外周化学感受器 | Peripheral chemoreceptor | 177 |
| 微循环 | Microcirculation | 125 |
| 微胶粒 | Micelle | 206 |
| 微绒毛 | Microvilli | 213 |
| 微音器电位 | Microphonic potential | 292 |
| 胃蛋白酶原 | Pepsinogen | 195 |
| 胃动素 | Motilin | 189 |
| 胃排空 | Gastric empty | 201 |
| 网状结构上行激动系统 | Ascending reticular activating system | 329 |
| 位置性眼震颤 | Positional nystagmus | 342 |
| 舞蹈病 | Chorea | 344 |
X
| 性激素结合球蛋白 | Sex hormone-binding globulin,SHBG | 412 |
| 雄激素结合蛋白 | Androgen binding protein,ABP | 412 |
| 信号系统学说 | Signal system theory | 359 |
| 效应器酶(膜) | Effector enzyme | 21 |
| 腺苷酸环化酶 | Adenylate cyclase | 374 |
| 细胞间通道 | Intercellular channel | 21 |
| 兴奋性 | Excitability | 24 |
| 兴奋 | Excitation | 24 |
| 兴奋-收缩耦联 | Excitation-contraction coupling | 45 |
| 兴奋性突触后电位 | Excitatory postsynaptic potential | 319 |
| 相对不应期 | Relative refractory period | 107 |
| Rh血型系统 | Rh system | 81 |
| 血量 | Blood volume | 55 |
| 血小板 | Platelets,thrombocyte | 66,75 |
| 血清 | Serum | 68 |
| 血液凝固 | Blood coagulation | 68 |
| 血浆 | Plasma | 56 |
| 血型 | Blood flow | 77 |
| 血小板的聚集过程 | Velocity of blood flow | 75 |
| 血流量 | Blood flow | 116 |
| 血流速度 | Velocity of blood flow | 116 |
| 血压 | Bolld pressure | 116 |
| 血管升压素 | Vasopressin | 139 |
| 血-脑屏障 | Bolld-cerebrospinal fluid barrier | 148 |
| 血红蛋白 | Hemoglobin | 167 |
| 循环系统平均充盈压 | Mean circulatory filling pressure | 118 |
| 纤维蛋白溶解酶 | Plasmin | 73 |
| 纤维蛋白溶解系统 | Fibrinolytic system | 68,73 |
| 心电图 | Electrocardiogram | 113 |
| 心动周期 | Cardiac cycle | 85 |
| 心房收缩期 | Atrial systole | 86 |
| 收肌收缩性 | Myocardial contractility | 94 |
| 心力贮备 | Cardiac reserve | 96 |
| 心室功能曲线 | Ventricular function curve | 92 |
| 心室收缩期 | Ventricular systole | 86 |
| 心室舒张期 | Venrticular diastole | 88 |
| 心输出量 | Cardiac output | 89 |
| 心音 | Heart sound | 96 |
| 心音图 | Phonocardiogram | 96 |
| 心指数 | Cardiac index | 89 |
| 心作功 | Myocardial work | 90 |
| 心血管中枢 | Cardiovascular center | 133 |
| 心房利尿钠肽 | Atrial natriuretic peptide | 140,261 |
| 吸气 | Inspiration | 151 |
| 吸气切断机制 | Inspiratory off-switch mechanism | 175 |
| 吸收 | Absorption | 184 |
| 吸能反应 | Endergonic reaction | 218 |
| 小汗腺 | Eccrine gland | 233 |
| 小脑性共济失调 | Cerebellar ataxia | 243 |
| 胸式呼吸 | Thoracic breathing | 152 |
| 胸膜腔内压 | Intrapulmonary pressure | 154 |
| 胸腺素 | Thymosin | 409 |
| 消化 | Digestion | 154 |
| 行波理论 | Theory of traveling wave | 290 |
| 行为觉醒 | Behavioral arousal | 367 |
| 下运动神经元麻痹 | Lower motorneuron paralysis | 348 |
| 下丘脑调节肽 | Hypothalamus regulatory peptide,HRP | 377 |
Y
| 液态镶嵌模型 | Fluid massaic model | 8 |
| 易化扩散 | Facilitated diffusion | 12 |
| 易化区 | Facilitatory region | 339 |
| 原癌基因 | Proto-oncogene | 23 |
| 原肌凝蛋白 | Tropomyosin | 44 |
| 阈值 | Thresshold | 26 |
