一、巨噬细胞与TH细胞之间的相互作用
TH细胞必须由静止状态的TH转变活化为TH才能辅助B细胞产生抗体。关于TH细胞活化的机制,根据现有实验资料提出一个双信号假说。
近年的实验证明在抗原呈递细胞(APC)表面至少有二种分子与TH细胞的活化相关。一种是抗原呈递分子,它是由MHC分子组成,它可与外源性或内源性抗原肽片段结合,然后运送至细胞表面并呈递给T细胞,通过TCR/CE3刺激产生第1活化信号。另一种分子即所谓协同刺激分子(costimulating molecules,CM),它是由一组粘附分子组成,它不仅能促进APC与T细胞的直接接触,而且也具有诱导信号传递的功能。这组分子可与T细胞上的协同刺激分子受体(costimulatory moleculesrecptor,CMR)结合,刺激其产生协同激信号,即所谓第2信号。
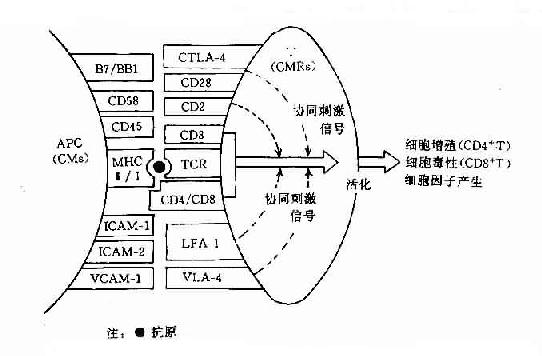
图11-2 T细胞活化主要协同刺激分子(CM)及其受体分子(CMR)的组成和作用示意图
T细胞上的CMR或称为辅助分子(accessory molecules)也是由一组粘附分子组成(表11-7)。对CMR及其配体的分子结构与功能及信号传导途径有待深入研究,但对其中的CD28分子和CTLA-4分子及其配体B7/BB1分子被认为是产生协同刺激信号的主要分子(图11-2)。
在这二种信号的作用下,才能使T细胞活化并合成和分泌IL-2和表达IL-2R,最终导致细胞分裂和克隆扩增。如无第2信号存在则T细胞不被活化也不引起克隆扩增,处于克隆不应答状态(clonol aneergy)(图11-3)。
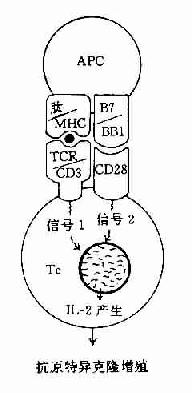
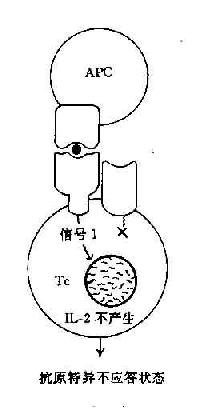
图11-3 协同刺激信号与T细胞活化状态
表11-7 T细胞表面主要辅助分子
| 名称 | 化学性质 | 基因族 | 细胞分子 | 配体分子(APC) | 功能 | |
| 粘附分子 | 信号传导 | |||||
| CD2(LFA-2) | 55kD单体 | Ig | 成熟T细胞 胸腺细胞 |
CD58(LFA-3) | + | + |
| CD4 | 55kD单体 | Ig | CD4+T细胞 | MHCⅡ分子 | + | + |
| CD8 | 78kD单体 | Ig | CD8+T细胞 | MHCⅡ分子 | + | + |
| CD11a/CD18 (LFA-1) |
180/95kD双体 | 粘合素 | 骨髓衍生细胞 | CD54(ICAM-1) | ? | + |
| CD28 | 90kD同二聚体 | CD4+ CD8+T(50%) |
B7/BB1 | + | + | |
| CDw49/CD29 (VLA-4、5、6) |
异二聚体 | 粘合素 | 白细胞 其它细胞 |
细胞外基质VCAM-1 | + | + |
| gP39(CD40L) | CD4+T细胞 | CD40 | ||||
阻断或给予第2信号,可以人为调节免疫应答使之增强或或抑制,对免疫治疗提供了新的手段。如阻断第2信号的产生可使T细胞处于免疫耐受状态,降低机体的免疫应答,这对防止移植排斥的发生和对超敏性疾病以及自身免疫性疾病的治疗是有利的。如将B7基因导入一定的肿瘤细胞则可增强机体的抗肿瘤免疫应答,目前这方面的实验研究已有较多的报道(图11-4)。
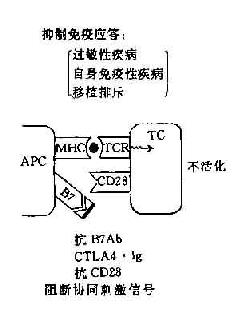
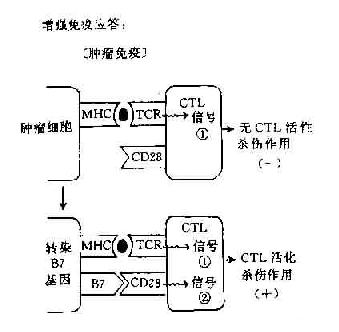
图11-4 协同刺激信号与免疫应答调节
二、TH细胞与B细胞之间的相互作用
