旁路激活途径与经典激活途径不同之处在于激活是越过了C1、C4、C2三种成分,直接激活C3继而完成C5至C9各成分的连锁反应,还在于激活物质并非抗原抗体复合物而是细菌的细胞壁成分—脂多糖,以及多糖、肽聚糖、磷壁酸和凝聚的IgA和IgG4等物质。旁路激活途径在细菌性感染早期,尚未产生特异性抗体时,即可发挥重要的抗感染作用。
(一)生理情况下的准备阶段
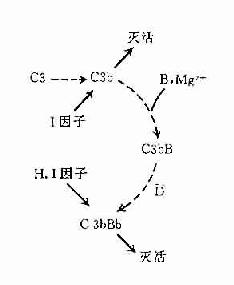
在正常生理情况下,C3与B因子、D因子等相互作用,可产生极少量的C3B和C3bBb(旁路途径的C3转化酶),但迅速受H因子和I因子的作用,不再能激活C3和后续的补体成分(图3-4,左)。只有当H因子和I因子的作用被阻挡之际,旁路途径方得以激活(图3-4,右)。
C3:血浆中的C3可自然地、缓慢地裂解,持续产生少量的C3b,释入液相中的C3b迅速被I因子灭活。
B因子:液相中缓慢产生的C3b在Mg2+存在下,可与B因子结合形成C3Bb。
D因子:体液中同时存在着无活性的D因子和有活性的D因子(B因子转化酶)。D因子作用于C3bB,可使此复合物中的B因子裂解,形成C3bBb和Ba游离于液相中。C3bBb可使C3裂解为C3a和C3b,但烊际上此酶效率不高亦不稳定,H因子可置换C3bBb复合物中的Bb,使C3b与Bb解离,解离或游离的C3b立即被I因子灭活。因此,在无激活物质存在的生理情况下,C3bBb保持在极低的水平,不能大量裂解C3,也不能激活后续补体成分。但是这种C3的低速度裂解和低浓度C3bBb的形成,具有重大意义。可比喻为处于“箭在弦上,一触即发”的状态。
(二)旁路途径的激活
旁路途径的激活在于激活物质(例如细菌脂多糖、肽聚糖;病素感染细胞、肿瘤细胞,痢疾阿米巴原虫等)的出现。目前认为,激活物质的存在为C3b或C3bBb提供不易受H因子置换Bb,不受Ⅰ因子灭活C3b的一种保护性微环境,使旁路激活途径从和缓进行的准备阶段过渡到正式激活的阶段(图3-4)。
·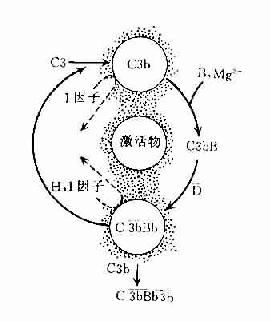
图3-4 旁路途径的激活
左:在正常后理情况下,可产生出少量C3bBb,但迅即被激活。
右:在激活物存在下,C3b不易被I因子灭活,C3bBb中的Bb不易被H因子置换,使激活过程得以进行。
P因子:P因子旧称备解素(properdin)。正常血浆中也有可以互相转换的两种P因子,P和P。C3bBb的半衰期甚短,当其与P因子结合成为C3bBbP时,半衰期可延长。这样可以获得更为稳定的、活性更强的C3转化酶。
C3bBb3b:C3bBb与其裂解C3所产生的C3b可进一步形成多分子复合物C3bBb3b。C3bBb3b像经典途径中的C5转化酶C423一样,也可使C5裂解成C5a和C5b。后续的C6~C9各成分与其相互作用的情况与经典途经相同。
(三)激活效应的扩大
C3在两条激活途径中都占据着重要的地位。C4是血清中含量最多的补体成分,这也正是适应其作用之所需。不论在经典途径还是在旁路途径,当C3被激活物质激活时,其裂解产物C3b又可在B因子和D因子的参与作用下合成新的C3bBb。后者又进一步使C3裂解。由于血浆中有丰富的C3,又有足够的B因子和Mg2+,因此这一过程一旦被触发。就可能激活的产生显著的扩大效应。有人称此为依赖C3Bb的正反馈途径,或称C3b的正反馈途径(图3-5)。
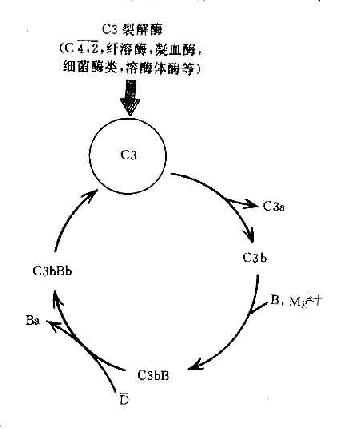
图3-5 C3b的正反馈途径