对人主要组织兼容性抗原系统及其基因复合体的认识比小鼠约晚10年,法国学者Dausset在1958年首先发现,肾移植后出现排斥反应的患者以及多次输血的患者血清中含有能与供者白细胞发生反应的抗体。后者所针对的抗原即人类主要组织相溶性抗原。由于该抗原首先在白细胞表面被发现且含量最高,而且白细胞抗原(human leucocyte antigen,HAL);人类MHC,即编码HLA的基因群自然数为HAL复合体。
(一)HLA复合体定位及结构
HLA复合体位于人第6号染色体的短臂上。该区DNA片段长度约3.5~4.0×千个碱基对,占人体整个基因组的1/3000。图5-2显示HLA复合体结构。HLA复合体共有数十个座,传统上按其产物的结构、表达方式、组织分布与功能可将这些基因座分为三类。
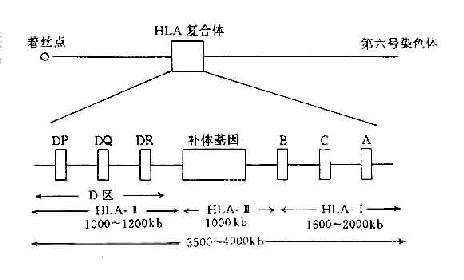
图5-2 人类HLA复合体结构示意图
1.HLA-Ⅰ类基因在Ⅰ基因区内存在多达31个有关的Ⅰ类基因座,其中HLA-A、HLA-B和HLA-C为经典的HLA-Ⅰ类基因,其它基因的产物分布有限,且其功能不明,另外还有许多伪基因.
2.HLA-Ⅱ类基因 HLA-Ⅱ类基因区包括近30个基因座,其中经典的Ⅱ类基因一般指DR、DP和DQ,它们编码的产物均为双肽链(α、β)分子。近年来,陆续发现了一些位于Ⅱ类基因区的新基因座,其中某些基因的产物与内源性抗原的处理与呈递有关。
3.HLA-Ⅲ类基因 HLA-Ⅲ类基因区域至少已发现36个基因座,其中C2、C4、Bf座编码相应的补体成分,另外还有21羧化酶基因(CYP21A、B)肿瘤坏死因子基因(TNFA、B)以及热休克蛋白70(heat shock protein70,HSP70)基因。补体C4由二个不同的基因(C4A与C4B)编码。HLA-Ⅲ类基因区结构见图5-3。
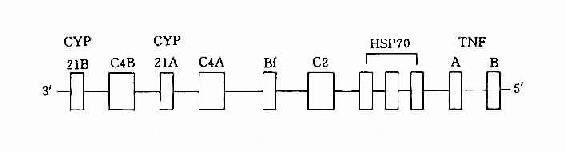
图5-3 HLA-Ⅲ基因区结构示意图
(二)HLA等位基因及编码产物的命名
按WHO-HLA命名委员会发布的资料,仅经典的HLA-Ⅰ、Ⅱ类座(A、B、C、DR、DQ、DP)等位基因即达279个。表5-1列出了至1991年11月已识别的HLA特异性。根据该委员会制定的命名原则,凡确定新的HLA抗原特异性都要明确其DNA序列。此外,下列几种情况在HLA特异性编号后加W(work shop)标记。
表5-1 已识别的HLA特异性(1991)
| A | B | C | D | DR | DQ | DF |
| A1 | B5 B51(5) | Cw1 | Dw1 | DR1 | DQ1 | DPw1 |
| A2 | B7 B5102 | Cw2 | Dw2 | DR103 | DQ2 | DPw2 |
| A210(2) | B703 B5103(7) | Cw3 | Dw3 | DR2 | DQ3 | DPw3 |
| A3 | B8 B52(5) | Cw4 | Dw4 | DR3 | DQ4 | DPw4 |
| A9 | B12 B53 | Cw5 | Dw5 | DR4 | DQ5(1) | DPw6 |
| A10 | B13 B54(22) | Cw6 | Dw6 | DR5 | DQ6(1) | |
| A11 | B14 B55(22) | Cw7 | Dw7 | DR6 | DQ7 | |
| A19 | B15 B54(22) | Cw8 | Dw8 | DR7 | DQ18(3) | |
| A23(9) | B16 B57(17) | Cw9(w3) | Dw10 | DR8 | DQ19(3) | |
| A24(9) | B17 B54(17) | Cw1(w3) | Dw11(w7) | DR9 | ||
| A2403(9) | B18 B59 | Dw12 | DR10 | |||
| A25(10) | B21 B60(40) | Dw13 | DR11(5) | |||
| A26(10) | B22 B61(40) | Dw14 | DR12(5) | |||
| A28 | B27 B62(15) | Dw15 | DR13(5) | |||
| A29(19) | B35 B63(15) | Dw16 | DR13(6) | |||
| A30(19) | B37B64k(14) | Dw17(w7) | DR14(6) | |||
| A31(19) | B38(16 B65(14) | Dw18(w6) | DR1403 | |||
| A32(19) | B39(16) B67 | Dw19(w6) | DR1404 | |||
| A33(19) | B40 B70 | Dw20 | DR15(2) | |||
| A34(10) | B4005(21) B71(70) | Dw21 | DR16(2) | |||