(一)B细胞对T细胞依赖抗原的应答
一般情况下当大量抗原进入未经免疫的机体后,诱发初次免疫应答时其抗原呈递细胞多由巨噬细胞完成。经Mφ活化TH细胞后再由活化的TH细胞辅助B细胞产生抗体和形成记忆B细胞。但当再次免疫应答发生时,抗原呈递细胞则主要由已扩增的B细胞克隆承担。由于其膜Ig受体亲和力增高故对少量抗原也能摄取,故可取代巨噬细胞的抗原呈递作用。
过去曾认为B细胞与TH细胞间的相互作用是通过抗原的桥联作用使之发生接触,即B细胞的抗原受体可识别抗原分子的半抗原部分,而TH细胞的抗原受体可识别抗原分子的载体部分。但近年的实验证明B细胞与Mφ相同,也是通过抗原呈递作用与TH细胞相互作用。即B细胞可通过其膜Ig受体蛋白质抗原,即T依赖性抗原结合,经内化作用将抗原摄入。然后经过加工处理,使T细胞决定簇与MHCⅡ类分子结合,形成MHC-肽分子复合物并运送至B细胞表面,将其呈递给TH细胞,所以B细胞与TH细胞间的相互作用也是MHC限制性的。
B细胞和TH细胞通过其粘附分子及抗原呈递作用可彼此直接接触,并能相互诱导使之活化。活化B细胞在TH细胞的辅助下,B细胞最终增殖分化为合成和分泌各类免疫球蛋白分子的浆细胞。
B细胞可通过其抗原呈递作用刺激T细胞抗原受体(TCR/CD3)产生活化信号1,并通过其表面B7及其它协同刺激分子与T细胞表面的相应受体分子如CD28、IFA-1等相结合,可激发产生协同刺激信号(即信号2),在双信号作用下可使TH细胞活化。
B细胞要通过其BCR即表面Ig受体与抗原结合,Iga和Igβ链与CD3分子相当,可传递活化信号1。近年证明在B细胞表面表达的CD40分子可与T细胞表面的相应配体分子gP39(即CD40配体分子,CD40L)结合可激发其产生活化信号2,在此双信号作用下可使B细胞活化(图11-5)。
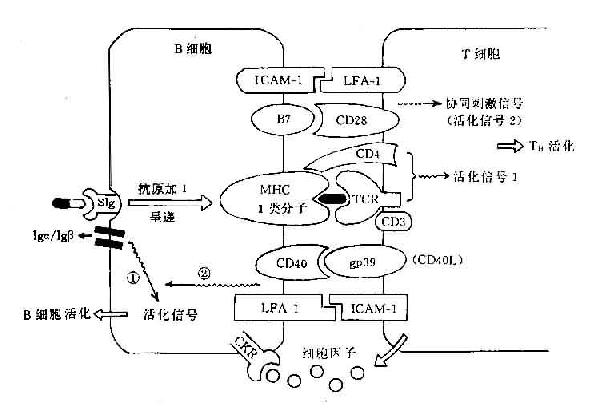
图11-5 B细胞与TH细胞间相互作用
活化的B细胞在活化的TH细胞分泌的多种细胞因子作用下,可增殖分化为分泌抗体的浆细胞和记忆B细胞。在再次应答时可发生Ig类别转换及抗体亲和力成熟等一系列变化。
(二)B细胞对非T细胞依赖抗原的应答
B细胞对非细胞依赖抗原(T1)的活化机制与T细胞依赖抗原完全不同。对Ⅰ型T1抗原,例如细菌脂多糖抗原,在高浓度时,它是B细胞的多克隆活化剂。这类抗原的决定簇部分可与B细胞表面的抗原受体结合,另外它具有促分裂素(mitogen)的结构,可与其相应受体结合。在这两部分结构的结合作用下,使B细胞多克隆活化。
而Ⅱ型T1抗原的结构特点是具有多个重复出现的抗原决定簇呈线性排列,在体内不易降解,例如肺炎球菌等多糖抗原。这些抗原决定簇对B细胞抗原受体亲和力强,由于受体交联而使B细胞活化(图11-6)。
A.Ⅰ型T1抗原是多克隆活化剂 B.Ⅱ型T1抗原有多个重复排的抗原决定簇使受体交联
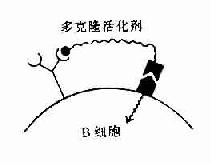
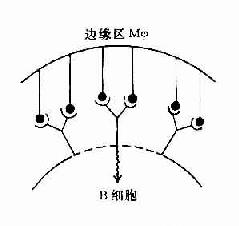
图11-6 T1抗原对B细胞的活化作用
T1抗原虽可被APC摄取,但不能被加工处理,所以不能与MHCⅡ类分子结合,故B细胞对T1抗原的应答无TH细胞参予,只表现初次应答的特性而不出现次应答的一系列变化。
三、巨噬细胞与B细胞的相互作用
这二种细胞间的相互作用尚不能肯定。但由于B细胞在大多数情况下需要有TH的协同,所以B细胞活化至少是间接的需要Mφ对B细胞的活化有直接作用,其分泌的IL-1对B细胞活化有促进作用(图11-7)。
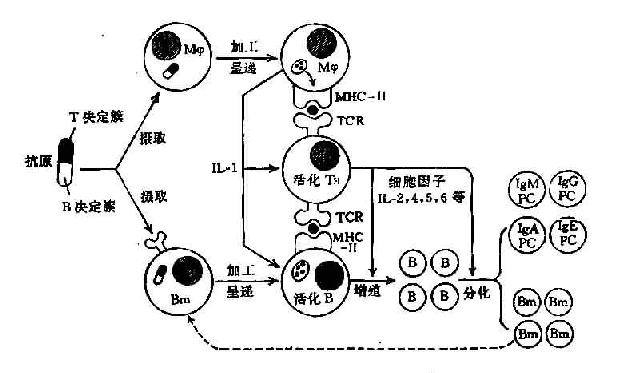
图11-7 抗体产生过程中免疫细胞间相互作用