| 阈下刺激 | Subliminal(subthreshold)stimulus | 26 |
| α-银环蛇毒 | α-bungarotoxin | 41 |
| 有效不应期 | Effective refractory period | 107 |
| 有效滤过压 | Effective filtration pressure | 242,244 |
| 压力感受性反射 | Baroreceptor reflex | 135 |
| 用力呼吸 | Forced breathing | 160 |
| 余气量 | Residual volume | 159 |
| 氧容量 | Oxygen capacity | 168 |
| 氧含量 | Oxygen content | 168 |
| 氧饱和度 | Oxygen saturation | 168 |
| 氧离曲线 | Oxygen dissociation curve | 168 |
| 氧热价 | Thermal equivalent of oxygen | 220 |
| 抑胃肽 | Gastric inhibitory polypeptide(GIP) | 189 |
| 抑制性突触后 | Inhibitory postsynaptic potential | 322 |
| 抑制区 | Suppressor region | 339 |
| 抑制素 | Inhibin | 412 |
| 孕酮 | Progesterone,P | 416 |
| 月经 | Menstruation | 414 |
| 盐酸 | Hydrochloric acid | 194 |
| 胰高血糖素 | Glucagon | 405,407 |
| 胰岛素 | Insulin | 405 |
| 胰蛋白酶原 | Trypsinogen | 204 |
| 胰脂肪酶 | Lioase | 204 |
| 胰淀粉酶 | Pencreatic amylase | 203 |
| 应激 | Stress | 400 |
| 移动性运动综合波 | Migrating motility complex(MMC) | 209 |
| 眼震颤 | Nystagmus | 296 |
| 菸碱型受体 | Nicotinic receptor | 313 |
| 营养性作用 | Trophic action | 190,317 |
| 运动辅助区 | Supplementary motor area | 331 |
| α运动神经元 | α-motor neruon | 335 |
| γ运动神经元 | γ-motor neuron | 335 |
| 运动区 | Motor area | 331,345 |
| 运动柱 | Notor column | 346 |
| 运动失语症 | Motor aphasia | 362 |
| 一侧优势 | Laterality,cerebral dominance | 362 |
| 异相睡眠 | Paradoxical sleep | 368 |
| 诱发电位 | Evoked potential | 364 |
| 依赖cAMP激酶 | CAMP-deopendent protein kinase,cAMP-PK,PKA | 373 |
| 允许作用 | Permissive action | 373 |
Z
| 自身调节 | Autoregulation | 4 |
| 自动去极化 | Spontaneous depolarization | 102 |
| 自动节律性(自律性) | Auto-rhythmicity | 107 |
| 自律细胞 | Rhythmic cell | 97 |
| 质膜 | Plasma membrane | 7 |
| 载体 | Carrier | 12 |
| 主动转运 | Active transport | 13 |
| 终板电位 | End-plate potential | 40 |
| 造血干细胞 | Hemopoitric stem cells | 58 |
| 造血生长因子 | Hematopoietic growth factor,HGF | 62 |
| 中性粒细胞 | Neutrophils | 63 |
| 中心静脉压 | Central venous pressure | 123 |
| 中枢化学感受器 | Central chenoreceptor | 178 |
| 中枢延搁 | Central delay | 320 |
| 张力速度关系曲线 | Force velocity relation curve | 47,93 |
| 最大复极电位 | Maximal repolarization potential | 101 |
| 最适初长 | Optimal length | 47 |
| 最适前负荷 | Optimal preload | 47 |
| 阻力血管 | Resistance vessel | 115 |
| 轴流 | Axial flow | 118 |
| 轴突反射 | Axon reflex | 133 |
| 轴浆运输 | Axoplasmic transport,axoplaxmic flow | 303 |
| 组织液 | Interstitial fluid | 128,129 |
| 组胺 | Histamine | 141 |
| 周期性呼吸 | Periodic breathing | 181 |
| 咀嚼 | Mastication | 192 |
| 蒸发 | Evaporation | 232 |
| 致密斑 | Macular densa | 239 |
| 昼视觉 | Photopic vision | 279 |
| 姿势反射 | Postural refelex | 341 |
| 状态反射 | Attitudinal reflex | 341 |
| 震颤麻痹 | Paralysis agitans | 344 |
| 锥体系 | Pyramidal system | 346 |
| 锥体外系 | Extrapyramidal system | 346 |