| A36 | B41 B72(70) | Dw22 | DR17(3) | |||
| A43 | B42 B73 | Dw23 | DR18(3) | |||
| A66(10) | B44(12) B75(15) | DR51 | ||||
| A68(28) | B45(12) B76(15) | Dw24 | ||||
| A69(28) | B46 B77(15) | Dw25 | DR52 | |||
| A74(19) | B47 B7801 | B-26 | DR53 | |||
| B48 | ||||||
| B49(21) Bw4 | ||||||
| B50(21) Bw6 |
①Bw4和Bw6作为表位以其它B座等位基因个区别;②C座的特异性加w,以与补体相区分;③由经典细胞学分型方法鉴定D和DP特异性加W。
(三)HLA复合机遗传特征
HLA复合体具备某些有别于其它真核基因系统的特征。
1.单体型遗传方式 HLA复合体是一组紧密连锁的基因群。这些连锁在一条染色体上的等位基因很少发生同源染色体间的交换,构成一个单体型(haplotype)。在遗传过程中,HLA单体型作为一个完整的遗传单位由亲代传给子代。有必要区分HLA表型、基因型与单体型这三个概念。某一个体HLA抗原特异性型别称为表型(phenotype);HLA基因在体细胞两条染色体上的组合称为基因型(genotype);HLA基因在同一条染色体上的组合称为单体型(haplotype)(表5-2)。
| 受检者 | 甲 A1 A2 B8 B12 |
乙 A1 A1 B8 B12 |
丙 A1 A1 B8 B8 |
| 表型 | HLA-A1、2:B8、12 | HLA-A1:B8、12 | HLA-A1、B8 |
| 基因型 | HLA-A1、A2 HLA-B8、B12 |
HLA-A1、A1 HLA-B8、B12 |
HLA-A1、A1 HLA-B8、B8 |
| 单体型 | HLA-A1、B8/A2、B12 | HLA-A1、B8/A1、B12 | HLA-A1、B8/A1、B8 |
二倍体(diploid)生物的每一细胞均有两个同源染色体组,分别来自父母双方。故子女的HLA单体型也是一个来自父方,一个来自母方。在同胞之间比较HLA单体型型别只会出现下列三种可能性:二个单体型完全相同或完全不同的机率各占25%;有一个单体型相同的机率占50%。至于亲代与子代之间则必然有一个单体型相同,也只能有一个单体型相同(图5-40。这一遗传特点在器官移植供者的选择以及法医的亲子鉴定中得到了应用。
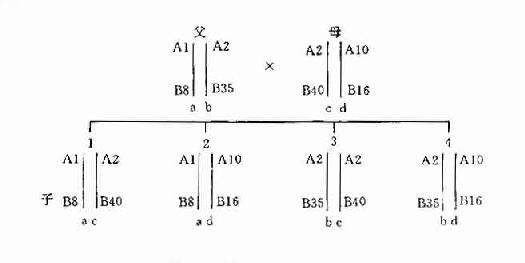
图5-4 HLA 单体型遗传示意图
注:a、b、c、d代表单体型
A1、B8、A2、B35等代表HLA基因座等位基因
2.多态性现象 多态性(polymorphism)是指在一随机婚配的群体中,染色体同一基因座有两种以上基因型,即可能编码二种以上的产物。HLA复合体是迄今已知人体最复杂的基因复合体,有高度的多态性。HLA的多态性现象乃由于下列原因所致:①复等位基因(multiple alleles):位于一对同源染色体上对应位置的一对基因称为等位基因(allele);由于群体中的突变,同一座的基因系列称为复等位基因。前已述及,HLA复合体的每一座均存在为数众多的复等位基因,这是HLA高度多态性的最主要原因。由于各个座位基因是随机组合的,故人群中的基因型可达108之多。②共显性(codominance);一对等位基因同为显性称为共显性。HLA复合体中每一个等位基因均为共显性,从而大大增加了人群中HLA表型的多样性,达到107数量级。因此,除了同卵双生外,无关个体间HLA型别全相同的可能性极小。
HLA的高度多态性显示了遗传背景的多样性,这可能是高等动物抵御不利环境因素的一种适应性表现,从而维持种属的生存与延续具有重要的生物意义,但也对组织移植过程中寻找配型合的供体带来很大的困难。
3.连锁不平衡 HLA复合体各等位基因均有其各自的基因频率。基因频率是指某一特定等位基因与该基因座中全部等位基因总和的比例。随机婚配的群体中,在无新的突变和自然选择的情况下,基因频率可以代代维持不变,由于HLA复合体和各基因座是紧密连锁的,若各座的等位基因随机组合构成单体型,则某一单体型型别的出现频率应等于该单体型各基因比其它基因能更多或更少地连锁在起,从而出现连锁不平衡(linkage disepuilibrium)。例如,在北欧白人中HLA-A1和HLA-B8频率分别为0.17和0.11。若随机组合,则单体型A1-B8的预期频率为0.17×0.11=0.019。但实际所测行的A1-B8单体型频率是0.088故A1-B8处于连锁不平衡,实测频率与预期频率间的差值(△0.088-0.19=0.069)为连锁不平衡参数。在HLA复合体中已发现有50对以上等位基因显示连锁不平衡。产生连锁不平衡的机制尚不清楚。