1.糖皮质激素分泌的调节肾上腺皮质分泌皮质激素的束状带及网状带,处于腺垂体保肾上腺皮质激素(adrenocortiotropin,ACTH)的经常性控制之下,无论是糖皮质激素的基础分泌,还是在应激状态下的分泌,都受ACTH的调控,切除动物的垂体后,束状带与网状带萎缩,糖皮质激素有分泌显着减少,如及时补充ACTH,可使已发生萎缩的束状带与网状带基本恢复,糖皮质激素有分泌回升。
(1)ACTH:ACTH是一个含39个氨基酸的多肽,分子量为4500,其化学结构示于图11-17。
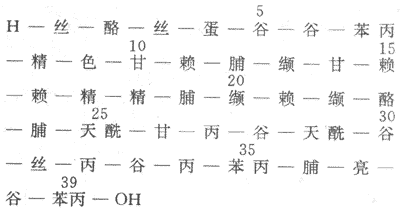
图11-17 人ACTH的化学结构
ACTH分子上的1-24位氨基酸为生物活性所必需的,25-39位氨基酸可保护激素,减慢降解,延长作用时间。各种动物的ACTH前24位氨基酸均相同,因此,从动物(牛、羊、猪等)腺垂体提到的ACTH对人有效。目前,ACTH已能人工合成。在垂体,ACTH是由阿黑皮素原(POMC)经酶分散而来,同时产生β-MSH。ACTH再经酶分解生成α-MSH,ACTH的第4-10位氨基酸与α-MSH第4-10位氨基酸和β-MSH第11-17位氨基酸相同,这部分氨基酸是产生MSH活性最小单位,因此ACTH也具有促黑素细胞产生黑色素的作用。
ACTH的分泌呈现日节律波动,入睡后ACTH分泌逐渐减少,午夜最低,随后又逐渐增多,至觉醒起床前进入分泌高峰,白天维持在较低水平,入睡时再减少。由于ACTH分泌的日节律波动,促糖皮质激素的分泌也出现相应的波动。ACTH分泌的这种日节律波动,是由下丘脑CRH节律性释放所决定的。
ACTH 不但刺激糖皮质激素的分泌,也刺激束状带与网状带细胞的生长发育,关于ACTH的作用机制已基本清楚。在束状还与网状带细胞膜上存在ACTH特异性受体,在Ca2+存在的条件下,ACTH与膜受体结合,激活腺苷酸环化酶,通过cAMP激活蛋白激酶,蛋白激酶起三项重要作用;①使核糖蛋白磷酸化,促进mRNA形成一种特殊蛋白质,使胆固醇得以进入线粒体,并经侧链解形成孕烯醇酮,以进一步合成糖皮质激素;②使磷酸化酶活化,促进糖原分解,产生ATP,提供能量,另外还通过戊糖旁路产生还原型辅酶Ⅱ(NADPH),以利胆固醇的羟化过程;③使胆固醇酯活化,促进其转变为胆固醇,提供激素合成的原料。在ACTH促进肾上腺皮质细胞合成糖皮质激素的同时,束状带细胞膜对葡萄糖与胆固醇的转运机制增强,使较多的葡萄糖与胆固醇进入细胞内(图11-18)。
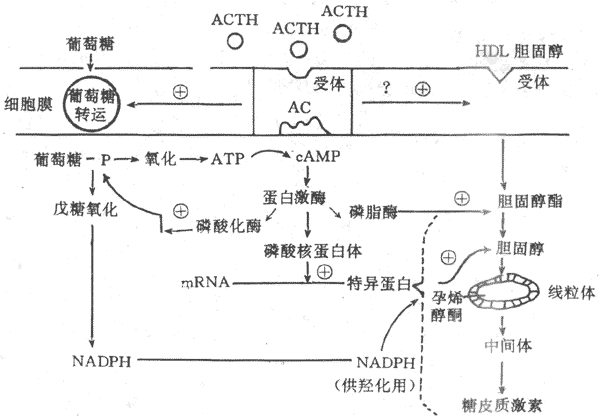
图11-18 ACTH作用机制示意图
HDL:高密度脂蛋白 AC:腺苷酸环化酶
(2)ACTH分泌的调节:ACTH调节糖皮质激素的分泌,而ACTH的分泌受下丘脑CRH的控制又与糖皮质激素有反馈调节。下丘脑CRH神经元和其他下丘脑调节肽神经元一样,又受脑内神经递质的调控。应激刺激作用于神经系统的不同部位,最后通过神经递质,将信息汇集于CRH神经元,然后借CRH控制腺垂体的促肾上腺皮质激素细胞分泌ACTH。此外,当血中糖皮质激素浓度升高时,可使腺垂体释放ACTH减少,ACTH的合成也受到抑制,腺垂体对CRH的反应也性减弱。糖皮质激素的负反馈调节主要作用于垂体,也可作用于下丘脑,这后一种反馈称为长反馈。ACTH还可反馈抑制CRH神经元,称为短反馈。至于是否存在CRH对CRH神经地的超短反馈,尚不能肯定。
综上所述,下丘脑、垂体和肾上腺皮质组成一个密切联系、协调统一的功能活动轴,从而维持血中糖皮质激素浓度的相对稳定和在不同状态下的适应性变化(图11-19)。
2.盐皮质激素分泌的调节 醛固酮的分泌主要受肾素-血管紧张素系统的调节。另外,血K+,血Na+浓度可以直接作用于球状带,影响醛固酮的分泌(详见第四章与第八章)。
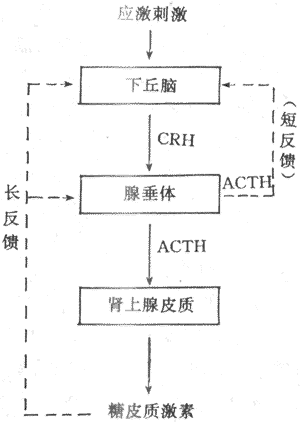
图11-19 糖皮质激素分泌的调节示意图
实线表示促进 点线表示抑制
在正常情况下,ACTH对醛固酮的分泌并无调节作用,但切除垂体后,在应激醛固酮的分泌反应减弱,提示在应激情况下,ACTH对醛固酮的分泌可能起到一定的支持作用。